
Plant-Disease-Suppressive and Growth-Promoting Activities of Endophytic and Rhizobacterial Isolates Associated with Citrullus colocynthis

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Bacteria Isolation from Citrullus colocynthis Rhizosphere and Endosphere
2.3. DNA Extraction, PCR Amplification, and Sequence Analysis of the Bacterial Strains
2.4. Screening for Antagonistic Activity
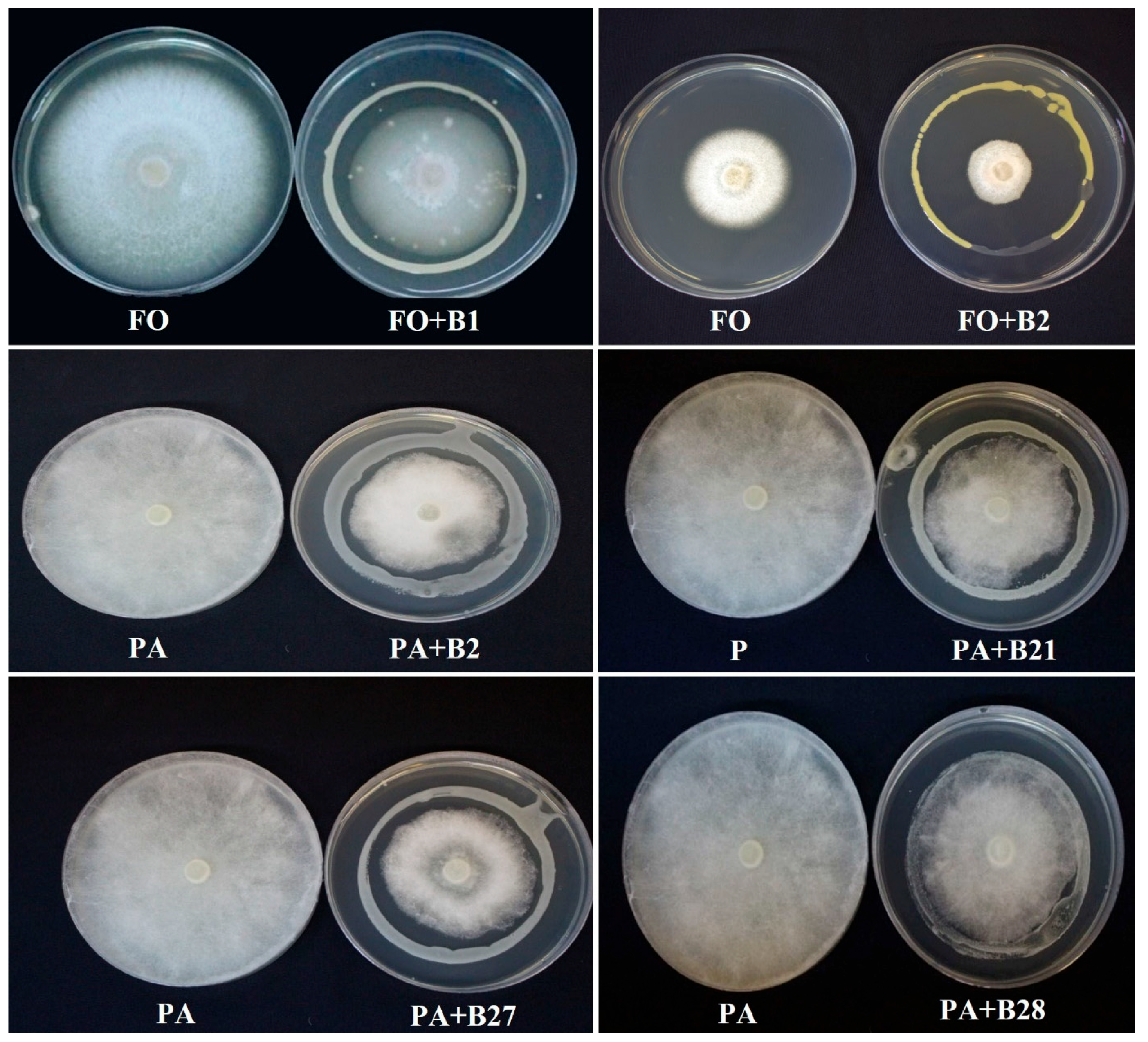
2.4.1. In Vitro Antifungal Assays
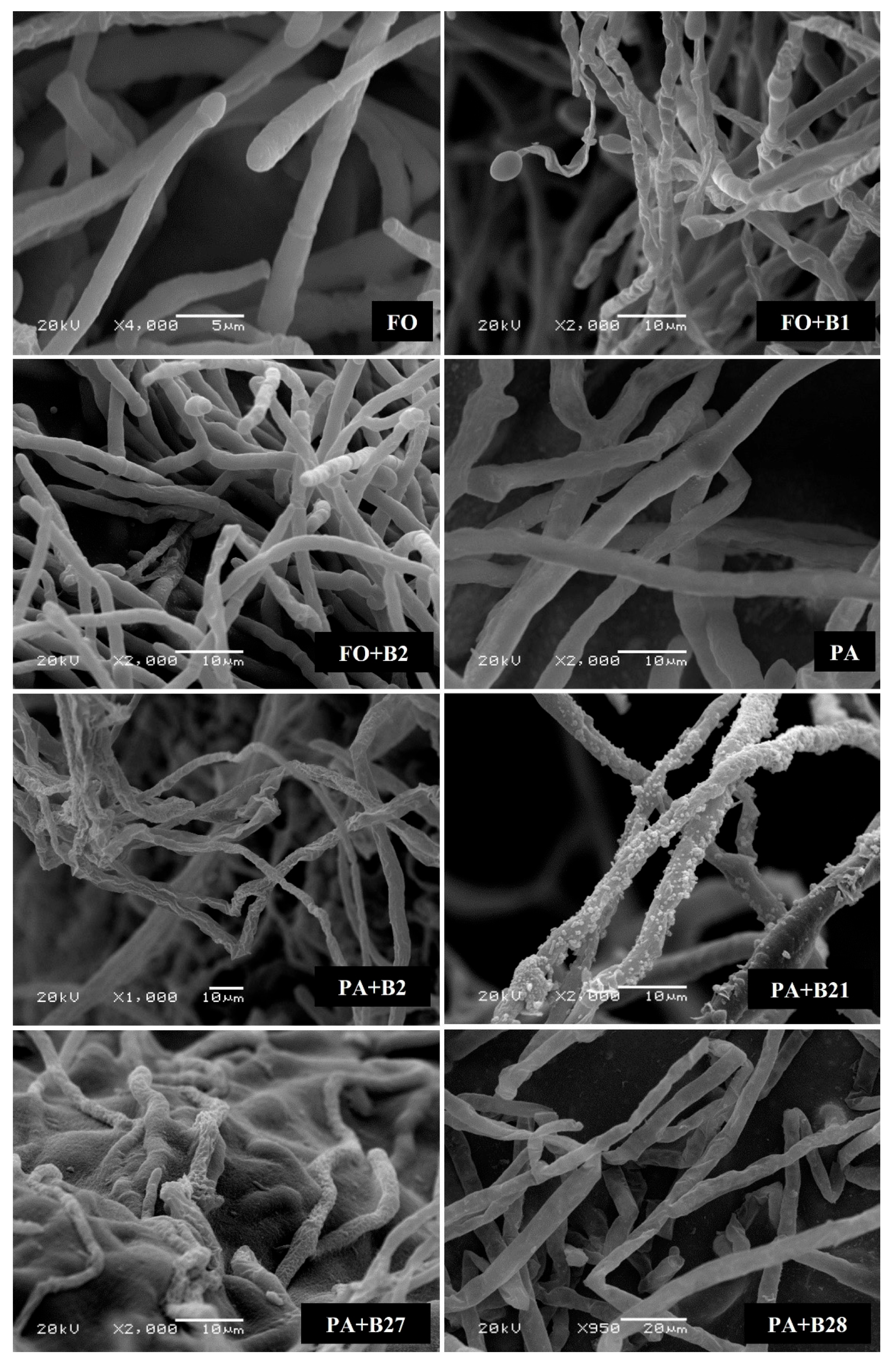
2.4.2. Scanning Electron Microscope (SEM)
2.5. Determination of Growth-Promoting Potential
2.5.1. Indole Acetic Acid (IAA) Quantification
2.5.2. Seed Germination Test via Paper Towel Method
2.6. Statistical Analysis
3. Results
3.1. Isolation of Endophytic and Rhizospheric Bacteria
3.2. Molecular Identification
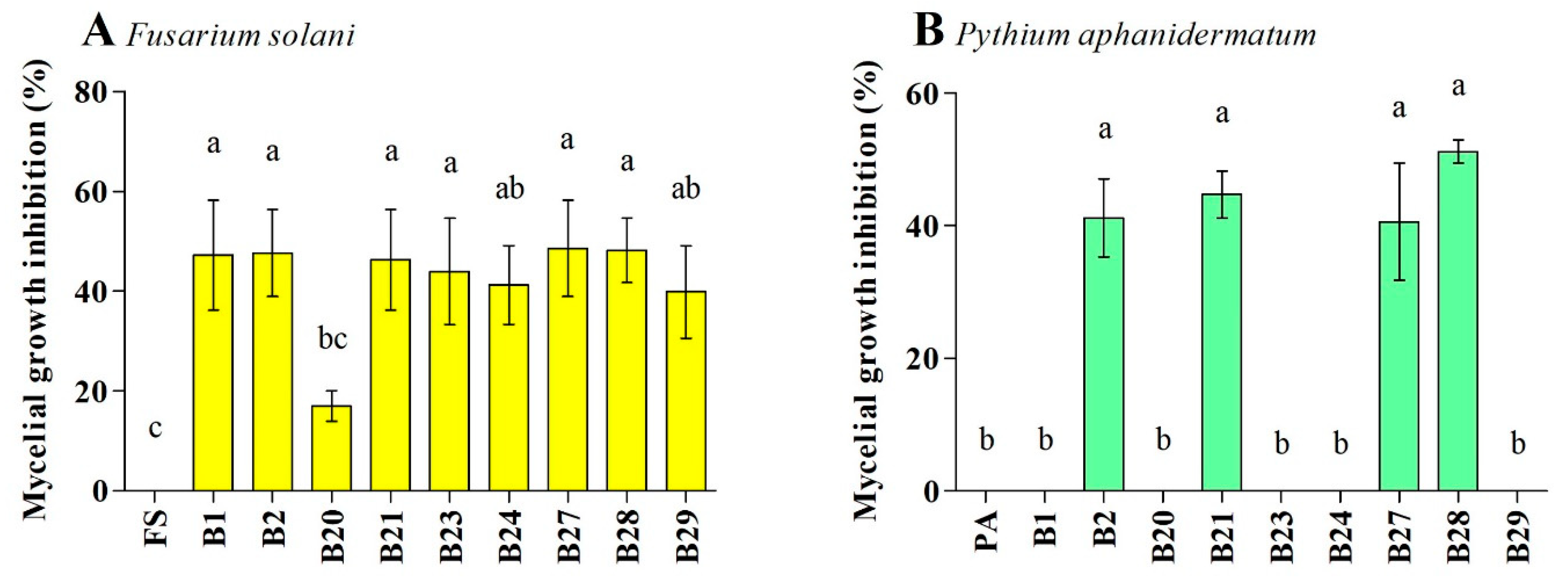
3.3. Antagonistic Activity against F. solani and P. aphanidermatum
3.4. Growth-Promoting Assays
3.4.1. Determination of IAA Production
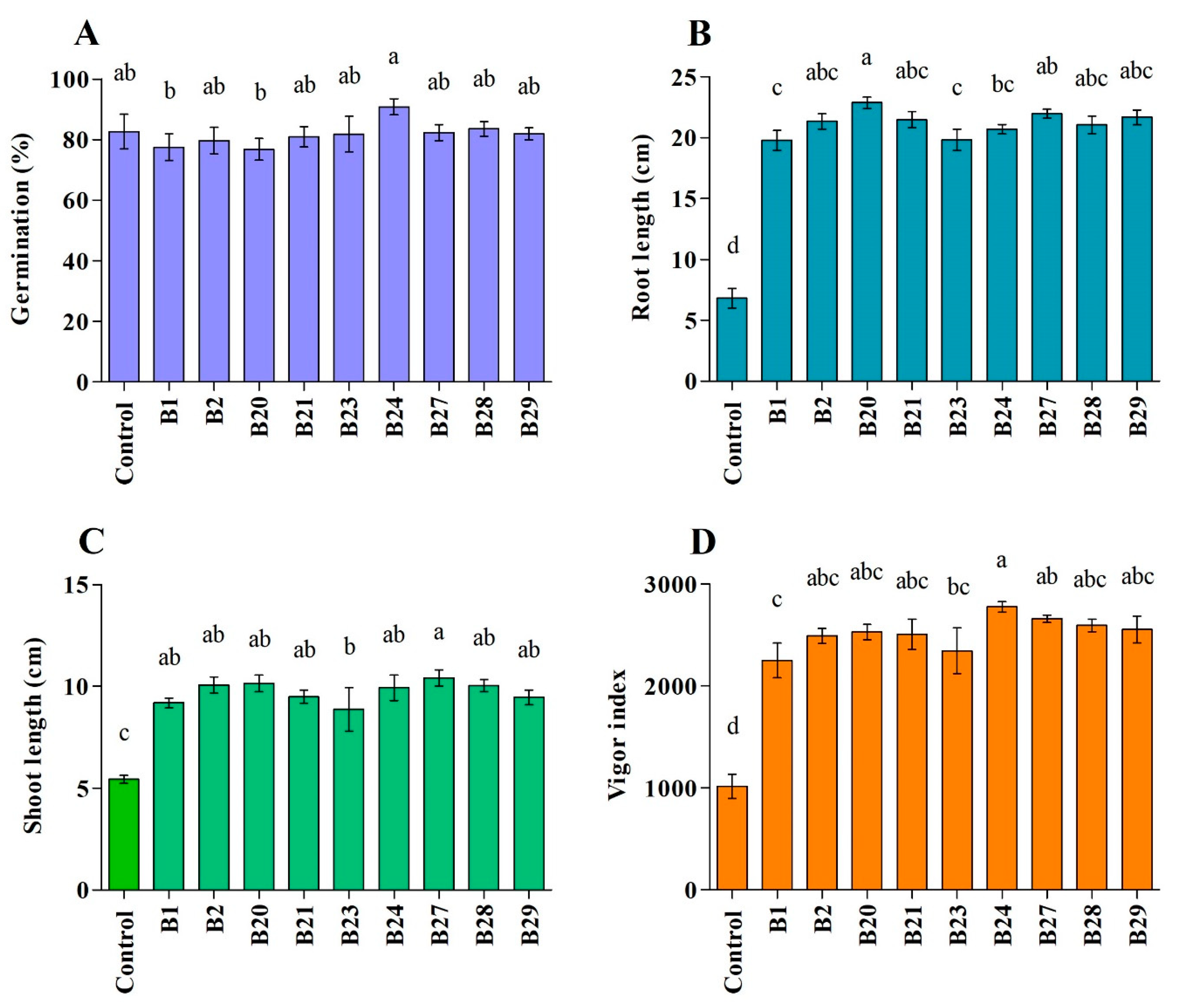
3.4.2. Effect of Bacterial Strains on Plant Growth Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 1 September 2023).
- Hashemi, L.; Golparvar, A.R.; Nasr Esfahani, M.; Golabadi, M. Correlation between cucumber genotype and resistance to damping-off disease caused by Phytophthora melonis. Biotechnol. Biotechnol. Equip. 2019, 33, 1494–1504. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Graber, E.R.; Elad, Y.; Frenkel, O. Biochar as a management tool for soilborne diseases affecting early stage nursery seedling production. Crop Prot. 2019, 120, 34–42. [Google Scholar] [CrossRef]
- Abbasi, P.A.; Renderos, W.; Fillmore, S. Soil incorporation of buckwheat as a pre-plant amendment provides control of Rhizoctonia damping-off and root rot of radish and Pythium damping-off and root rot of cucumber. Can. J. Plant Pathol. 2019, 41, 24–34. [Google Scholar] [CrossRef]
- Owen, W.G.; Jackson, B.E.; Whipker, B.E.; Fonteno, W.C.; Benson, M.D. Assessing the severity of damping-off caused by Pythium ultimum and Rhizoctonia solani in peat-based greenhouse substrates amended with pine wood chip aggregates. Acta Hortic. 2019, 1266, 27–33. [Google Scholar] [CrossRef]
- You, X.; Kimura, N.; Okura, T.; Murakami, S.; Okano, R.; Shimogami, Y.; Matsumura, A.; Tokumoto, H.; Ogata, Y.; Tojo, M. Suppressive effects of vermicomposted-bamboo powder on cucumber damping-off. Jpn. Agric. Res. Q. 2019, 53, 13–19. [Google Scholar] [CrossRef]
- Chen, W.L.; Chien, C.S. Improving productive efficiency of cucumber under greenhouse cultivation by grafting and bee pollination. Acta Hortic. 2019, 1257, 187–194. [Google Scholar] [CrossRef]
- Roberts, D.P.; McKenna, L.F.; Buyer, J.S. Consistency of control of damping-off of cucumber is improved by combining ethanol extract of Serratia marcescens with other biologically based technologies. Crop Prot. 2017, 96, 59–67. [Google Scholar] [CrossRef]
- Al-Mawaali, Q.S.; Al-Sadi, A.M.; Al-Said, F.A.; Deadman, M.L. Etiology, development and reaction of muskmelon to vine decline under arid conditions of Oman. Phytopathol. Mediterr. 2013, 52, 457–465. [Google Scholar] [CrossRef]
- Al-Mawaali, Q.S.; Al-Sadi, A.M.; Khan, A.J.; Al-Hasani, H.D.; Deadman, M.L. Response of cucurbit rootstocks to Pythium aphanidermatum. Crop Prot. 2012, 42, 64–68. [Google Scholar] [CrossRef]
- Al-Sadi, A.M.; Al-Said, F.A.; Al-Kiyumi, K.S.; Al-Mahrouqi, R.S.; Al-Mahmooli, I.H.; Deadman, M.L. Etiology and characterization of cucumber vine decline in Oman. Crop Prot. 2011, 30, 192–197. [Google Scholar] [CrossRef]
- Al-Azizi, A.; Al-Sadi, A.M.; Dietz, H.; Al-Said, F.A.; Deadman, M.L. Influence of carbon/nitrogen ratio on Pythium aphanidermatum and on Pythium-induced damping-off of radish. J. Plant Pathol. 2013, 95, 181–185. [Google Scholar]
- Halo, B.A.; Al-Yahyai, R.A.; Maharachchikumbura, S.S.N.; Al-Sadi, A.M. Talaromyces variabilis interferes with Pythium aphanidermatum growth and suppresses Pythium-induced damping-off of cucumbers and tomatoes. Sci. Rep. 2019, 9, 11255. [Google Scholar] [CrossRef] [PubMed]
- Al-Jaradi, A.; Al-Mahmooli, I.; Janke, R.; Maharachchikumbura, S.; Al-Saady, N.; Al-Sadi, A.M. Isolation and identification of pathogenic fungi and oomycetes associated with beans and cowpea root diseases in Oman. PeerJ 2018, 2018, e6064. [Google Scholar] [CrossRef] [PubMed]
- Al-Balushi, Z.M.; Agrama, H.; Al-Mahmooli, I.H.; Maharachchikumbura, S.S.N.; Al-Sadi, A.M. Development of resistance to hymexazol among Pythium species in cucumber greenhouses in Oman. Plant Dis. 2018, 102, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, A.M.; Al-Ghaithi, A.G.; Al-Balushi, Z.M.; Al-Jabri, A.H. Analysis of diversity in Pythium aphanidermatum populations from a single greenhouse reveals phenotypic and genotypic changes over 2006 to 2011. Plant Dis. 2012, 96, 852–858. [Google Scholar] [CrossRef]
- Al-Sadi, A.M.; Al-Masoodi, R.S.; Al-Ismaili, M.; Al-Mahmooli, I.H. Population structure and development of resistance to hymexazol among Fusarium solani populations from date palm, citrus and cucumber. J. Phytopathol. 2015, 163, 947–955. [Google Scholar] [CrossRef]
- Zhang, Q.; Hua, X.D.; Shi, H.Y.; Liu, J.S.; Tian, M.M.; Wang, M.H. Enantioselective bioactivity, acute toxicity and dissipation in vegetables of the chiral triazole fungicide flutriafol. J. Hazard. Mater. 2015, 284, 65–72. [Google Scholar] [CrossRef]
- Kitchen, J.L.; Van Den Bosch, F.; Paveley, N.D.; Helps, J.; Van Den Berg, F. The evolution of fungicide resistance resulting from combinations of foliar-acting systemic seed treatments and foliar-applied fungicides: A modeling analysis. PLoS ONE 2016, 11, e0161887. [Google Scholar] [CrossRef]
- Meena, R.P.; Kalariya, K.A.; Saran, P.L.; Roy, S. Efficacy of fungicides and biocontrol agents against Pythium aphanidermatum causes damping off disease in ashwagandha (Withania somnifera L. dunal). Med. Plants 2019, 11, 404–409. [Google Scholar] [CrossRef]
- Charoenrak, P.; Chamswarng, C.; Intanoo, W.; Keawprasert, N. The effects of vermicompost mixed with Trichoderma asperellum on the growth and pythium root rot of lettuces. Int. J. GEOMATE 2019, 17, 215–221. [Google Scholar] [CrossRef]
- Al-Ghafri, H.M.; Velazhahan, R.; Shahid, M.S.; Al-Sadi, A.M. Antagonistic activity of Pseudomonas aeruginosa from compost against Pythium aphanidermatum and Fusarium solani. Biocontrol. Sci. Technol. 2020, 30, 642–658. [Google Scholar] [CrossRef]
- Qulsum, M.U.; Islam, M.M.; Chowdhury, M.E.K.; Hossain, S.M.M.; Hasan, M.M. Management of bacterial wilt (Ralstonia solanacearum) of brinjal using Bacillus cereus, Trichoderma harzianum and Calotropis gigantea consortia in Bangladesh. Egyp. J. Biol. Pest Cont. 2023, 33, 74. [Google Scholar] [CrossRef]
- Ramzan, N.; Noreen, N.; Shahzad, S. Inhibition of in vitro growth of soil-borne pathogens by compost-inhabiting indigenous bacteria and fungi. Pak. J. Bot. 2014, 46, 1093–1099. [Google Scholar]
- Ramzan, N.; Noreen, N.; Perveen, Z.; Shahzad, S. Effect of seed pelleting with biocontrol agents on growth and colonisation of roots of mungbean by root-infecting fungi. J. Sci. Food Agric. 2016, 96, 3694–3700. [Google Scholar] [CrossRef] [PubMed]
- Segaran, G.; Sathiavelu, M. Fungal endophytes: A potent biocontrol agent and a bioactive metabolites reservoir. Biocatal. Agric. Biotechnol. 2019, 21, 101284. [Google Scholar] [CrossRef]
- De Silva, N.I.; Brooks, S.; Lumyong, S.; Hyde, K.D. Use of endophytes as biocontrol agents. Fungal Biol. Rev. 2019, 33, 133–148. [Google Scholar] [CrossRef]
- Al-Hinai, A.H.; Al-Sadi, A.M.; Al-Bahry, S.N.; Mothershaw, A.S.; Al-Said, F.A.; Al-Harthi, S.A.; Deadman, M.L. Isolation and characterization of Pseudomonas aeruginosa with antagonistic activity against Pythium aphanidermatum. J. Plant Pathol. 2010, 92, 653–660. [Google Scholar]
- Halo, B.A.; Al-Yahyai, R.A.; Al-Sadi, A.M. Aspergillus terreus inhibits growth and induces morphological abnormalities in Pythium aphanidermatum and suppresses Pythium-induced damping-off of cucumber. Front. Microbiol. 2018, 9, 95. [Google Scholar] [CrossRef]
- Halo, B.A.; Al-Yahyai, R.A.; Al-Sadi, A.M. Biological control of Pythium aphanidermatum-induced cucumber and radish damping-off by an endophytic fungus, Cladosporium omanense isolate 31R. Biocontrol Sci. Technol. 2021, 31, 235–251. [Google Scholar] [CrossRef]
- Kazerooni, E.A.; Al-Shibli, H.; Nasehi, A.; Al-Sadi, A.M. Endophytic Enterobacter cloacae exhibits antagonistic activity against Pythium Damping-off of cucumber. Ciência Rural 2020, 50, e20191035. [Google Scholar] [CrossRef]
- Al-Daghari, D.S.S.; Al-Abri, S.A.; Al-Mahmooli, I.H.; Al-Sadi, A.M.; Velazhahan, R. Efficacy of native antagonistic rhizobacteria in the biological control of Pythium aphanidermatum-induced damping-off of cucumber in Oman. J. Plant Pathol. 2019, 102, 305–310. [Google Scholar] [CrossRef]
- Al-Shibli, H.; Dobretsov, S.; Al-Nabhani, A.; Maharachchikumbura, S.S.N.; Rethinasamy, V.; Al-Sadi, A.M. Aspergillus terreus obtained from mangrove exhibits antagonistic activities against Pythium aphanidermatum-induced damping-off of cucumber. PeerJ 2019, 2019, 7884. [Google Scholar] [CrossRef]
- Gaya Karunasinghe, T.; Hashil Al-Mahmooli, I.; Al-Sadi, A.M.; Velazhahan, R. The effect of salt-tolerant antagonistic bacteria from tomato rhizosphere on plant growth promotion and damping-off disease suppression under salt-stress conditions. Acta Agric. Scand. Sect. B Soil Plant Sci. 2020, 70, 69–75. [Google Scholar] [CrossRef]
- Al-Badri, B.A.S.; Al-Maawali, S.S.; Al-Balushi, Z.M.; Al-Mahmooli, I.H.; Al-Sadi, A.M.; Velazhahan, R. Cyanide degradation and antagonistic potential of endophytic Bacillus subtilis strain BEB1 from Bougainvillea spectabilis Willd. All Life 2020, 13, 92–98. [Google Scholar] [CrossRef]
- Al-Daghari, D.S.S.; Al-Sadi, A.M.; Janke, R.; Al-Mahmooli, I.H.; Velazhahan, R. Potential of indigenous antagonistic rhizobacteria in the biological control of Monosporascus root rot and vine decline disease of muskmelon. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2020, 70, 371–380. [Google Scholar] [CrossRef]
- Dhiman, K.; Gupta, A.; Sharma, D.K.; Gill, N.S.; Goyal, A. A review on the medicinally important plants of the family Cucurbitaceae. Asian J. Clin. Nutr. 2012, 4, 16–26. [Google Scholar] [CrossRef]
- Yatoo, M.I.; Saxena, A.; Gopalakrishnan, A.; Alagawany, M.; Dhama, K. Promising antidiabetic drugs, medicinal plants and herbs: An update. Int. J. Pharmacol. 2017, 13, 732–745. [Google Scholar] [CrossRef]
- Hussain, A.I.; Rathore, H.A.; Sattar, M.Z.A.; Chatha, S.A.S.; Sarker, S.D.; Gilani, A.H. Citrullus colocynthis (L.) Schrad (bitter apple fruit): A review of its phytochemistry, pharmacology, traditional uses and nutritional potential. J. Ethnopharmacol. 2014, 155, 54–66. [Google Scholar] [CrossRef]
- Luu, V.T.; Weinhold, A.; Ullah, C.; Dressel, S.; Schoettner, M.; Gase, K.; Gaquerel, E.; Xu, S.; Baldwin, I.T. O-Acyl sugars protect a wild Tobacco from both native fungal pathogens and a specialist herbivore. Plant Physiol. 2017, 174, 370–386. [Google Scholar] [CrossRef]
- Lebeda, A.; Křístková, E.; Kitner, M.; Mieslerová, B.; Jemelková, M.; Pink, D.A.C. Wild Lactuca species, their genetic diversity, resistance to diseases and pests, and exploitation in lettuce breeding. Eur. J. Plant Pathol. 2014, 138, 597–640. [Google Scholar] [CrossRef]
- Ferjani, R.; Marasco, R.; Rolli, E.; Cherif, H.; Cherif, A.; Gtari, M.; Boudabous, A.; Daffonchio, D.; Ouzari, H.-I. The Date Palm Tree Rhizosphere Is a Niche for Plant Growth Promoting Bacteria in the Oasis Ecosystem. BioMed Res. Int. 2015, 2015, 153851. [Google Scholar] [CrossRef]
- Abid, L.; Smiri, M.; Federici, E.; Lievens, B.; Manai, M.; Yan, Y.; Sadfi-Zouaoui, N. Diversity of rhizospheric and endophytic bacteria isolated from dried fruit of Ficus carica. Saudi J. Biol. Sci. 2022, 29, 103398. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, H.R.M.; Argolo, C.S.; Argôlo-Filho, R.C.; Loguercio, L.L. A 16S rDNA PCR-based theoretical to actual delta approach on culturable mock communities revealed severe losses of diversity information. BMC Microbiol. 2019, 19, 74. [Google Scholar] [CrossRef]
- Anith, K.N.; Nysanth, N.S.; Natarajan, C. Novel and rapid agar plate methods for in vitro assessment of bacterial biocontrol isolates’ antagonism against multiple fungal phytopathogens. Lett. Appl. Microbiol. 2021, 73, 229–236. [Google Scholar] [CrossRef]
- Vinayarani, G.; Prakash, H. Growth promoting rhizospheric and endophytic bacteria from Curcuma longa L. as biocontrol agents against rhizome rot and leaf blight diseases. Plant Pathol. J. 2018, 34, 218. [Google Scholar] [CrossRef] [PubMed]
- KC, B.M.; Gauchan, D.P.; Khanal, S.N.; Lamichhane, J. Quantification of indole-3-acetic acid from Bambusa tulda Roxb. seedlings using high performance liquid chromatography. Afr. J. Biotechnol. 2020, 19, 781–788. [Google Scholar] [CrossRef]
- International Seed Testing Association (ISTA). International Rules of Seed Testing. Seed Sci. Technol. 2005, 15, 1–9. [Google Scholar]
- Lee, D.G.; Lee, J.M.; Choi, C.G.; Lee, H.; Moon, J.C.; Chung, N. Effect of plant growth-promoting rhizobacterial treatment on growth and physiological characteristics of Triticum aestivum L. under salt stress. Appl. Biol. Chem. 2021, 64, 89. [Google Scholar] [CrossRef]
- Philosoph, A.M.; Dombrovsky, A.; Elad, Y.; Koren, A.; Frenkel, O. Insight into late wilting disease of cucumber demonstrates the complexity of the phenomenon in fluctuating environments. Plant Dis. 2019, 103, 2877–2883. [Google Scholar] [CrossRef]
- Philosoph, A.M.; Dombrovsky, A.; Elad, Y.; Jaiswal, A.K.; Koren, A.; Lachman, O.; Frenkel, O. Combined infection with Cucumber green mottle mosaic virus and Pythium species causes extensive collapse in cucumber plants. Plant Dis. 2018, 102, 753–759. [Google Scholar] [CrossRef]
- Klochkova, T.A.; Jung, S.; Kim, G.H. Host range and salinity tolerance of Pythium porphyrae may indicate its terrestrial origin. J. Appl. Phycol. 2017, 29, 371–379. [Google Scholar] [CrossRef]
- Morris, K.A.; Langston, D.B.; Dutta, B.; Davis, R.F.; Timper, P.; Noe, J.P.; Dickson, D.W. Evidence for a disease complex between Pythium aphanidermatum and root-knot nematodes in cucumber. Plant Health Prog. 2016, 17, 200–201. [Google Scholar] [CrossRef]
- Abdalmoohsin, R.G.; Alhumairy, Y.N.; Abood, N.T.; Lahuf, A.A. Bioefficacy of Pseudomonas fluorescens and Saccharomyces cerevisiae against Pythium aphanidermatum under laboratory and greenhouse aquaculture conditions in cucumber plants, Cucumis sativus. Biopestic. Int. 2019, 15, 15–22. [Google Scholar]
- Saberi-Riseh, R.; Fathi, F. Biocontrol of Fusarium oxysporum in cucumber by some antagonist bacteria under drought stress. J. Crop Prot. 2018, 7, 375–385. [Google Scholar]
- Prabhukarthikeyan, S.R.; Keerthana, U.; Raguchander, T. Antibiotic-producing Pseudomonas fluorescens mediates rhizome rot disease resistance and promotes plant growth in turmeric plants. Microbiol Res. 2018, 210, 65–73. [Google Scholar] [CrossRef]
- Abo-Zaid, G.A.; Wagih, E.E.; Matar, S.M.; Ashmawy, N.A.; Hafez, E.E. Scaling-up production of pyocyanin from Pseudomonas aeruginosa JY21 as biocontrol agent against certain plant pathogenic fungi. Int. J. ChemTech Res. 2015, 8, 213–224. [Google Scholar]
- Islam, M.T.; Hashidoko, Y.; Deora, A.; Ito, T.; Tahara, S. Suppression of damping-off disease in host plants by the rhizoplane bacterium Lysobacter sp. strain SB-K88 is linked to plant colonization and antibiosis against soilborne peronosporomycetes. Appl. Environ. Microbiol. 2005, 71, 3786–3796. [Google Scholar] [CrossRef]
- Janahiraman, V.; Anandham, R.; Kwon, S.W.; Sundaram, S.; Pandi, V.K.; Krishnamoorthy, R.; Kim, K.; Samaddar, S.; Sa, T. Control of wilt and rot pathogens of tomato by antagonistic pink pigmented facultative methylotrophic Delftia lacustris and Bacillus spp. Front. Plant Sci. 2016, 7, 1626. [Google Scholar] [CrossRef]
- Kipngeno, P.; Losenge, T.; Maina, N.; Kahangi, E.; Juma, P. Efficacy of Bacillus subtilis and Trichoderma asperellum against Pythium aphanidermatum in tomatoes. Biol. Control 2015, 90, 92–95. [Google Scholar] [CrossRef]
- Khabbaz, S.E.; Zhang, L.; Cáceres, L.A.; Sumarah, M.; Wang, A.; Abbasi, P.A. Characterisation of antagonistic Bacillus and Pseudomonas strains for biocontrol potential and suppression of damping-off and root rot diseases. Ann. App. Biol. 2015, 166, 456–471. [Google Scholar] [CrossRef]
- Khabbaz, S.E.; Abbasi, P.A. Isolation, characterization, and formulation of antagonistic bacteria for the management of seedlings damping-off and root rot disease of cucumber. Can. J. Microbiol. 2014, 60, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Shahzad, R.; Khan, A.L.; Halo, B.A.; Al-Yahyai, R.; Al-Harrasi, A.; Al-Rawahi, A.; Lee, I.J. Endophytic bacterial diversity of Avicennia marina helps to confer resistance against salinity stress in Solanum lycopersicum. J. Plant Interact. 2017, 12, 312–322. [Google Scholar] [CrossRef]
- Selvakumar, G.; Joshi, P.; Nazim, S.; Mishra, P.K.; Kundu, S.; Gupta, H.S. Exiguobacterium acetylicum strain 1P (MTCC 8707) a novel bacterial antagonist from the North Western Indian Himalayas. World J. Microbiol. Biotechnol. 2009, 25, 131–137. [Google Scholar] [CrossRef]
- Al-Hussini, H.S.; Al-Rawahi, A.Y.; Al-Marhoon, A.A.; Al-Abri, S.A.; Al-Mahmooli, I.H.; Al-Sadi, A.M.; Velazhahan, R. Biological control of damping-off of tomato caused by Pythium aphanidermatum by using native antagonistic rhizobacteria isolated from Omani soil. J. Plant Pathol. 2019, 101, 315–322. [Google Scholar] [CrossRef]
- Dhaouadi, S.; Rouissi, W.; Mougou-Hamdane, A.; Nasraoui, B. Evaluation of biocontrol potential of Achromobacter xylosoxidans against Fusarium wilt of melon. Eur. J. Plant Pathol. 2019, 154, 179–188. [Google Scholar] [CrossRef]
- Wang, K.; Li, Y.; Wu, Y.; Qiu, Z.; Ding, Z.; Wang, X.; Chen, W.; Wang, R.; Fu, F.; Rensing, C.; et al. Improved grain yield and lowered arsenic accumulation in rice plants by inoculation with arsenite-oxidizing Achromobacter xylosoxidans GD03. Ecotoxicol. Environ. Saf. 2020, 206, 111229. [Google Scholar] [CrossRef]
- Danish, S.; Zafar-ul-Hye, M.; Fahad, S.; Saud, S.; Brtnicky, M.; Hammerschmiedt, T.; Datta, R. Drought stress alleviation by ACC deaminase producing Achromobacter xylosoxidans and Enterobacter cloacae, with and without timber waste biochar in maize. Sustainabilty 2020, 12, 6286. [Google Scholar] [CrossRef]
- Ramadan, W.A.; Soliman, G.M. Effect of different applications of bio-agent Achromobacter xylosoxidans against Meloidogyne incognita and gene expression in infected eggplant. Jordan J. Biol. Sci. 2020, 13, 363–370. [Google Scholar]
- Jiang, L.; Jeong, J.C.; Lee, J.S.; Park, J.M.; Yang, J.W.; Lee, M.H.; Choi, S.H.; Kim, C.Y.; Kim, D.H.; Kim, S.W.; et al. Potential of Pantoea dispersa as an effective biocontrol agent for black rot in sweet potato. Sci. Rep. 2019, 9, 16354. [Google Scholar] [CrossRef]
- Maisuria, V.B.; Gohel, V.; Mehta, A.N.; Patel, R.R.; Chhatpar, H.S. Biological control of Fusarium wilt of pigeonpea by Pantoea dispersa, a field assessment. Ann. Microbiol. 2008, 58, 411–419. [Google Scholar] [CrossRef]
- Li, Y.; Guo, P.Y.; Sun, J.G. Isolation, identification, phylogeny and growth promoting characteristics of endophytic diazotrophs from tuber and root crops. Sci. Agric. Sin. 2017, 50, 104–122. [Google Scholar] [CrossRef]
- Shahzad, R.; Waqas, M.; Khan, A.L.; Al-Hosni, K.; Kang, S.M.; Seo, C.W.; Lee, I.J. Indoleacetic acid production and plant growth promoting potential of bacterial endophytes isolated from rice (Oryza sativa L.) seeds. Acta Biol. Hung. 2017, 68, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Selvakumar, G.; Kundu, S.; Joshi, P.; Nazim, S.; Gupta, A.D.; Mishra, P.K.; Gupta, H.S. Characterization of a cold-tolerant plant growth-promoting bacterium Pantoea dispersa 1A isolated from a sub-alpine soil in the North Western Indian Himalayas. World J. Microbiol. Biotechnol. 2008, 24, 955–960. [Google Scholar] [CrossRef]
- Moustafa, S.M.N. Influence of Pythium oligandrum on mycelia and production of zoo-and oo-spores of two phytopathogenic Pythium spp. J. Pure Appl. Microbiol. 2019, 13, 233–240. [Google Scholar] [CrossRef]
- Culebro-Ricaldi, J.M.; Ruíz-Valdiviezo, V.M.; Rodríguez-Mendiola, M.A.; Ávila-Miranda, M.E.; Gutiérrez-Miceli, F.A.; Cruz-Rodríguez, R.I.; Dendooven, L.; Montes-Molina, J.A. Antifungal properties of Beauveria bassiana strains against Fusarium oxysporum f. Sp. Lycopersici race 3 in tomato crop. J. Environ. Biol. 2017, 38, 821–827. [Google Scholar] [CrossRef]
- Kunova, A.; Bonaldi, M.; Saracchi, M.; Pizzatti, C.; Chen, X.; Cortesi, P. Selection of Streptomyces against soil borne fungal pathogens by a standardized dual culture assay and evaluation of their effects on seed germination and plant growth. BMC Microbiol. 2016, 16, 272. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, Y.; Chen, K.; Wang, Y.; Hu, J.; Wei, Y.; Yang, H. Trichoderma cyanodichotomus sp. nov., a new soil-inhabiting species with a potential for biological control. Can. J. Microbiol. 2018, 64, 1020–1029. [Google Scholar] [CrossRef]
- Wu, Q.; Sun, R.; Ni, M.; Yu, J.; Li, Y.; Yu, C.; Dou, K.; Ren, J.; Chen, J. Identification of a novel fungus, Trichoderma asperellum GDFS1009, and comprehensive evaluation of its biocontrol efficacy. PLoS ONE 2017, 12, e0179957. [Google Scholar] [CrossRef]
- Yin, J.; Yuan, L. Phytophthora disease control and growth promotion of pepper by Pythium oligandrum. Acta Hortic. Sin. 2017, 44, 2327–2337. [Google Scholar] [CrossRef]
- Wang, S.; Liang, Y.; Shen, T.; Yang, H.; Shen, B. Biological characteristics of Streptomyces albospinus CT205 and its biocontrol potential against cucumber Fusarium wilt. Biocontrol Sci. Technol. 2016, 26, 951–963. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Yu, C.; Dou, K.; Wang, M.; Li, Y.; Chen, J. Synergistic effect of Trichoderma-derived antifungal metabolites and cell wall degrading enzymes on enhanced biocontrol of Fusarium oxysporum f. sp. cucumerinum. Biol. Control 2016, 94, 37–46. [Google Scholar] [CrossRef]
- Mayo, S.; Gutiérrez, S.; Cardoza, R.E.; Hermosa, R.; Monte, E.; Casquero, P.A. Trichoderma species as biocontrol agents in legumes. In Legumes for Global Food Security; Jimenez-Lopez, J.C., Clemente, A., Eds.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2017; pp. 73–100. [Google Scholar]
- Inglis, G.D.; Kawchuk, L.M. Comparative degradation of oomycete, ascomycete, and basidiomycete cell walls by mycoparasitic and biocontrol fungi. Can. J. Microbiol. 2002, 48, 60–70. [Google Scholar] [CrossRef]
- Gu, Y.; Wang, J.; Xia, Z.; Wei, H.L. Characterization of a versatile plant growth-promoting rhizobacterium Pseudomonas mediterranea strain s58. Microorg. 2020, 8, 334. [Google Scholar] [CrossRef]
- Ullah, H.; Yasmin, H.; Mumtaz, S.; Jabeen, Z.; Naz, R.; Nosheen, A.; Hassan, M.N. Multitrait Pseudomonas spp. isolated from monocropped wheat (Triticum aestivum) suppress Fusarium root and crown rot. Phytopathology 2020, 110, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, C.; Banerjee, S.; Acharya, U.; Mitra, A.; Mallick, I.; Haldar, A.; Haldar, S.; Ghosh, A.; Ghosh, A. Evaluation of plant growth promotion properties and induction of antioxidative defense mechanism by tea rhizobacteria of Darjeeling, India. Sci. Rep. 2020, 10, 15536. [Google Scholar] [CrossRef]
- Rincón-Molina, C.I.; Martínez-Romero, E.; Ruiz-Valdiviezo, V.M.; Velázquez, E.; Ruiz-Lau, N.; Rogel-Hernández, M.A.; Villalobos-Maldonado, J.J.; Rincón-Rosales, R. Plant growth-promoting potential of bacteria associated to pioneer plants from an active volcanic site of Chiapas (Mexico). Appl. Soil Ecol. 2020, 146, 103390. [Google Scholar] [CrossRef]
- Aeron, A.; Maheshwari, D.K.; Meena, V.S. Endophytic bacteria promote growth of the medicinal legume Clitoria ternatea L. by chemotactic activity. Arch. Microbiol. 2020, 202, 1049–1058. [Google Scholar] [CrossRef]
- Singh, T.B.; Sahai, V.; Ali, A.; Prasad, M.; Yadav, A.; Shrivastav, P.; Goyal, D.; Dantu, P.K. Screening and evaluation of PGPR strains having multiple PGP traits from hilly terrain. J. Appl. Biol. Biotechnol. 2020, 8, 38–44. [Google Scholar] [CrossRef]
- Minaeva, O.M.; Akimova, E.E.; Tereshchenko, N.N.; Kravets, A.V.; Zyubanova, T.I.; Apenysheva, M.V. Pseudomonads associated with soil lumbricides as promising agents in root rod biocontrol for spring grain crops. Sel’skokhozyaistvennaya Biol. 2019, 54, 91–100. [Google Scholar] [CrossRef]
- Liu, X.; Li, Q.; Li, Y.; Guan, G.; Chen, S. Paenibacillus strains with nitrogen fixation and multiple beneficial properties for promoting plant growth. PeerJ 2019, 2019, e7445. [Google Scholar] [CrossRef]
- Cui, W.; He, P.; Munir, S.; He, P.; Li, X.; Li, Y.; Wu, J.; Wu, Y.; Yang, L.; He, P.; et al. Efficacy of plant growth promoting bacteria Bacillus amyloliquefaciens B9601-Y2 for biocontrol of southern corn leaf blight. Biol. Control 2019, 139, 104080. [Google Scholar] [CrossRef]
- Jha, P.; Kumar, A. Characterization of novel plant growth promoting endophytic bacterium Achromobacter xylosoxidans from wheat plant. Microb. Ecol. 2009, 58, 179–188. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Code | Source of Isolation | Identified Species | Max. Identity (%) | GenBank Accession No. | Place of Collection |
|---|---|---|---|---|---|
| B1 | Soil | Achromobacter xylosoxidans | 99.79% | JQ724537 | Wadi Al Alaq |
| B2 | Soil | Pantoea dispersa | 100% | MN725743 | Wadi Al Alaq |
| B3 | Soil | Pseudomonas lini | 99.75% | MT102313 | Wadi Al Alaq |
| B4 | Soil | Pseudomonas plecoglossicida | 99.71% | MF574326 | Wadi Al Alaq |
| B5 | Soil | Pseudomonas aeruginosa | 100% | JQ659890 | Wadi Al Alaq |
| B6 | Soil | Pseudomonas fluorescens | 100% | HM439968 | Wadi Al Alaq |
| B7 | Soil | Rhizobium pusense | 100% | MK734334 | Wadi Al Alaq |
| B8 | Soil | Sphingobacterium spiritivorum | 100% | KR349259 | Wadi Bani Khalid |
| B9 | Soil | Pseudomonas plecoglossicida | 100% | MF574326 | Wadi Bani Khalid |
| B10 | Soil | Achromobacter xylosoxidans | 99.82% | MK537386 | Wadi Bani Khalid |
| B11 | Soil | Pseudomonas putida | 100% | KU672371 | Wadi Bani Khalid |
| B12 | Root | Bacillus aryabhattai | 100% | MT078622 | Wadi Al Alaq |
| B13 | Root | Sinorhizobium meliloti | 99.85% | MT197365 | Wadi Al Alaq |
| B14 | Root | Bacillus anthracis | 98.89% | KF601916 | Wadi Al Alaq |
| B15 | Root | Bacillus anthracis | 99.66% | KF601916 | Wadi Bani Khalid |
| B16 | Root | Unknown | Wadi Bani Khalid | ||
| B17 | Root | Achromobacter mucicolens | 99.48% | MT534143 | Wadi Bani Khalid |
| B18 | Root | Achromobacter xylosoxidans | 98.79% | MN889379 | Wadi Bani Khalid |
| B19 | Stem | Achromobacter xylosoxidans | 99.86% | MK332530 | Wadi Al Alaq |
| B20 | Stem | Unknown | Wadi Al Alaq | ||
| B21 | Stem | Bacillus cereus | 100% | MK648340 | Wadi Bani Khalid |
| B22 | Stem | Pseudomonas plecoglossicida | 99.86% | MF574326 | Wadi Bani Khalid |
| B23 | Leaf | Pseudomonas stutzeri | 97.34% | KY606628 | Wadi Al Alaq |
| B24 | Leaf | Staphylococcus gallinarum | 99.93% | MH542297 | Wadi Al Alaq |
| B25 | Leaf | Cupriavidus gilardii | 99.50% | AY860225 | Wadi Bani Khalid |
| B26 | Leaf | Achromobacter xylosoxidans | 100% | MK537386 | Wadi Bani Khalid |
| B27 | Fruit | Pantoea dispersa | 100% | MN725743 | Wadi Al Alaq |
| B28 | Fruit | Exiguobacterium indicum | 99.86% | KT986092 | Wadi Al Alaq |
| B29 | Fruit | Unknown | Wadi Bani Khalid | ||
| B30 | Fruit | Achromobacter xylosoxidans | 98% | HQ288926 | Wadi Bani Khalid |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Shuaibi, B.K.; Kazerooni, E.A.; Hussain, S.; Velazhahan, R.; Al-Sadi, A.M. Plant-Disease-Suppressive and Growth-Promoting Activities of Endophytic and Rhizobacterial Isolates Associated with Citrullus colocynthis. Pathogens 2023, 12, 1275. https://doi.org/10.3390/pathogens12111275
Al-Shuaibi BK, Kazerooni EA, Hussain S, Velazhahan R, Al-Sadi AM. Plant-Disease-Suppressive and Growth-Promoting Activities of Endophytic and Rhizobacterial Isolates Associated with Citrullus colocynthis. Pathogens. 2023; 12(11):1275. https://doi.org/10.3390/pathogens12111275
Chicago/Turabian StyleAl-Shuaibi, Badriya Khalfan, Elham Ahmed Kazerooni, Shah Hussain, Rethinasamy Velazhahan, and Abdullah Mohammed Al-Sadi. 2023. "Plant-Disease-Suppressive and Growth-Promoting Activities of Endophytic and Rhizobacterial Isolates Associated with Citrullus colocynthis" Pathogens 12, no. 11: 1275. https://doi.org/10.3390/pathogens12111275