
Saline-Tolerant Pathogenic Acanthamoeba spp. Isolated from a Geothermal Power Plant
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
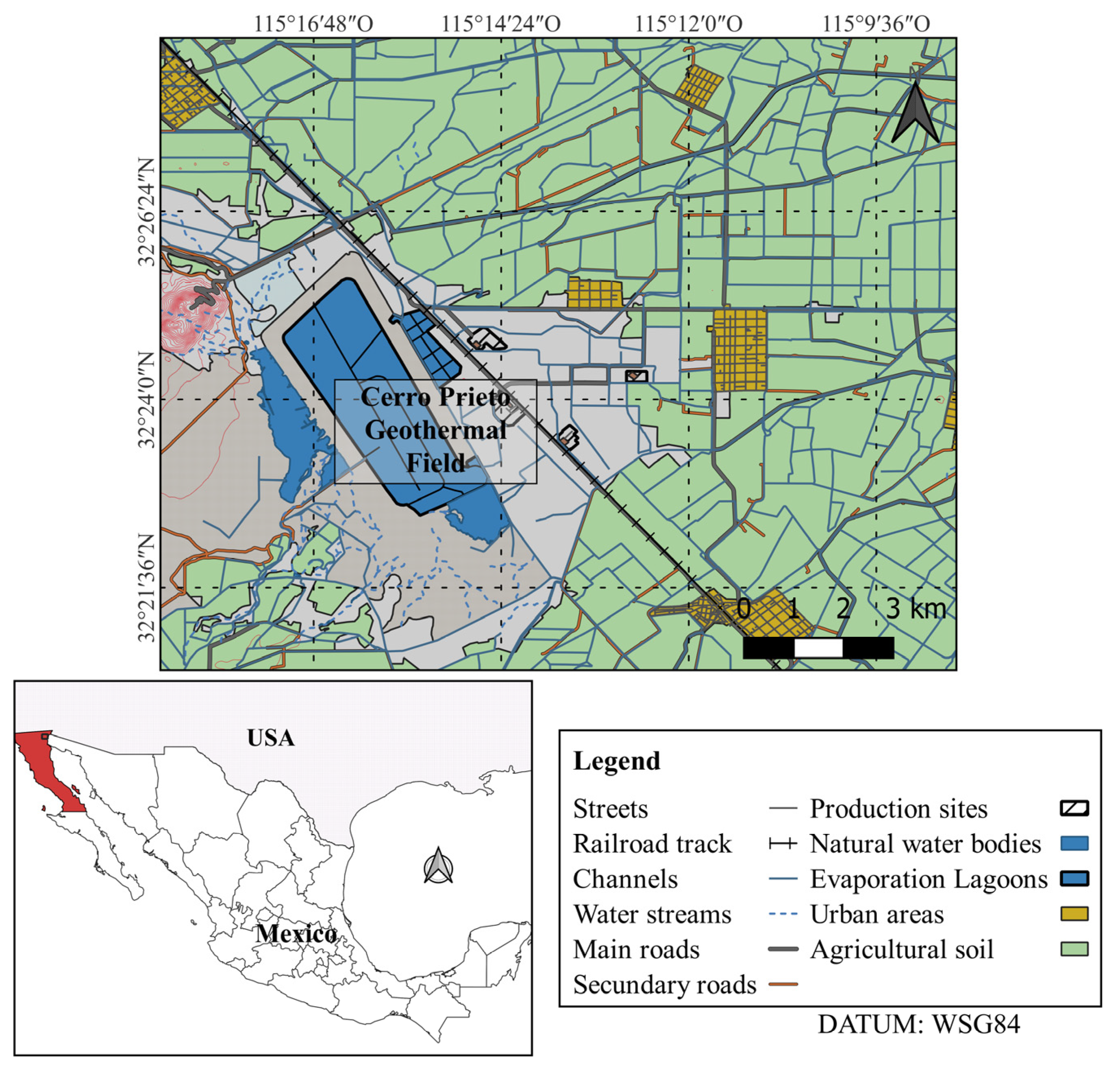
2.1. Sampling
2.2. Processing Samples
2.3. Morphological Identification
2.4. Pathogenicity Test
2.5. DNA Isolation and PCR
2.6. Sequencing and Phylogenetic Analysis
3. Results
3.1. Physicochemical Parameters
3.2. Presence of Free-Living Amoebae in the Sampling Sites
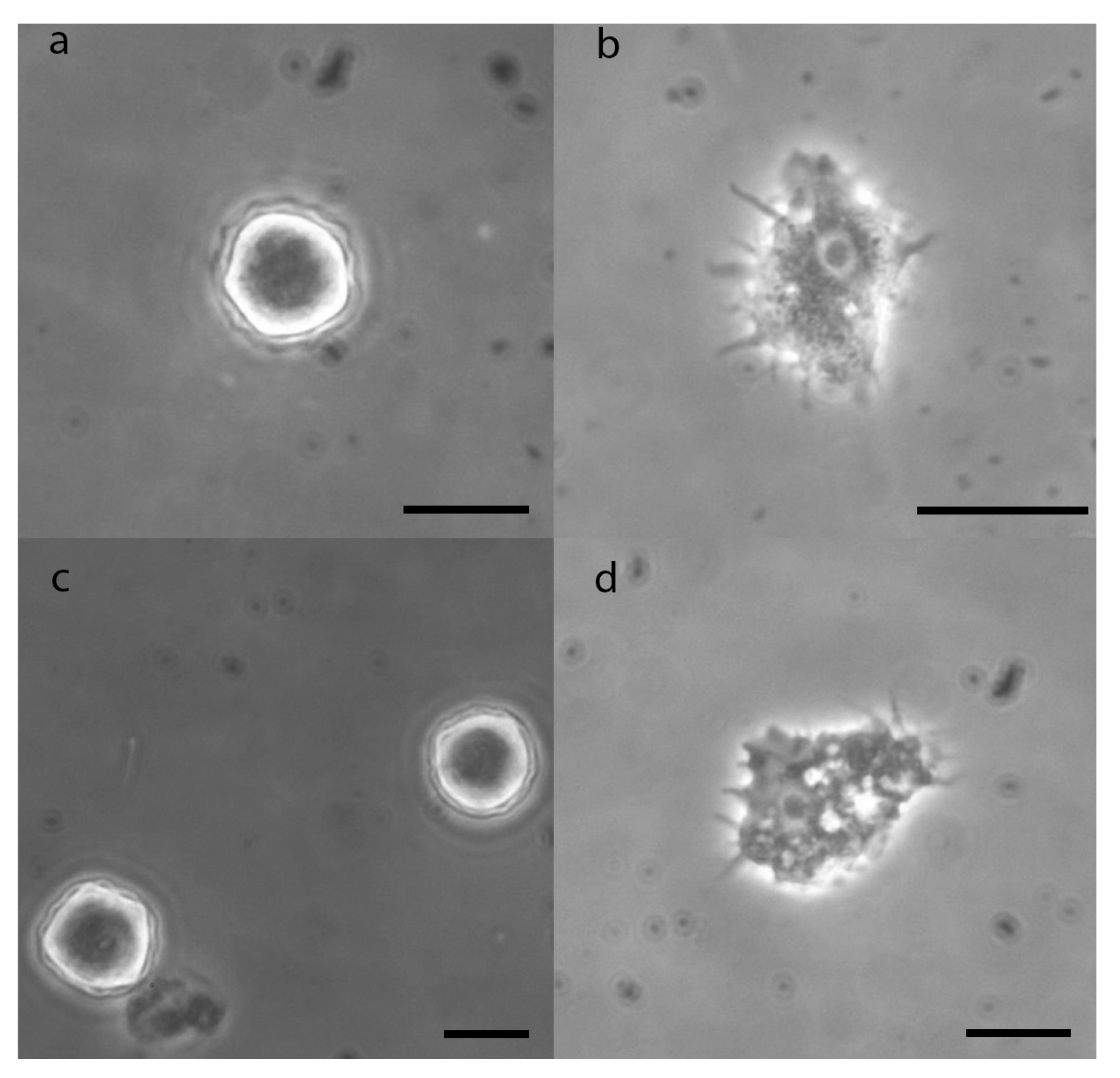
3.3. Morphological Description of Acanthamoeba
3.4. Morphological and Molecular Biology Identification of Acanthamoeba Strains
3.5. Pathogenicity Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Visvesvara, G.S. Infections with free-living amoebae. Handb. Clin. Neurol. 2013, 114, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Magnet, A.; Peralta, R.H.S.; Gomes, T.S.; Izquierdo, F.; Fernandez-Vadillo, C.; Galvan, A.L.; Pozuelo, M.J.; Pelaz, C.; Fenoy, S.; Del Águila, C. Vectorial role of Acanthamoeba in Legionella propagation in water for human use. Sci. Total Environ. 2015, 505, 889–895. [Google Scholar] [CrossRef]
- Scheid, P.L. Vermamoeba vermiformis—A free-living amoeba with public health and environmental health significance. Open Parasitol. J. 2019, 7, 40–47. [Google Scholar] [CrossRef]
- De Jonckheere, J.; Van Dijck, P.; Van de Voorde, V. The effect of thermal pollution on the distribution of Naegleria fowleri. J. Hyg. 1975, 75, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Dive, D.; Delattre, J.M.; Leclerc, H. Occurrence of thermotolerant amoebae in an electric power plant cooling pond. J. Therm. Biol. 1982, 7, 11–14. [Google Scholar] [CrossRef]
- Huizinga, H.W.; McLaughlin, G.L. Thermal ecology of Naegleria fowleri from a power plant cooling reservoir. Appl. Environ. Microbiol. 1990, 56, 2200–2205. [Google Scholar] [CrossRef] [PubMed]
- Montalbano, D.F.; Novelletto, M.A.; Di Cave, D.; Berrilli, F. Identification and phylogenetic position of Naegleria spp. from geothermal springs in Italy. Exp. Parasitol. 2017, 183, 143–149. [Google Scholar] [CrossRef]
- Moussa, M.; De Jonckheere, J.F.; Guerlotté, J.; Richard, V.; Bastaraud, A.; Romana, M.; Talarmin, A. Survey of Naegleria fowleri in geothermal recreational waters of Guadeloupe (French West Indies). PLoS ONE 2013, 8, e54414. [Google Scholar] [CrossRef]
- Tyndall, R.L.; Ironside, K.S.; Metler, P.L.; Tan, E.L.; Hazen, T.C.; Fliermans, C.B. Effect of thermal additions on the density and distribution of thermophilic amoebae and pathogenic Naegleria fowleri in a newly created cooling lake. Appl. Environ. Microbiol. 1989, 55, 22–32. [Google Scholar] [CrossRef]
- Lloyd, D. Encystment in Acanthamoeba castellanii: A Review. Exp. Parasitol. 2014, 145, S20–S27. [Google Scholar] [CrossRef]
- Delafont, V.; Bouchon, D.; Héchard, Y.; Moulin, L. Environmental factors shaping cultured free-living amoebae and their associated bacterial community within drinking water network. Water 2016, 100, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Canals, O.; Serrano-Suárez, A.; Salvadó, H.; Méndez, J.; Cervero-Aragó, S.; Ruiz de Porras, V. Effect of chlorine and temperature on free-living protozoa in operational man-made water systems (cooling towers and hot sanitary water systems) in Catalonia. Environ. Sci. Pollut. Res. 2015, 22, 6610–6618. [Google Scholar] [CrossRef] [PubMed]
- Kagel, A.; Bates, D.; Gawell, K. Guide to Geothermal Energy and the Environment; Technical Report; United States Department of Energy: Washington, DC, USA, 2005. [Google Scholar] [CrossRef]
- Morales-Arredondo, I.; Armienta, M.A.; Segovia, N. Groundwater chemistry and overpressure evidence in Cerro Prieto Geothermal Field. Geofluids 2017, 2017, e2395730. [Google Scholar] [CrossRef]
- American Public Health Association; American Water Works Association; Water Environment Federation. Standard Methods for the Examination of Water and Wastewater, 19th ed.; American Public Health Association: Washington, DC, USA, 1995. [Google Scholar]
- Robles, E.; Ramírez, E.; Sáinz, M.G.; Martínez, B.; Ayala, R.; González, M.E. Microbiological and physico-chemical study of swimming pool water. Int. Res. J. Adv. Eng. Sci. 2019, 4, 15–20. [Google Scholar] [CrossRef]
- Bonilla-Lemus, P.; Rojas-Hernández, S.; Ramírez-Flores, E.; Castillo-Ramírez, D.A.; Monsalvo-Reyes, A.C.; Ramírez-Flores, M.A. Barrón-Graciano, K.; Reyes-Batlle, M.; Lorenzo-Morales, J.; Carrasco-Yépez, M.M. Isolation and identification of Naegleria species in irrigation channels for recreational use in Mexicali Valley, Mexico. Pathogens 2020, 9, 820. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Batlle, M.; Gabriel, M.F.; Rodríguez-Expósito, R.; Felgueiras, F.; Sifaoui, I.; Mourão, Z.; de Oliveira Fernandes, E.; Piñero, J.E.; Lorenzo-Morales, J. Evaluation of the occurrence of pathogenic free-living amoeba and bacteria in 20 public indoor swimming pool facilities. Microbiol. Open 2021, 10, e1159. [Google Scholar] [CrossRef] [PubMed]
- Hauer, G.; Rogerson, A.; Anderson, O.R. Platyamoeba pseudovannellida n. sp., a naked amoeba with wide salt tolerance isolated from the Salton Sea, California. J. Eukaryot. Microbiol. 2001, 48, 663–669. [Google Scholar] [CrossRef]
- Hauer, G.; Rogerson, A. Remarkable salinity tolerance of seven species of naked amoebae (Gymnamoebae). Hydrobiologia 2005, 549, 33–42. [Google Scholar] [CrossRef]
- Rogerson, A.; Hauer, G. Naked Amoebae (Protozoa) of the Salton Sea, California. Hydrobiologia 2002, 473, 161–177. [Google Scholar] [CrossRef]
- Smirnov, A.V. Vertical distribution and abundance of Gymnamoebae amoebae (Rhizopoda) in bottom sediments of the Brackish Water Nivå Bay (Baltic Sea, The Sound). Protist 2002, 153, 239–250. [Google Scholar] [CrossRef]
- Smirnov, A.V. Cryptic freshwater amoeba species in the bottom sediments of Nivå Bay (Øresund, Baltic Sea). Eur. J. Protistol. 2007, 43, 87–94. [Google Scholar] [CrossRef]
- SEMARNART; CFE. Proyecto Geotermoeléctrico Cerro Prieto. Manifestación de Impacto Ambiental; Modalidad Particular, Secretaría de Manejo de Recursos Naturales: Mexico, 2007; Available online: http://sinat.semarnat.gob.mx (accessed on 27 August 2023).
- Rivera, F.; Roy-Ocotla, G.; Rosas, I.; Ramirez, E.; Bonilla, P.; Lares, F. Amoebae isolated from the atmosphere of Mexico City and environs. Environ. Res. 1987, 42, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Page, F.C. A New Key to Freshwater and Soil Gymnamoebae: With Instructions for Culture; Freshwater Biological Association: Cumbria, UK, 1988. [Google Scholar]
- De Jonckheere, J.F. Growth characteristics, cytopathic effect in cell culture, and virulence in mice of 36 type strains belonging to 19 different Acanthamoeba spp. Appl. Environ. Microbiol. 1980, 39, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, J.M.; Booton, G.C.; Hay, J.; Niszl, I.A.; Seal, D.V.; Markus, M.B.; Fuerst, P.A.; Byers, T.J. Use of subgenic 18S ribosomal DNA PCR and sequencing for genus and genotype identification of Acanthamoeba from humans with keratitis and from sewage sludge. J. Clin. Microbiol. 2001, 39, 1903–1911. [Google Scholar] [CrossRef] [PubMed]
- Portugal, E.; Pal Verma, M. Hydrochemistry of the evaporation lagoon in Cerro Prieto, Baja California, Mexico. Hidroquímica de la laguna de evaporación en Cerro Prieto, Baja California, México. Ing. Hidráulica México 2001, XVI, 153–173. [Google Scholar]
- Delafont, V.; Rodier, M.H.; Maisonneuve, E.; Cateau, E. Vermamoeba vermiformis: A free-living amoeba of interest. Microb. Ecol. 2018, 76, 991–1001. [Google Scholar] [CrossRef]
- Siddiqui, R.; Makhlouf, Z.; Khan, N.A. The increasing importance of Vermamoeba vermiformis. J. Eukaryot. Microbiol. 2021, 68, e12857. [Google Scholar] [CrossRef] [PubMed]
- Mantyla, A.W. Comparaciones de estándares de agua de mar. J. Phys. Oceanogr. 1987, 17, 543–548. [Google Scholar] [CrossRef]
- Tiewchaloren, S.; Junnu, V. Factors affecting the viability of pathogenic Naegleria species isolated from Thai patients. J. Trop. Med. Parasitol. 1999, 22, 172–178. [Google Scholar]
- Todd, C.D.; Reyes-Batlle, M.; Piñero, J.E.; Martínez-Carretero, E.; Valladares, B.; Streete, D.; Lorenzo-Morales, J.; Lindo, J.F. Isolation and molecular characterization of Acanthamoeba genotypes in recreational and domestic water sources from Jamaica, West Indies. J. Water Health 2015, 13, 909–919. [Google Scholar] [CrossRef]
- Booton, G.C.; Visvesvara, G.S.; Byers, T.J.; Kelly, D.J.; Fuerst, P.A. Identification and distribution of Acanthamoeba species genotypes associated with nonkeratitis infections. J. Clin. Microbiol. 2005, 43, 1689–1693. [Google Scholar] [CrossRef] [PubMed]
- CONAGUA. Actualización de la Disponibilidad Media Anual en el Acuífero Valle de Mexicali (0210), Estado de Baja California; CONAGUA, Subdirección General Técnica, Gerencia De Aguas Subterránea: Ciudad de Mexico, Mexico, 2020. [Google Scholar]
- Rivera, F.; Lares, F.; Gallegos, E.; Ramirez, E.; Bonilla, P.; Calderon, A.; Martinez, J.J.; Rodriguez, S.; Alcocer, J. Pathogenic amoebae in natural thermal waters of three resorts of Hidalgo, Mexico. Environ. Res. 1989, 50, 289–295. [Google Scholar] [CrossRef]
- Rivera, F.; Ramirez, E.; Bonilla, P.; Calderon, A.; Gallegos, E.; Rodriguez, S.; Ortiz, R.; Zaldívar, B.; Ramírez, P.; Duran, A.N. Pathogenic and free-living amoebae isolated from swimming pools and physiotherapy tubs in Mexico. Environ. Res. 1993, 62, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, E.; Robles, E.; Sáinz, M.G.; Ayala, R.; Campoy, E. Microbiological quality of the aquifer of Zacatepec, Morelos, Mexico. Calidad microbiológica del acuífero de Zacatepec, Morelos, México. Rev. Int. Contam. Ambient. 2009, 5, 98–105. [Google Scholar]
- Bonilla-Lemus, P.; Ramírez-Bautista, G.A.; Zamora-Muñoz, C.; Ibarra-Montes, M.R.; Ramírez-Flores, E.; Hernández-Martínez, M.D. Acanthamoeba spp. in domestic tap water in houses of contact lens wearers in the Metropolitan Area of Mexico City. Exp. Parasitol. 2010, 126, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Bonilla-Lemus, P.; Caballero, A.; Carmona, J.; Lugo, A. Occurrence of free-living amoebae in streams of the Mexico Basin. Exp. Parasitol. 2014, 145, S28–S33. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, E.; Robles, E.; Martínez, B.; Ayala, R.; Sáinz, M.G.; Martínez, M.E.; Gonzalez, M.E. Distribution of free-living amoebae in a treatment system of textile industrial wastewater. Exp. Parasitol. 2014, 145, S34–S38. [Google Scholar] [CrossRef]
- Ramírez, E.; Robles, E.; Martínez, M.E.; Sáinz, M.G.; Martínez, B.; Rivas, B.I.; Rocha, A. Distribution of free-living amoebae in springs in Morelos, Mexico. Glob. Adv. Res. J. Microbiol. 2016, 5, 57–67. [Google Scholar]
- Centeno, M.; Rivera, F.; Cerva, L.; Tsutsumi, V.; Gallegos, E.; Calderon, A.; Ortiz, R.; Bonilla, P.; Ramirez, E.; Suarez, G. Hartmannella vermiformis isolated from the cerebrospinal fluid of a young male patient with meningoencephalitis and bronchopneumonia. Arch. Med. Res. 1996, 27, 579–586. [Google Scholar]
- Kinnear, F.B. Cytopathogenicity of Acanthamoeba, Vahlkampfia and Hartmannella: Quantitative and qualitative in vitro studies on keratocytes. J. Infect. 2003, 46, 228–237. [Google Scholar] [CrossRef]
- Abedkhojasteh, H.; Niyyati, M.; Rahimi, F.; Heidari, M.; Farnia, S.; Rezaeian, M. First Report of Hartmannella keratitis in a cosmetic soft contact lens wearer in Iran. Iran. J. Parasitol. 2013, 8, 481–485. [Google Scholar]
- Todd, C.H.D.; Reyes-Batlle, M.; Valladares, B.; Lindo, J.F.; Lorenzo-Morales, J. Vannellid species isolated from freshwater source in a park in Jamaica, West Indies. Microbiol. Insights 2015, 8, 7–9. [Google Scholar] [CrossRef]
- Scheid, P. Mechanism of intrusion of a microspordian-like organism into the nucleus of host amoebae (Vannella sp.) isolated from a keratitis patient. Parasitol. Res. 2007, 101, 1097–1102. [Google Scholar] [CrossRef]
- Anderson, O.R.; Rogerson, A.; Hannah, F. Three new limax amoebae isolated from marine surface sediments: Vahlkampfia caledonica N. Sp., Saccamoeba marina N. Sp., and Hartmannella vacuolata N. sp. J. Eukaryot. Microbiol. 1997, 44, 33–42. [Google Scholar] [CrossRef]
- Corsaro, D.; Michel, R.; Walochnik, J.; Müller, K.D.; Greub, G. Saccamoeba lacustris sp. nov. (Amoebozoa: Lobosea: Hartmannellidae), a new lobose amoeba, parisitized by the novel chlamydia ´Candidatus Metachlamydia lacustris´ (Chlamydiae: Parachlamydiaceae). Eur. J. Protistol. 2010, 46, 86–95. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Sites | Site ID | pH | Water Temperature (° C) | Dissolved Oxygen (mg/L) | Conductivity (µS/cm) | * Salinity (%) |
|---|---|---|---|---|---|---|
| Secondary Discharge Channel 1 | SDC1 | 7.9 | 42.5 | 2.0 | 4.3 × 104 | 2.75 |
| Secondary Discharge Channel 2 | SDC2 | 8.1 | 43.0 | 1.8 | 4.2 × 104 | 2.68 |
| Primary Discharge Channel | PDC | 7.3 | 34.0 | 2.2 | 4.0 × 104 | 2.56 |
| Evaporation Lagoon 1 | EvL1 | 6.6 | 22.0 | 5.0 | 4.1 × 104 | 2.62 |
| Evaporation Lagoon 2 | EvL2 | 6.7 | 18.0 | 5.0 | 8.7 × 104 | 5.56 |
| Site | Genus |
|---|---|
| SDC1 | Acanthamoeba (Volkonsky 1931) Vannella (Bovee 1965) Vermamoeba (Cavalier-Smith and Smirnov 2011) |
| SDC2 | Acanthamoeba (Volkonsky 1931) Saccamoeba (Frenzel 1982 emend. Bovee 1972) Thecamoeba (Fromentel 1874) Vannella (Bovee 1965) |
| PDC | Acanthamoeba (Volkonsky 1931) Vannella (Bovee 1965) Vermamoeba (Cavalier-Smith and Smirnov 2011) |
| EvL1 | Acanthamoeba (Volkonsky 1931) |
| EvL2 | Acanthamoeba (Volkonsky 1931) |
| Strain | Morphology | Molecular Biology (JDP1/2) | ||
|---|---|---|---|---|
| Species | Genotype | GenBank Accession Number | ||
| SDC1 | Acanthamoeba Group III | Acanthamoeba culbertsoni | T10 | OR767829 |
| SDC2a | Acanthamoeba Group III | Acanthamoeba culbertsoni | T10 | OR767828 |
| SDC2b | Acanthamoeba Group III | Acanthamoeba lenticulata | T5 | OR767826 |
| PDC | Acanthamoeba Group III | Acanthamoeba culbertsoni | T10 | OR767830 |
| EvL1 | Acanthamoeba Group III | Acanthamoeba culbertsoni | T10 | OR767831 |
| EvL2 | Acanthamoeba Group III | Acanthamoeba lenticulata | T5 | OR767827 |
| Strain | Mortality (%) | Death Days | Organs from FLA Were Recovered | Temperature Tolerance (°C) |
|---|---|---|---|---|
| SDC1 | 60 | 15–17 | Brain and Lung | 30, 37 *, 42 |
| SDC2a | 100 | 2–11 | Brain | 30, 37 *, 42 |
| SDC2b | 20 | 16 | Brain and Lung | 30 *, 37, 42 |
| PDC | 80 | 12–13 | Brain and Lung | 30, 37 *, 42 |
| EvL1 | 40 | 18–20 | Lung | 30, 37 *, 42 |
| EvL2 | 0 | - | - | 30 *, 37, 42 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Flores, E.; Bonilla-Lemus, P.; Carrasco-Yépez, M.M.; Ramírez-Flores, M.A.; Barrón-Graciano, K.A.; Rojas-Hernández, S.; Reyes-Batlle, M.; Lorenzo-Morales, J. Saline-Tolerant Pathogenic Acanthamoeba spp. Isolated from a Geothermal Power Plant. Pathogens 2023, 12, 1363. https://doi.org/10.3390/pathogens12111363
Ramírez-Flores E, Bonilla-Lemus P, Carrasco-Yépez MM, Ramírez-Flores MA, Barrón-Graciano KA, Rojas-Hernández S, Reyes-Batlle M, Lorenzo-Morales J. Saline-Tolerant Pathogenic Acanthamoeba spp. Isolated from a Geothermal Power Plant. Pathogens. 2023; 12(11):1363. https://doi.org/10.3390/pathogens12111363
Chicago/Turabian StyleRamírez-Flores, Elizabeth, Patricia Bonilla-Lemus, María M. Carrasco-Yépez, Miguel A. Ramírez-Flores, Karla A. Barrón-Graciano, Saúl Rojas-Hernández, María Reyes-Batlle, and Jacob Lorenzo-Morales. 2023. "Saline-Tolerant Pathogenic Acanthamoeba spp. Isolated from a Geothermal Power Plant" Pathogens 12, no. 11: 1363. https://doi.org/10.3390/pathogens12111363