

In Situ Hybridization (RNAscope) Detection of Bluetongue Virus Serotypes 10 and 17 in Experimentally Co-Infected Culicoides sonorensis
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culicoides Sonorensis and Viruses
2.2. C. sonorensis Infection and Maintenance
2.3. In Situ Hybridization: RNAscope 2.5 HD Chromogenic Duplex Detection
2.4. Image Capture
3. Results
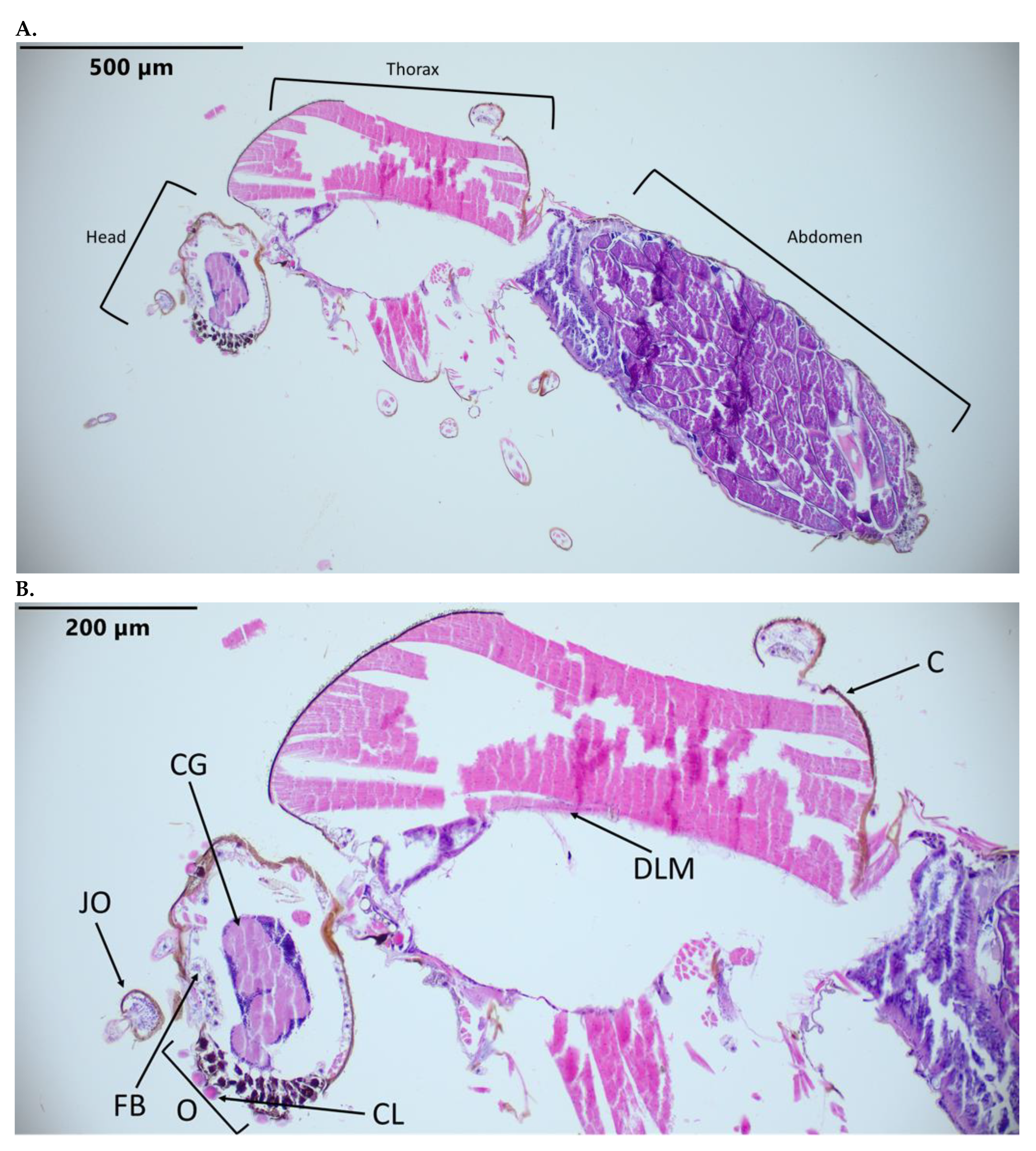
3.1. Identification of C. sonorensis Structures in Hematoxylin-and-Eosin-Stained Slides
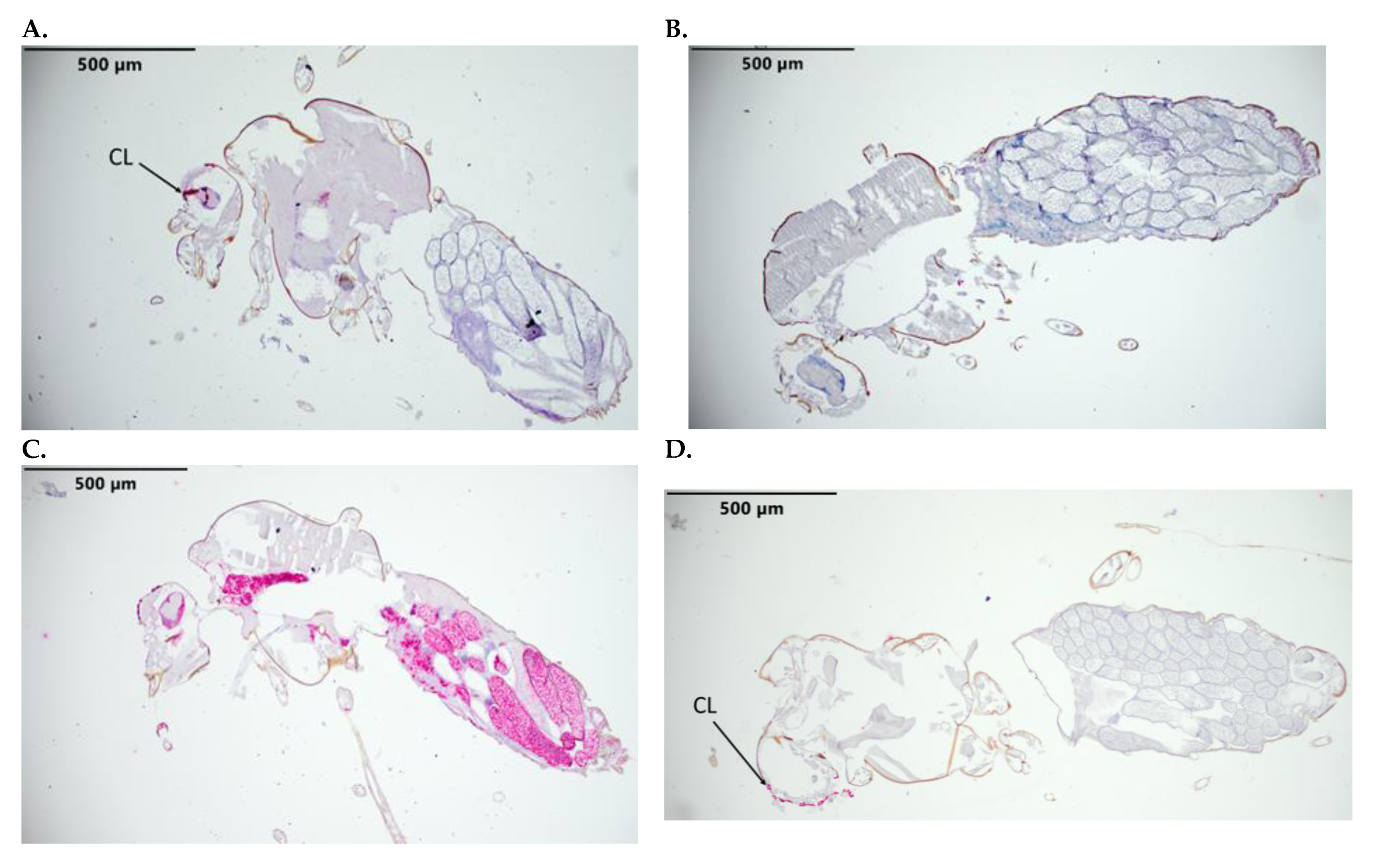
3.2. Control Probes Indicate Successful RNAscope Hybridization and Probe Detection
3.3. Detection and Distribution of BTV-10 and BTV-17 in Single Infection Treatment Groups
3.4. Detection and Distribution of BTV-10 and BTV-17 in Co-Infection Treatment Group
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gibbs, E.P. Bluetongue—An Analysis of Current Problems, with Particular Reference to Importation of Ruminants to the United States. J. Am. Vet. Med. Assoc. 1983, 182, 1190–1194. [Google Scholar] [PubMed]
- Maclachlan, N.J.; Mayo, C.E.; Daniels, P.W.; Savini, G.; Zientara, S.; Gibbs, E.P.J. Bluetongue. Rev. Sci. Tech. 2015, 34, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Mellor, P.S.; Boorman, J.; Baylis, M. Culicoides Biting Midges: Their Role as Arbovirus Vectors. Annu. Rev. Entomol. 2000, 45, 307–340. [Google Scholar] [CrossRef]
- Gibbs, E.P.; Greiner, E.C. The Epidemiology of Bluetongue. Comp. Immunol. Microbiol. Infect. Dis. 1994, 17, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Schirtzinger, E.E.; Jasperson, D.C.; Ostlund, E.N.; Johnson, D.J.; Wilson, W.C. Recent US Bluetongue Virus Serotype 3 Isolates Found Outside of Florida Indicate Evidence of Reassortment with Co-Circulating Endemic Serotypes. J. Gen. Virol. 2018, 99, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Verdezoto, J.; Breard, E.; Viarouge, C.; Quenault, H.; Lucas, P.; Sailleau, C.; Zientara, S.; Augot, D.; Zapata, S. Novel Serotype of Bluetongue Virus in South America and First Report of Epizootic Haemorrhagic Disease Virus in Ecuador. Transbound. Emerg. Dis. 2018, 65, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Brenner, J.; Oura, C.; Asis, I.; Maan, S.; Elad, D.; Maan, N.; Friedgut, O.; Nomikou, K.; Rotenberg, D.; Bumbarov, V.; et al. Multiple Serotypes of Bluetongue Virus in Sheep and Cattle, Israel. Emerg. Infect. Dis. 2010, 16, 2003–2004. [Google Scholar] [CrossRef]
- Golender, N.; Eldar, A.; Ehrlich, M.; Khinich, Y.; Kenigswald, G.; Varsano, J.S.; Ertracht, S.; Abramovitz, I.; Assis, I.; Shlamovitz, I.; et al. Emergence of a Novel Reassortant Strain of Bluetongue Serotype 6 in Israel, 2017: Clinical Manifestations of the Disease and Molecular Characterization. Viruses 2019, 11, 633. [Google Scholar] [CrossRef]
- White, J.R.; Williams, D.T.; Davies, K.; Wang, J.; Chen, H.; Certoma, A.; Davis, S.S.; Weir, R.P.; Melville, L.F.; Eagles, D. Bluetongue Virus Serotype 12 Enters Australia—A Further Incursion of Novel Western Lineage Genome Segments. J. Gen. Virol. 2021, 102, 001536. [Google Scholar] [CrossRef]
- Purse, B.V.; Mellor, P.S.; Rogers, D.J.; Samuel, A.R.; Mertens, P.P.C.; Baylis, M. Climate Change and the Recent Emergence of Bluetongue in Europe. Nat. Rev. Microbiol. 2005, 3, 171–181. [Google Scholar] [CrossRef]
- Purse, B.V.; Brown, H.E.; Harrup, L.; Mertens, P.P.C.; Rogers, D.J. Invasion of Bluetongue and Other Orbivirus Infections into Europe: The Role of Biological and Climatic Processes. Rev. Sci. Tech. 2008, 27, 427–442. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.A.; Higgs, S. Impact of Climate Change and Other Factors on Emerging Arbovirus Diseases. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 109–121. [Google Scholar] [CrossRef]
- Mertens, P.P.; Brown, F.; Sangar, D. V Assignment of the Genome Segments of Bluetongue Virus Type 1 to the Proteins Which They Encode. Virology 1984, 135, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Roy, P. The Molecular Biology of Bluetongue Virus Replication. Virus Res. 2014, 182, 5–20. [Google Scholar] [CrossRef]
- Matthijnssens, J.; Attoui, H.; Bányai, K.; Brussaard, C.P.D.; Danthi, P.; del Vas, M.; Dermody, T.S.; Duncan, R.; Fāng, Q.; Johne, R.; et al. ICTV Virus Taxonomy Profile: Sedoreoviridae 2022. J. Gen. Virol. 2022, 103, 001782. [Google Scholar] [CrossRef] [PubMed]
- Huismans, H.; van der Walt, N.T.; Cloete, M.; Erasmus, B.J. Isolation of a Capsid Protein of Bluetongue Virus That Induces a Protective Immune Response in Sheep. Virology 1987, 157, 172–179. [Google Scholar] [CrossRef]
- Maan, N.S.; Maan, S.; Belaganahalli, M.N.; Ostlund, E.N.; Johnson, D.J.; Nomikou, K.; Mertens, P.P.C. Identification and Differentiation of the Twenty Six Bluetongue Virus Serotypes by RT-PCR Amplification of the Serotype-Specific Genome Segment 2. PLoS ONE 2012, 7, e32601. [Google Scholar] [CrossRef] [PubMed]
- Ries, C.; Vögtlin, A.; Hüssy, D.; Jandt, T.; Gobet, H.; Hilbe, M.; Burgener, C.; Schweizer, L.; Häfliger-Speiser, S.; Beer, M.; et al. Putative Novel Atypical BTV Serotype “36” Identified in Small Ruminants in Switzerland. Viruses 2021, 13, 721. [Google Scholar] [CrossRef]
- Savini, G.; Puggioni, G.; Meloni, G.; Marcacci, M.; Di Domenico, M.; Rocchigiani, A.M.; Spedicato, M.; Oggiano, A.; Manunta, D.; Teodori, L.; et al. Novel Putative Bluetongue Virus in Healthy Goats from Sardinia, Italy. Infect. Genet. Evol. 2017, 51, 108–117. [Google Scholar] [CrossRef]
- Schulz, C.; Bréard, E.; Sailleau, C.; Jenckel, M.; Viarouge, C.; Vitour, D.; Palmarini, M.; Gallois, M.; Höper, D.; Hoffmann, B.; et al. Bluetongue Virus Serotype 27: Detection and Characterization of Two Novel Variants in Corsica, France. J. Gen. Virol. 2016, 97, 2073–2083. [Google Scholar] [CrossRef]
- Cavany, S.M.; Barbera, C.; Carpenter, M.; Rodgers, C.; Sherman, T.; Stenglein, M.; Mayo, C.; Perkins, T.A. Modeling Cellular Co-Infection and Reassortment of Bluetongue Virus in Culicoides Midges. Virus Evol. 2022, 8, veac094. [Google Scholar] [CrossRef] [PubMed]
- Lowen, A.C. It’s in the Mix: Reassortment of Segmented Viral Genomes. PLoS Pathog. 2018, 14, e1007200. [Google Scholar] [CrossRef] [PubMed]
- Sanders, C.; Veronesi, E.; Rajko-Nenow, P.; Mertens, P.P.C.; Batten, C.; Gubbins, S.; Carpenter, S.; Darpel, K. Field-Reassortment of Bluetongue Virus Illustrates Plasticity of Virus Associated Phenotypic Traits in the Arthropod Vector and Mammalian Host In Vivo. J. Virol. 2022, 96, e0053122. [Google Scholar] [CrossRef] [PubMed]
- Jacquot, M.; Rao, P.P.; Yadav, S.; Nomikou, K.; Maan, S.; Jyothi, Y.K.; Reddy, N.; Putty, K.; Hemadri, D.; Singh, K.P.; et al. Contrasting Selective Patterns across the Segmented Genome of Bluetongue Virus in a Global Reassortment Hotspot. Virus Evol. 2019, 5, vez027. [Google Scholar] [CrossRef] [PubMed]
- Lumley, S.; Hunter, L.; Emery, K.; Hewson, R.; Fooks, A.R.; Horton, D.L.; Johnson, N. Detection of Rift Valley Fever Virus RNA in Formalin-Fixed Mosquitoes by In Situ Hybridization (RNAscope®). Viruses 2021, 13, 1079. [Google Scholar] [CrossRef] [PubMed]
- Ragan, I.K.; Schuck, K.N.; Upreti, D.; Odendaal, L.; Richt, J.A.; Trujillo, J.D.; Wilson, W.C.; Davis, A.S. Rift Valley Fever Viral RNA Detection by In Situ Hybridization in Formalin-Fixed, Paraffin-Embedded Tissues. Vector Borne Zoonotic Dis. 2019, 19, 553–556. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.H.; Foster, N.M. Relevance of Laboratory Colonies of the Vector in Arbovirus Research--Culicoides Variipennis and Bluetongue. Am. J. Trop. Med. Hyg. 1978, 27, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Nayduch, D.; Cohnstaedt, L.W.; Saski, C.; Lawson, D.; Kersey, P.; Fife, M.; Carpenter, S. Studying Culicoides Vectors of BTV in the Post-Genomic Era: Resources, Bottlenecks to Progress and Future Directions. Virus Res. 2014, 182, 43–49. [Google Scholar] [CrossRef]
- MCKERCHER, D.G.; MCGOWAN, B.; HOWARTH, J.A.; SAITO, J.K. A Preliminary Report on the Isolation and Identification of the Bluetongue Virus from Sheep in California. J. Am. Vet. Med. Assoc. 1953, 122, 300–301. [Google Scholar]
- Kopanke, J.H. Characterizing The Genetic Evolution of Endemic Bluetongue Virus Strains. Ph.D. Thesis, Colorado State University, Fort Collins, CO, USA, 2019. [Google Scholar]
- Kopanke, J.; Lee, J.; Stenglein, M.; Carpenter, M.; Cohnstaedt, L.W.; Wilson, W.C.; Mayo, C. Exposure of Culicoides sonorensis to Enzootic Strains of Bluetongue Virus Demonstrates Temperature- and Virus-Specific Effects on Virogenesis. Viruses 2021, 13, 1016. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty Per Cent Endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Nayduch, D.; Lee, M.B.; Saski, C.A. The Reference Transcriptome of the Adult Female Biting Midge (Culicoides sonorensis) and Differential Gene Expression Profiling during Teneral, Blood, and Sucrose Feeding Conditions. PLoS ONE 2014, 9, e98123. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.L.; Vandyke, K.A.; Letchworth, G.J.; Drolet, B.S.; Hanekamp, T.; Wilson, W.C. Midgut and Salivary Gland Transcriptomes of the Arbovirus Vector Culicoides sonorensis (Diptera: Ceratopogonidae). Insect Mol. Biol. 2005, 14, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Morales-Hojas, R.; Hinsley, M.; Armean, I.M.; Silk, R.; Harrup, L.E.; Gonzalez-Uriarte, A.; Veronesi, E.; Campbell, L.; Nayduch, D.; Saski, C.; et al. The Genome of the Biting Midge Culicoides sonorensis and Gene Expression Analyses of Vector Competence for Bluetongue Virus. BMC Genom. 2018, 19, 624. [Google Scholar] [CrossRef]
- Anderson, G.A.; Phillips, D.L.; Waldvogel, A.S.; Osburn, B.I. Detection of Bluetongue Virus in Bovine Fetuses Using the Avidin-Biotin Complex Immunoperoxidase Method. J. Vet. Diagn. Investig. 1989, 1, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Drolet, B.S.; van Rijn, P.; Howerth, E.W.; Beer, M.; Mertens, P.P. A Review of Knowledge Gaps and Tools for Orbivirus Research. Vector Borne Zoonotic Dis. 2015, 15, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Lean, F.Z.X.; Payne, J.; Harper, J.; Devlin, J.; Williams, D.T.; Bingham, J. Evaluation of Bluetongue Virus (BTV) Antibodies for the Immunohistochemical Detection of BTV and Other Orbiviruses. Microorganisms 2020, 8, 1207. [Google Scholar] [CrossRef]
- Fu, H.; Leake, C.J.; Mertens, P.P.; Mellor, P.S. The Barriers to Bluetongue Virus Infection, Dissemination and Transmission in the Vector, Culicoides Variipennis (Diptera: Ceratopogonidae). Arch. Virol. 1999, 144, 747–761. [Google Scholar] [CrossRef]
- Molina Palacios, D.; Stoffolano, J.G.; Fausto, A.M.; Gambellini, G.; Burand, J. The Effect of the Hypertrophy Virus (MdSGHV) on the Ultrastructure of the Salivary Glands of Musca domestica (Diptera: Muscidae). J. Insect Sci. 2021, 21, 12. [Google Scholar] [CrossRef]
- Labadie, T.; Roy, P. A Non-Enveloped Arbovirus Released in Lysosome-Derived Extracellular Vesicles Induces Super-Infection Exclusion. PLoS Pathog. 2020, 16, e1009015. [Google Scholar] [CrossRef]
- Gaburro, J.; Bhatti, A.; Harper, J.; Jeanne, I.; Dearnley, M.; Green, D.; Nahavandi, S.; Paradkar, P.N.; Duchemin, J.-B. Neurotropism and Behavioral Changes Associated with Zika Infection in the Vector Aedes Aegypti. Emerg. Microbes Infect. 2018, 7, 68. [Google Scholar] [CrossRef] [PubMed]
- Chotiwan, N.; Andre, B.G.; Sanchez-Vargas, I.; Islam, M.N.; Grabowski, J.M.; Hopf-Jannasch, A.; Gough, E.; Nakayasu, E.; Blair, C.D.; Belisle, J.T.; et al. Dynamic Remodeling of Lipids Coincides with Dengue Virus Replication in the Midgut of Aedes Aegypti Mosquitoes. PLoS Pathog. 2018, 14, e1006853. [Google Scholar] [CrossRef] [PubMed]
- Valles, S.M. Solenopsis Invicta Virus 3 Infection Alters Foraging Behavior in Its Host Solenopsisinvicta. Virology 2023, 581, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Cross, S.T.; Brehm, A.L.; Dunham, T.J.; Rodgers, C.P.; Keene, A.H.; Borlee, G.I.; Stenglein, M.D. Galbut Virus Infection Minimally Influences Drosophila Melanogaster Fitness Traits in a Strain and Sex-Dependent Manner. Viruses 2023, 15, 539. [Google Scholar] [CrossRef] [PubMed]
- Visconti, V.; Eychenne, M.; Darboux, I. Modulation of Antiviral Immunity by the Ichnovirus HdIV in Spodoptera Frugiperda. Mol. Immunol. 2019, 108, 89–101. [Google Scholar] [CrossRef] [PubMed]
- McDermott, E.G.; Mayo, C.E.; Gerry, A.C.; Laudier, D.; MacLachlan, N.J.; Mullens, B.A. Bluetongue Virus Infection Creates Light Averse Culicoides Vectors and Serious Errors in Transmission Risk Estimates. Parasit. Vectors 2015, 8, 460. [Google Scholar] [CrossRef] [PubMed]
- Mills, M.K.; Ruder, M.G.; Nayduch, D.; Michel, K.; Drolet, B.S. Dynamics of Epizootic Hemorrhagic Disease Virus Infection within the Vector, Culicoides sonorensis (Diptera: Ceratopogonidae). PLoS ONE 2017, 12, e0188865. [Google Scholar] [CrossRef] [PubMed]
- Nayduch, D.; Shankar, V.; Mills, M.K.; Robl, T.; Drolet, B.S.; Ruder, M.G.; Scully, E.D.; Saski, C.A. Transcriptome Response of Female Culicoides sonorensis Biting Midges (Diptera: Ceratopogonidae) to Early Infection with Epizootic Hemorrhagic Disease Virus (EHDV-2). Viruses 2019, 11, 473. [Google Scholar] [CrossRef]
- Osborne, C.J.; Mayo, C.E.; Mullens, B.A.; McDermott, E.G.; Gerry, A.C.; Reisen, W.K.; MacLachlan, N.J. Lack of Evidence for Laboratory and Natural Vertical Transmission of Bluetongue Virus in Culicoides sonorensis (Diptera: Ceratopogonidae). J. Med. Entomol. 2015, 52, 274–277. [Google Scholar] [CrossRef]
- Al-Doaiss, A.A.; Al-Mekhlafi, F.A.; Abutaha, N.M.; Al-Keridis, L.A.; Shati, A.A.; Al-Kahtani, M.A.; Alfaifi, M.Y. Morphological, Histological and Ultrastructural Characterisation of Culex pipiens (Diptera: Culicidae) Larval Midgut. African Entomol. 2021, 29, 274–288. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infection Treatment Group | Detection of BTV-10 Segment 2 Only | Detection Of BTV-17 Segment 2 Only | Detection of BTV-10 and BTV-17 Segment 2 | No Detection |
|---|---|---|---|---|
| BTV-10 | 2/15 (13.3%) | 0/15 (0%) | 0/15 (0%) | 13/15 (86.7%) |
| BTV-17 | 0/13 (0%) | 8/13 (61.5%) | 0/13 (0%) | 5/13 (38.5%) |
| BTV-10 and BTV-17 | 1/14 (7.1%) | 3/14 (21.4%) | 3/14 (21.4%) | 7/14 (50.0%) |
| BTV-10 Infection Treatment Group | BTV-17 Infection Treatment Group | ||
|---|---|---|---|
| Detection of BTV-10 | Detection of BTV-17 | ||
| Number of Midges with BTV Detection | 2 Out of 15 Midges | 8 Out of 13 Midges | |
| Dissemination Pattern | 1 Midge with BTV-10 Dissemination | 1 Midge with BTV-10 Localized to Midgut | 8 Midges with BTV-17 Dissemination |
| Tissue | |||
| Johnston’s organ | n/a | n/a | 3/3 (++) to (+++) |
| ommatidia | n/a | n/a | 7/7 (+++) to (++++) |
| cerebral ganglion | n/a | n/a | 7/7 (++) to (+++) (periphery) |
| salivary gland | 1/1 (+) | 0/1 | 5/6 (++) to (+++) |
| dorsal longitudinal muscle | 1/1 (+) | 0/1 | 5/8 (+) |
| tracheal cuticle | n/a | n/a | 8/8 (++) to (+++) |
| midgut | 1/1 (++++) | 1/1 (++++) | 6/6 (+++) to (++++) |
| ovarian follicles | 0/1 | 0/1 | 0/8 |
| follicular epithelium | 1/1 (+++) | 0/1 | 6/8 (++) to (+++) |
| fat body | 1/1 (++++) | 0/1 | 8/8 (+++) |
| cuticle | 1/1 (+++) | 0/1 | 7/8 (++) to (++++) |
| BTV-10 and BTV-17 Infection Treatment Group | ||||
|---|---|---|---|---|
| Detection of BTV-10 | Detection of BTV-17 | Detection of BTV-10 and BTV-17 | ||
| Number of Midges with BTV Detection | 1 Out of 14 Midges | 3 Out of 14 Midges | 3 Out of 14 Midges | |
| Dissemination Pattern | 1 Midge with BTV-10 Dissemination | 3 Midges with BTV-17 Dissemination | 2 Midges with BTV-10 and 17 Dissemination | 1 Midge with BTV-17 Dissemination and BTV-10 Localized to Midgut |
| Tissue/Organ | ||||
| Johnston’s organ | n/a | 1/1 (++) | n/a | n/a |
| ommatidia | n/a | 2/2 (++) to (++++) | 2/2 (BTV-10 and 17) (++) to (+++) | n/a |
| cerebral ganglion | n/a | 2/2 (++) to (+++) | 2/2 (BTV-10 and 17) (++) to (+++) | n/a |
| salivary gland | n/a | 2/2 (+) | 2/2 (BTV-10 and 17) (++) | n/a |
| dorsal longitudinal muscle | 1/1 (+) | 3/3 (+) | 2/2 (BTV-10 and 17) (+) | 1/1 (BTV-17 only) (+) |
| tracheal cuticle | n/a | 3/3 (++) to (+++) | 2/2 (BTV-10 and 17) (++) | 1/1 (BTV-17 only) (++) |
| midgut | 1/1 (+++) | 2/2 (+++) to (++++) | 1/2 (BTV-10 and 17) (+++) to (++++) | 1/1 (BTV-10 and 17) (++++) |
| ovarian follicles | n/a | 0/3 | 0/2 | 0/1 |
| follicular epithelium | n/a | 3/3 (++) to (+++) | 2/2 (BTV-10 and 17) (++) | 1/1 (BTV-17 only) (+) |
| fat body | 1/1 (+++) | 3/3 (++) to (++++) | 2/2 (BTV-10 and 17) (+++) | 1/1 (BTV-17 only) (+++) |
| cuticle | 1/1 (+++) | 3/3 (++) to (++++) | 2/2 (BTV-10 and 17) (++) to (+++) | 1/1 (BTV-17 only) (++) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carpenter, M.; Benavides Obon, A.; Kopanke, J.; Lee, J.; Reed, K.; Sherman, T.; Rodgers, C.; Stenglein, M.; McDermott, E.; Mayo, C. In Situ Hybridization (RNAscope) Detection of Bluetongue Virus Serotypes 10 and 17 in Experimentally Co-Infected Culicoides sonorensis. Pathogens 2023, 12, 1207. https://doi.org/10.3390/pathogens12101207
Carpenter M, Benavides Obon A, Kopanke J, Lee J, Reed K, Sherman T, Rodgers C, Stenglein M, McDermott E, Mayo C. In Situ Hybridization (RNAscope) Detection of Bluetongue Virus Serotypes 10 and 17 in Experimentally Co-Infected Culicoides sonorensis. Pathogens. 2023; 12(10):1207. https://doi.org/10.3390/pathogens12101207
Chicago/Turabian StyleCarpenter, Molly, AnaMario Benavides Obon, Jennifer Kopanke, Justin Lee, Kirsten Reed, Tyler Sherman, Case Rodgers, Mark Stenglein, Emily McDermott, and Christie Mayo. 2023. "In Situ Hybridization (RNAscope) Detection of Bluetongue Virus Serotypes 10 and 17 in Experimentally Co-Infected Culicoides sonorensis" Pathogens 12, no. 10: 1207. https://doi.org/10.3390/pathogens12101207