
First Report of Plasmid-Mediated Macrolide-Clindamycin-Tetracycline Resistance in a High Virulent Isolate of Cutibacterium acnes ST115


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Culture Conditions
2.2. Genomic DNA Preparation and Genome Sequencing
2.3. Sequence Analysis
2.4. Linearization of Plasmid DNA
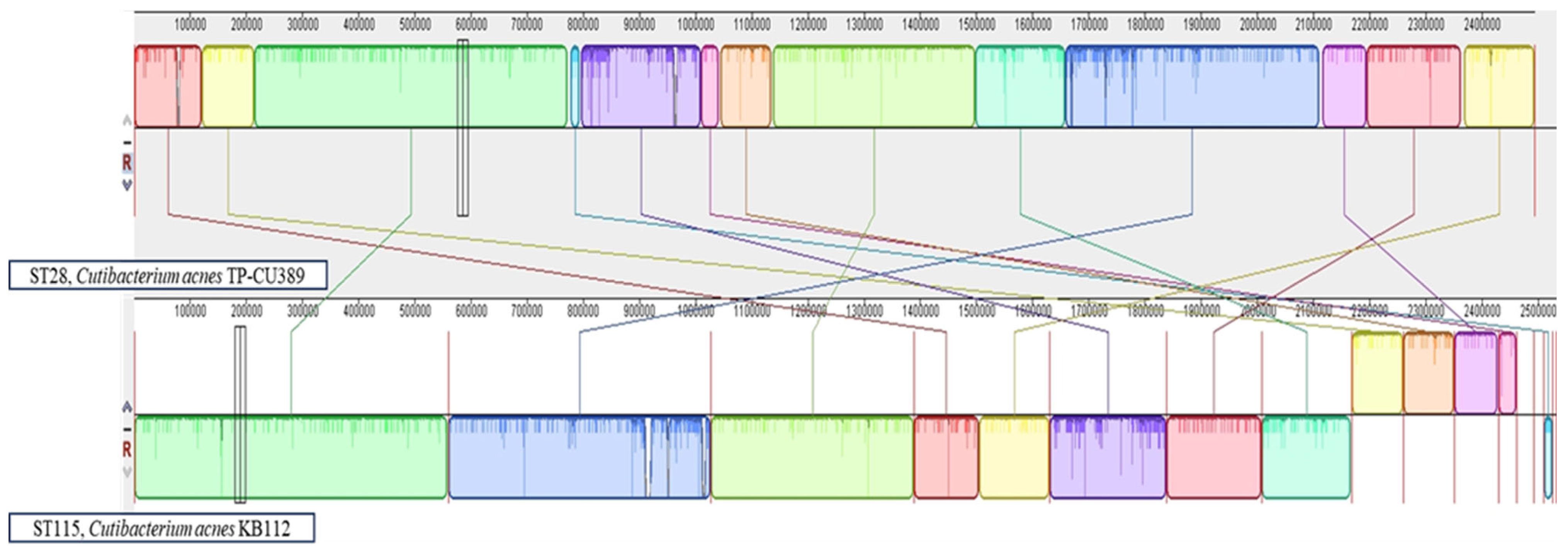
2.5. Phylogenetic and Genomic Arrangement Analysis
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Park, S.N.; Roh, H.; Lim, Y.K.; Kook, J.K. Complete genome sequence of Cutibacterium acnes KCOM 1861 isolated from a human jaw osteomyelitis lesion. Korean J. Microbiol. 2017, 53, 126–128. [Google Scholar]
- Hayashi, N.; Akamatsu, H.; Iwatsuki, K.; Shimada-Omori, R.; Kaminaka, C.; Kurokawa, I.; Kono, T.; Kobayashi, M.; Tanioka, M.; Furukawa, F.; et al. Japanese Dermatological Association Guidelines: Guidelines for the treatment of acne vulgaris 2017. J. Dermatol. 2018, 45, 898–935. [Google Scholar] [CrossRef]
- Kim, S.; Song, H.; Lee, W.J.; Kim, J. Antimicrobial susceptibility and characterization of Propionibacterium acnes by multilocus sequence typing and repetitive-sequence-based PCR. J. Bacteriol. Virol. 2016, 46, 135–141. [Google Scholar] [CrossRef]
- Wright, T.E.; Boyle, K.K.; Duquin, T.R.; Crane, J.K. Propionibacterium acnes Susceptibility and Correlation with Hemolytic Phenotype. Infect. Dis. 2016, 9, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Greenwood Quaintance, K.E.; Schuetz, A.N.; Shukla, D.R.; Cofield, R.H.; Sperling, J.W.; Patel, R.; Sanchez-Sotelo, J. Correlation between hemolytic profile and phylotype of Cutibacterium acnes (formerly Propionibacterium acnes) and orthopedic implant infection. Shoulder Elbow. 2020, 12, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.I.; Eady, E.A.; Carnegie, E.; Cove, J.H. Detection of transposon Tn5432-mediated macrolide-lincosamide-streptogramin B (MLSB) resistance in cutaneous propionibacteria from six European cities. J. Antimicrob. Chemother. 2002, 49, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Nakase, K.; Nakaminami, H.; Takenaka, Y.; Hayashi, N.; Kawashima, M.; Noguchi, N. A novel 23S rRNA mutation in Propionibacterium acnes confers resistance to 14-membered macrolides. J. Glob. Antimicrob. Resist. 2016, 6, 160–161. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.H.; Roh, H.S.; Kim, Y.H.; Kim, J.E.; Ko, J.Y.; Ro, Y.S. Antibiotic resistance of microbial strains isolated from Korean acne patients. J. Dermatol. 2012, 39, 833–837. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 8, 75. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [PubMed]
- Boyle, K.K.; Marzullo, B.J.; Yergeau, D.A.; Nodzo, S.R.; Crane, J.K.; Duquin, T.R. Pathogenic genetic variations of C. acnes are associated with clinically relevant orthopedic shoulder infections. J. Orthop. Res. 2020, 38, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Beirne, C.; McCann, E.; McDowell, A.; Miliotis, G. Genetic determinants of antimicrobial resistance in three multi-drug resistant strains of Cutibacterium acnes isolated from patients with acne: A predictive in silico study. Access Microbiol. 2022, 4, acmi000404. [Google Scholar] [CrossRef] [PubMed]
- Holland, C.; Mak, T.N.; Zimny-Arndt, U.; Schmid, M.; Meyer, T.F.; Jungblut, P.R.; Brüggemann, H. Proteomic identification of secreted proteins of Propionibacterium acnes. BMC Microbiol. 2010, 10, 230. [Google Scholar] [CrossRef] [PubMed]
- Picardeau, M.; Lobry, J.R.; Hinnebusch, B.J. Analyzing DNA strand compositional asymmetry to identify candidate replication origins of Borrelia burgdorferi linear and circular plasmids. Genome Res. 2000, 10, 1594–1604. [Google Scholar] [CrossRef]
- Aoki, S.; Nakase, K.; Nakaminami, H.; Wajima, T.; Hayashi, N.; Noguchi, N. Transferable Multidrug-Resistance Plasmid Carrying a Novel Macrolide-Clindamycin Resistance Gene, erm(50), in Cutibacterium acnes. Antimicrob. Agents Chemother. 2020, 64, e01810-19. [Google Scholar] [CrossRef]
- Fluit, A.C.; Visser, M.R.; Schmitz, F.J. Molecular detection of antimicrobial resistance. Clin. Microbiol. Rev. 2001, 14, 836–871. [Google Scholar] [CrossRef] [PubMed]
- Vilchez, H.H.; Escudero-Sanchez, R.; Fernandez-Sampedro, M.; Murillo, O.; Auñón, Á.; Rodríguez-Pardo, D.; Jover-Sáenz, A.; Del Toro, M.D.; Rico, A.; Falgueras, L.; et al. Prosthetic Shoulder Joint Infection by Cutibacterium acnes: Does Rifampin Improve Prognosis? A Retrospective, Multicenter, Observational Study. Antibiotics 2021, 10, 475. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, J.; Nakase, K.; Hayashi, N.; Takeo, C.; Nakaminami, H. Multidrug Resistance Plasmid pTZC1 Could Be Pooled among Cutibacterium Strains on the Skin Surface. Microbiol. Spectr. 2023, 11, e0362822. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
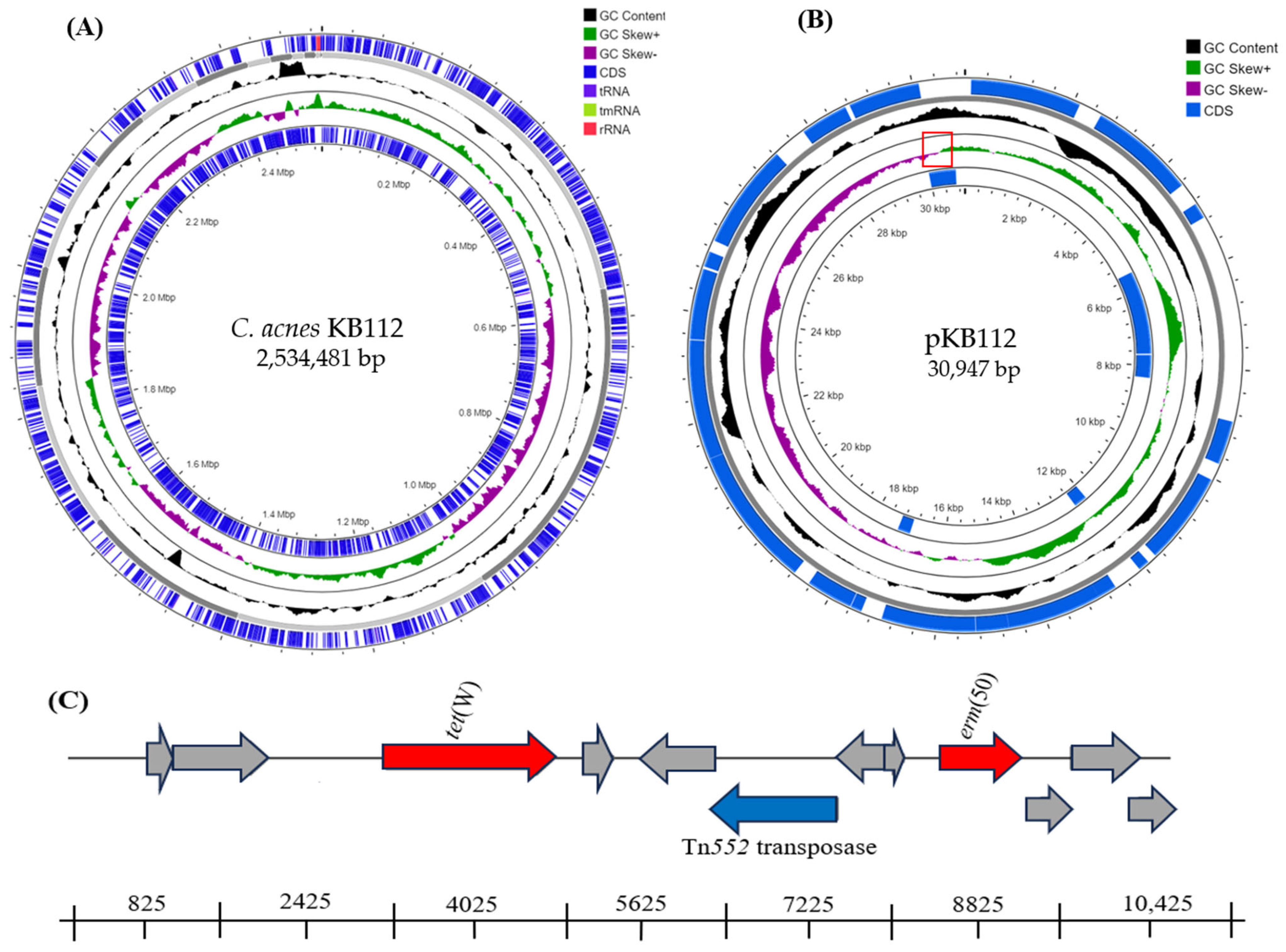
| No. | Location | Size (aa) | Description | Identity (%) | Accession No. |
|---|---|---|---|---|---|
| 1 | 112–1194 | 360 | DNA methyltransferase | 100 | WP_176453839 |
| 2 | 1184–2116 | 245 | hypothetical protein (plasmid) | 99.59 | BBJ25236 |
| 3 | 2541–4460 | 639 | TetW: tetracycline resistance ribosomal protection protein | 100 | WP_002586627 |
| 4 | 4873–5157 | 94 | recombinase (plasmid) | 100 | BBJ25234 |
| 5 | 6269–5454 | 271 | AAA family ATPase | 100 | WP_070434713 |
| 6 | 7687–6266 | 476 | Tn552 transposase (plasmid) | 100 | BBJ25232 |
| 7 | 8328–7723 | 202 | recombinase family protein | 99.5 | WP_234990909 |
| 8 | 8931–9722 | 263 | erm(50): rRNA adenine N-6-dimethyltransferase | 100 | WP_176453837 |
| 9 | 10037–10369 | 110 | transcriptional regulator | 100 | WP_176453836 |
| 10 | 10366–11142 | 258 | nucleotidyl transferase AbiEii/AbiGii toxin | 100 | WP_176453835 |
| 11 | 11139–11723 | 194 | recombinase family protein | 100 | WP_176453834 |
| 12 | 12632–12979 | 115 | plasmid mobilization relaxosome protein MobC | 100 | WP_176453832 |
| 13 | 12979–14685 | 569 | relaxase of type IV secretion system (plasmid) | 100 | BBJ25226 |
| 14 | 14687–15277 | 205 | DUF3801 domain-containing protein | 100 | WP_176453830 |
| 15 | 15288–16988 | 566 | type IV secretory system conjugative DNA transfer family protein | 100 | WP_176453829 |
| 16 | 18442–18978 | 178 | PrgI family protein | 100 | WP_176453825 |
| 17 | 19046–21316 | 756 | ATPase of type IV secretion system (plasmid) | 100 | BBJ25222 |
| 18 | 21322–22842 | 506 | bifunctional lytic transglycosylase/C40 family peptidase | 100 | WP_176453824 |
| 19 | 27866–28732 | 414 | anti-repressor of ImmR (plasmid) | 100 | BBJ25217 |
| 20 | 28816–29757 | 323 | ParA family protein | 100 | BBJ25216 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rana, M.S.; Kim, J.; Kim, S. First Report of Plasmid-Mediated Macrolide-Clindamycin-Tetracycline Resistance in a High Virulent Isolate of Cutibacterium acnes ST115. Pathogens 2023, 12, 1286. https://doi.org/10.3390/pathogens12111286
Rana MS, Kim J, Kim S. First Report of Plasmid-Mediated Macrolide-Clindamycin-Tetracycline Resistance in a High Virulent Isolate of Cutibacterium acnes ST115. Pathogens. 2023; 12(11):1286. https://doi.org/10.3390/pathogens12111286
Chicago/Turabian StyleRana, Md Shohel, Jungmin Kim, and Shukho Kim. 2023. "First Report of Plasmid-Mediated Macrolide-Clindamycin-Tetracycline Resistance in a High Virulent Isolate of Cutibacterium acnes ST115" Pathogens 12, no. 11: 1286. https://doi.org/10.3390/pathogens12111286