

Mathematical Modelling of the Spatial Distribution of a COVID-19 Outbreak with Vaccination Using Diffusion Equation
 , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
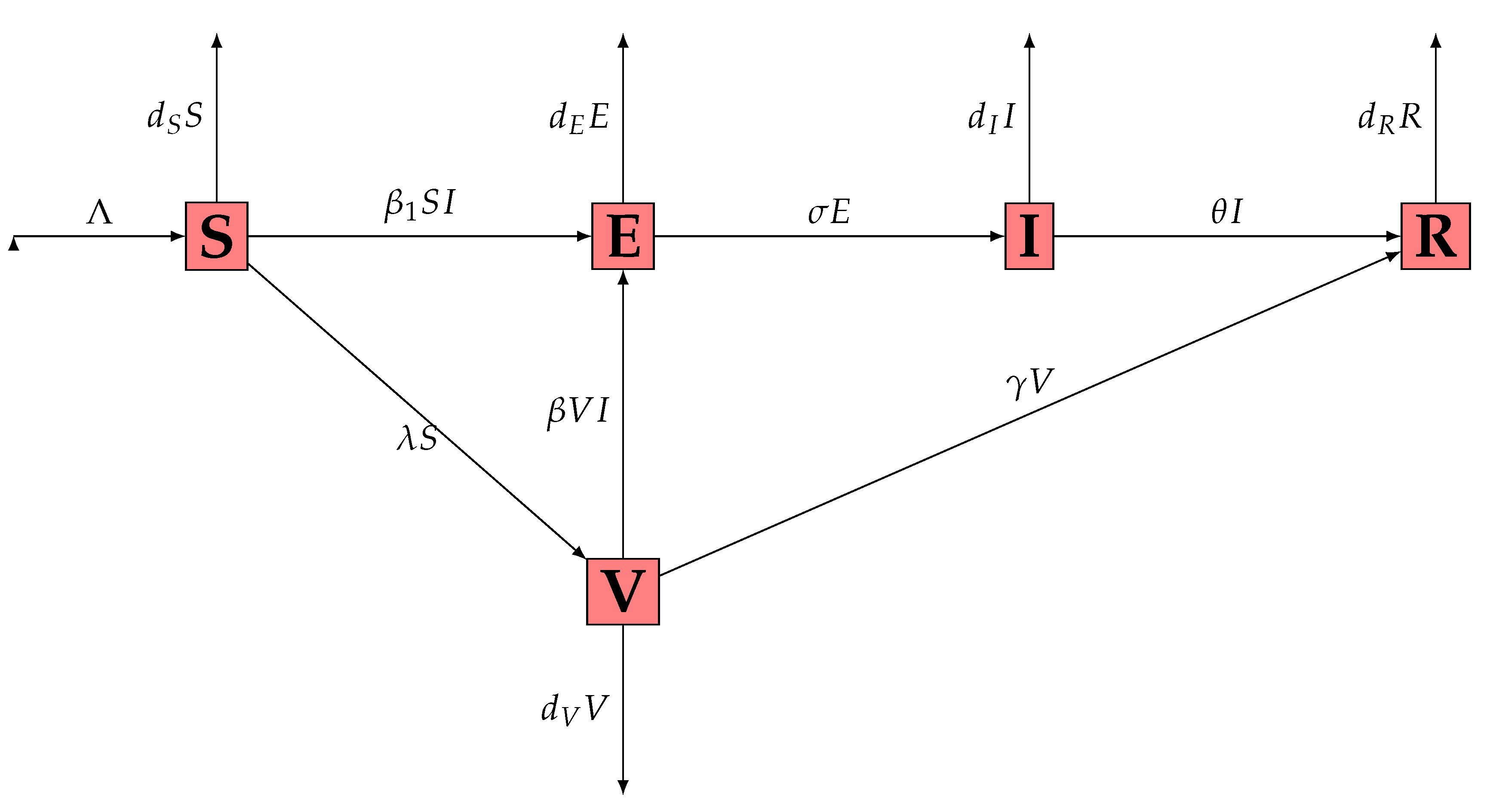
2. Mathematical Model
3. Dynamical Behaviour of the System and Qualitative Analysis
3.1. Existence, Uniqueness, and Positivity
3.2. Equilibria and Basic Reproduction Number
3.2.1. Disease-Free Equilibrium and Basic Reproduction Number
3.2.2. Existence and Uniqueness of the Endemic Equilibrium
3.3. Stability of Equilibrium
3.4. Global Stability of the Disease-Free and Endemic Equilibria by Means of a Lyapunov Function
4. Numerical Simulation and Sensitivity Analysis of
4.1. Numerical Simulation
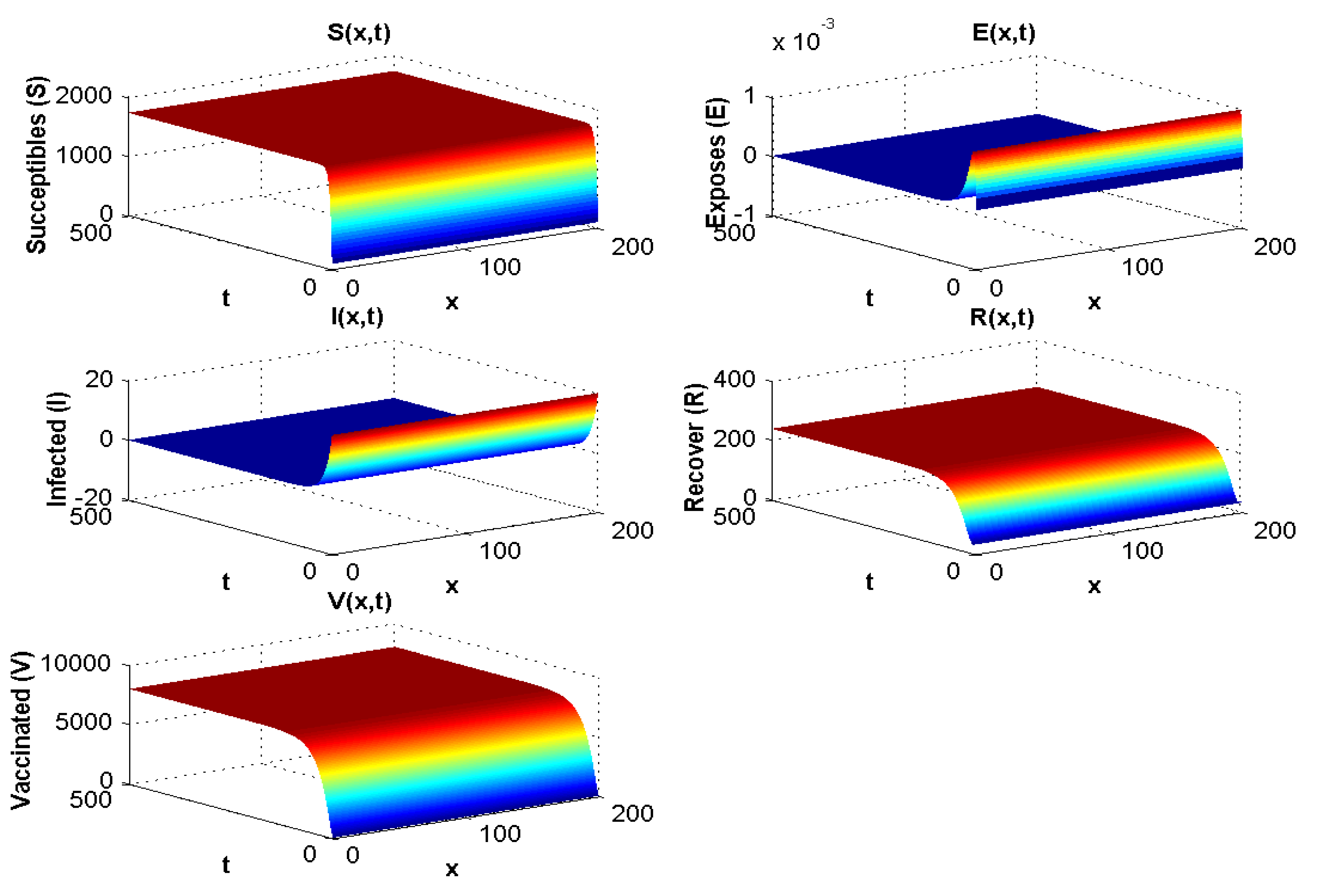
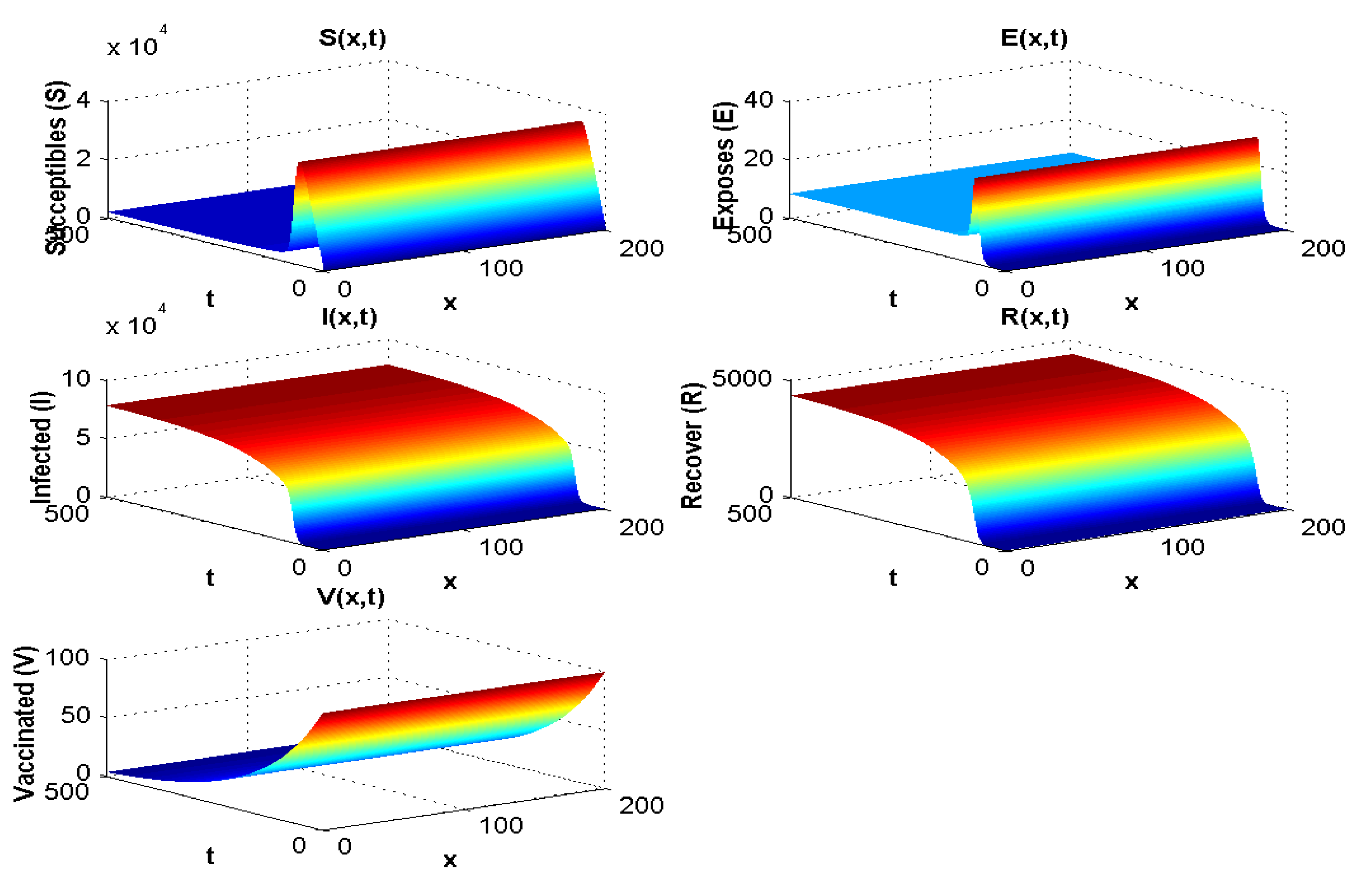
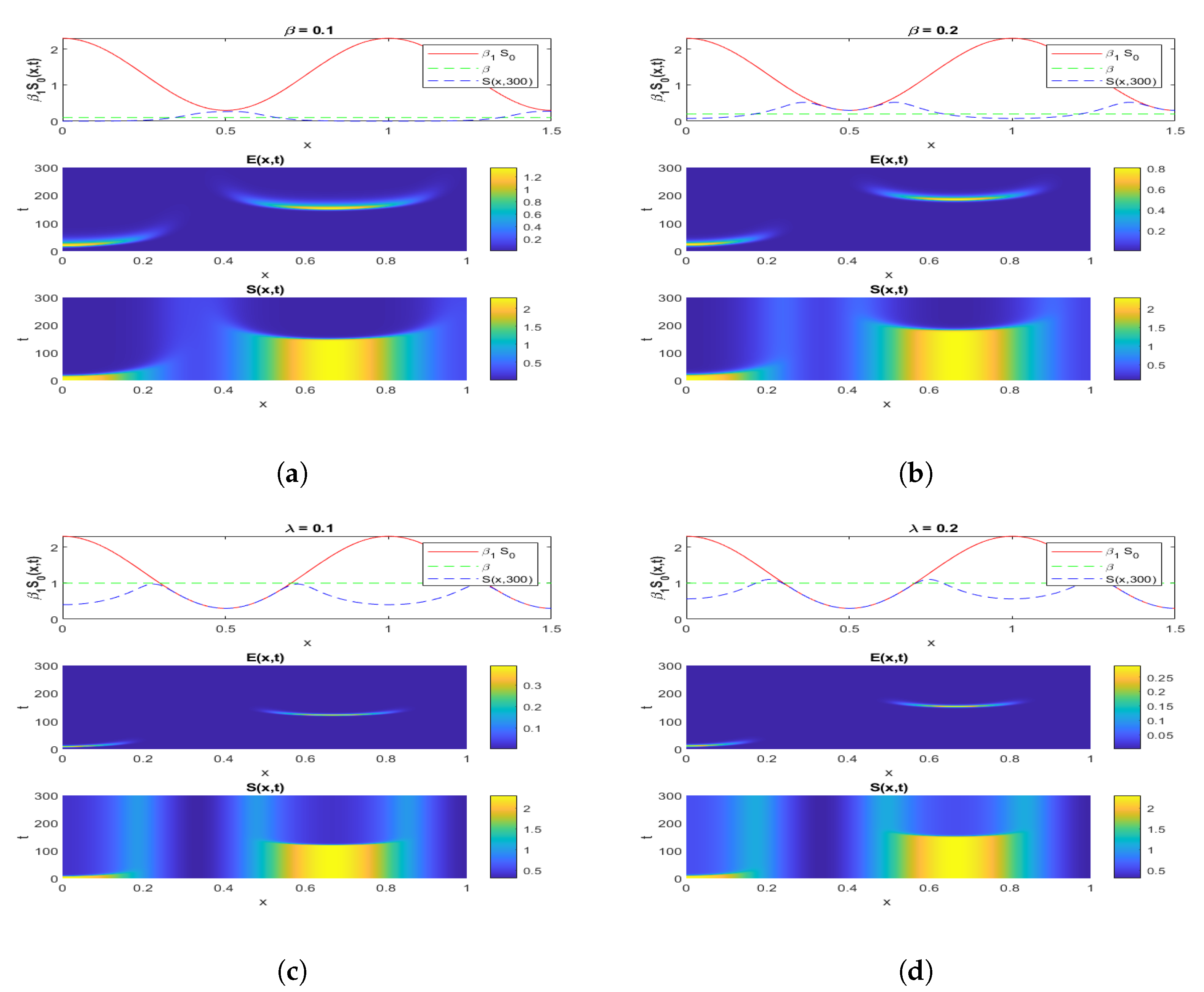
4.1.1. Experiment 1: Numerical Simulation When
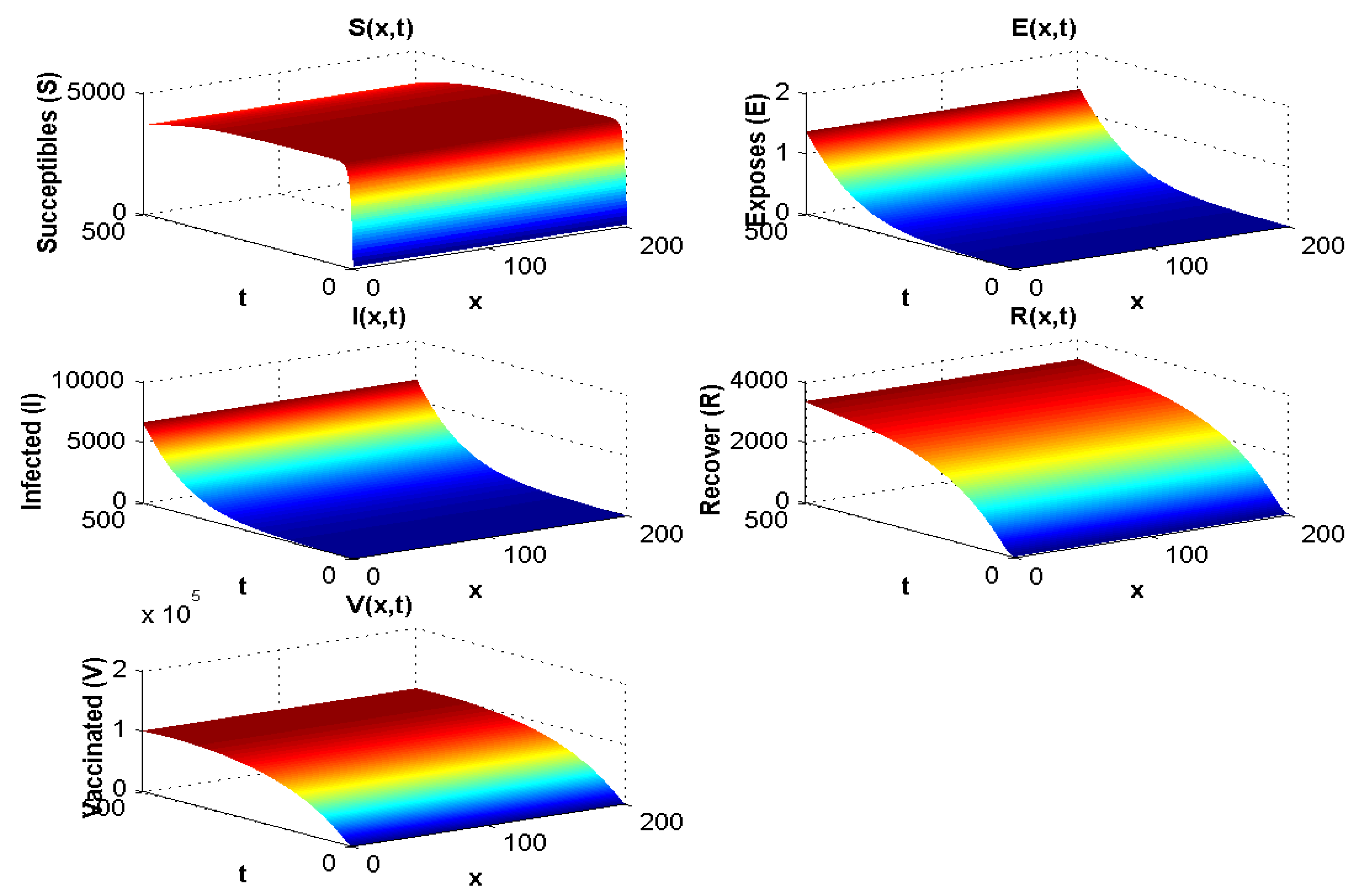
4.1.2. Experiment 2: Numerical Simulation When
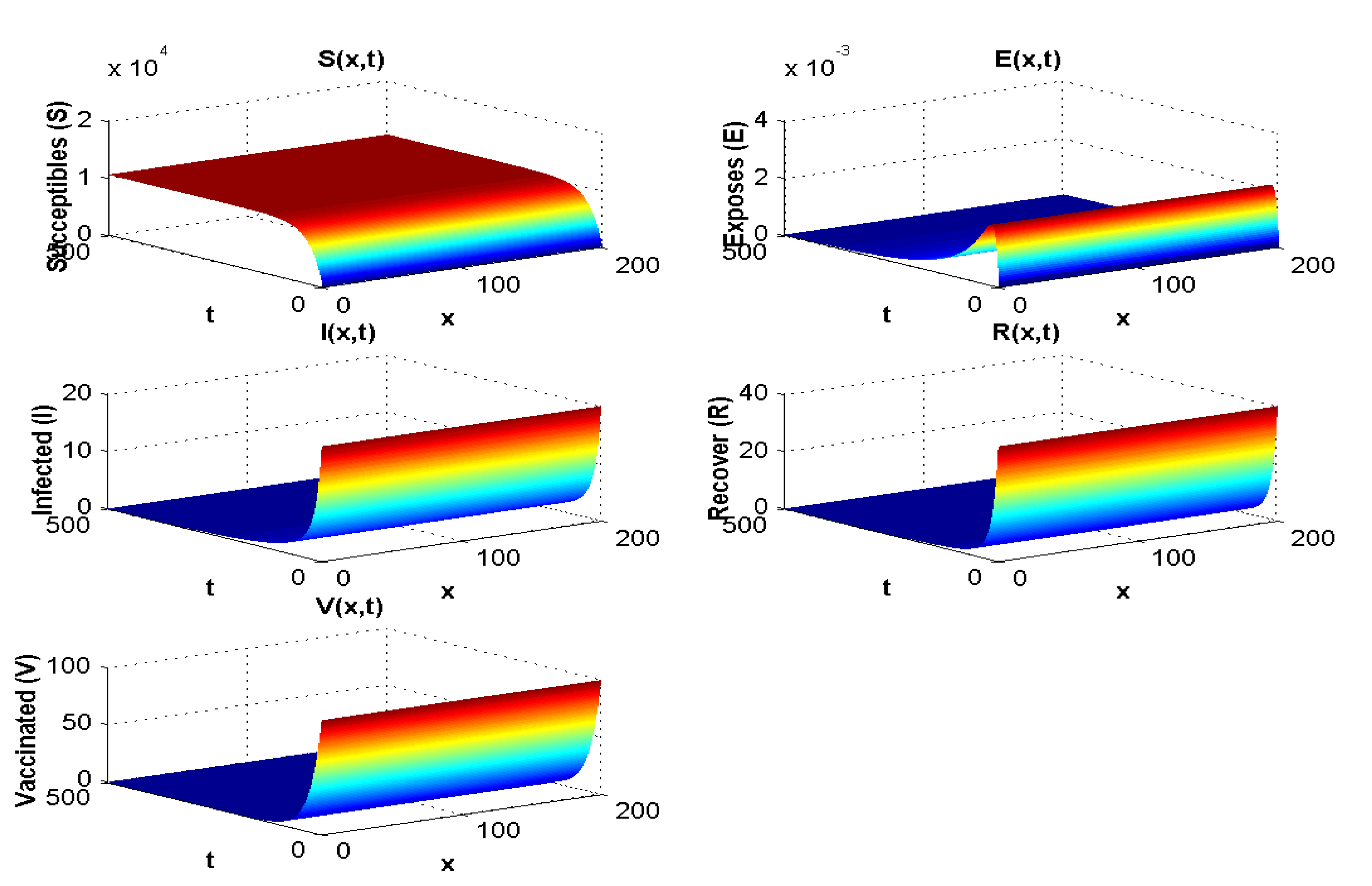
4.1.3. Experiment 3: Numerical Simulation When and
4.1.4. Experiment 4: Numerical Simulation When and
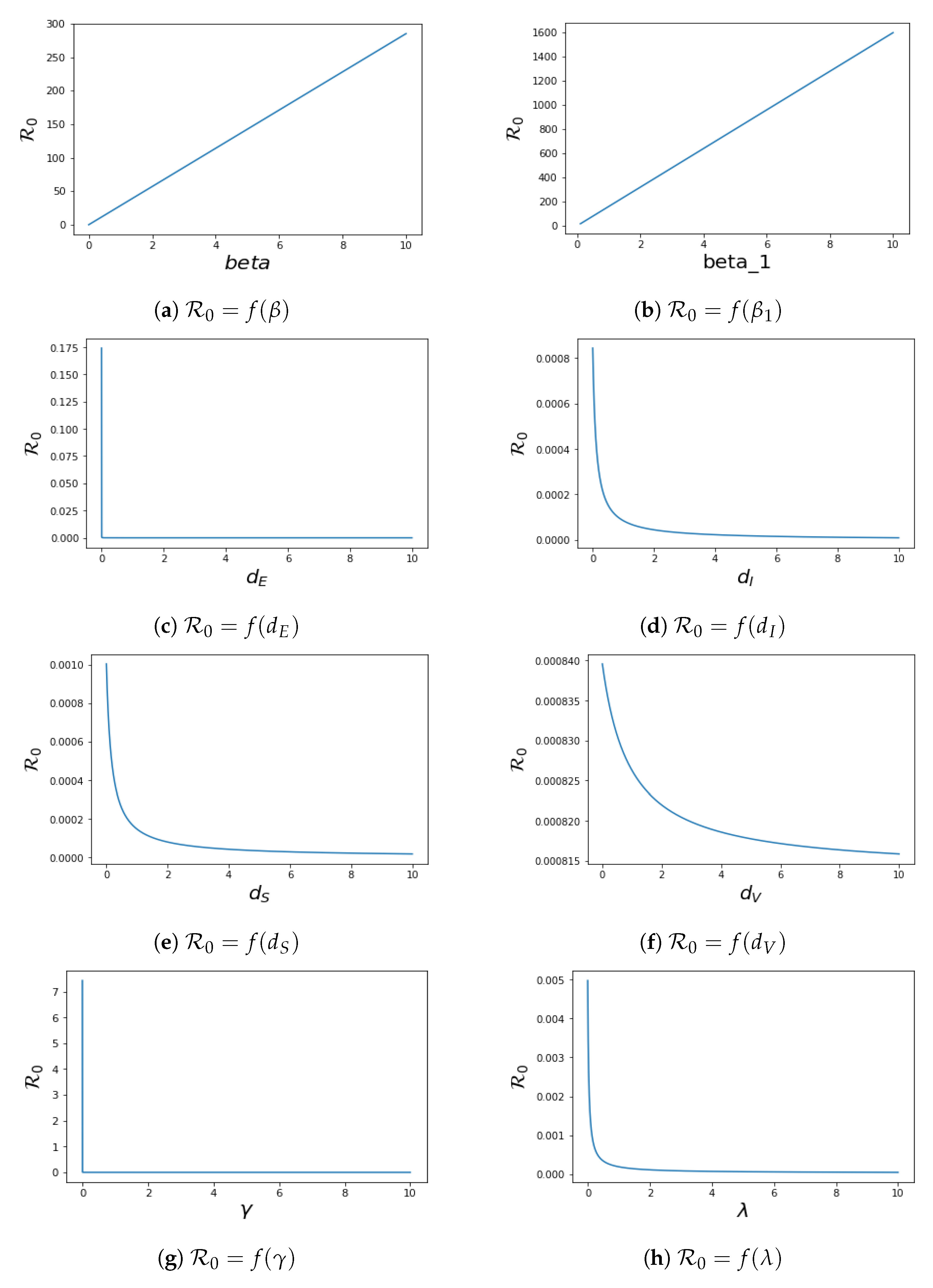
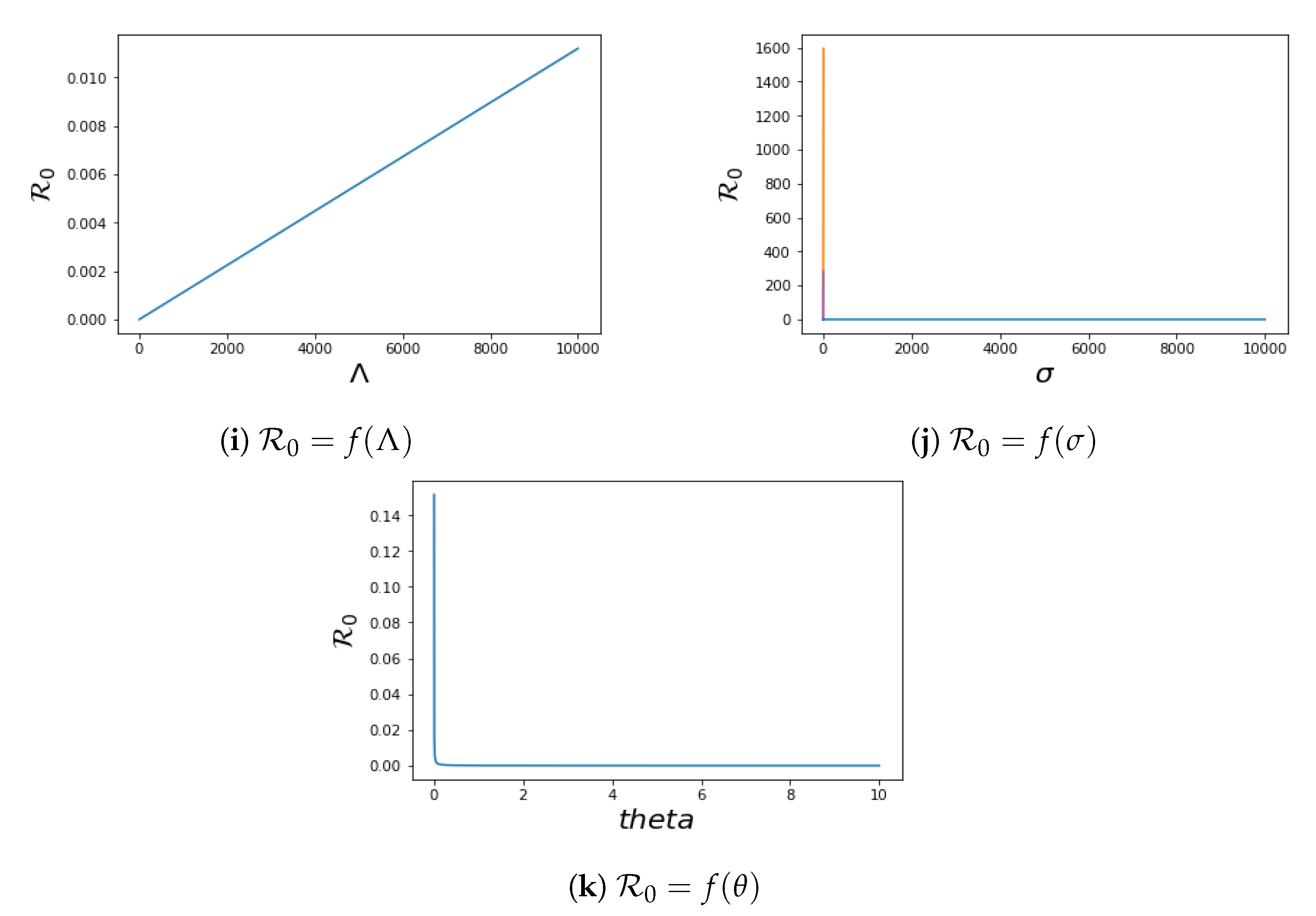
4.2. Local Sensitivity of
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haugh, J.M. Analysis of reaction-diffusion systems with anomalous subdiffusion. Biophys. J. 2009, 97, 435–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abioye, A.I.; Umoh, M.D.; Peter, O.J.; Edogbanya, H.O.; Oguntolu, F.A.; Oshinubi, K.; Amadiegwu, S. Forecasting of COVID-19 pandemic in Nigeria using real statistical data. Commun. Math. Biol. Neurosci. 2021, 2021, 2. [Google Scholar]
- Coccia, M. Factors determining the diffusion of COVID-19 and suggested strategy to prevent future accelerated viral infectivity similar to COVID. Sci. Total. Environ. 2020, 729, 138474. [Google Scholar] [CrossRef] [PubMed]
- Oshinubi, K.; Buhamra, S.S.; Al-Kandari, N.M.; Waku, J.; Rachdi, M.; Demongeot, J. Age Dependent Epidemic Modelling of COVID-19 Outbreak in Kuwait, France and Cameroon. Healthcare 2022, 10, 482. [Google Scholar] [CrossRef]
- Babasola, O.; Oshinubi, K.; Peter, O.J.; Onwuegbuche, F.C.; Oguntolu, F.A. Time-Delayed Modeling of the COVID-19 Dynamics with a Convex Incidence Rate. Inform. Med. Unlock 2022, 35, 101124. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, T.; Meng, X. Stability analysis of a chemostat model with maintenance energy. Appl. Math. Lett. 2017, 68, 1–7. [Google Scholar] [CrossRef]
- Urabe, C.T.; Tanaka, G.; Aihara, K.; Mimura, M. Parameter Scaling for Epidemic Size in a Spatial Epidemic Model with Mobile Individuals. PLoS ONE 2017, 11, e0168127. [Google Scholar] [CrossRef] [Green Version]
- Gaudart, J.; Touré, O.; Dessay, N.; Dicko, A.L.; Ranque, S.; Forest, L.; Demongeot, J.; Doumbo, O.K. Modelling malaria incidence with environmental dependency in a locality of Sudanese savannah area, Mali. Malaria J. 2009, 8, 61. [Google Scholar] [CrossRef] [Green Version]
- Gaudart, J.; Ghassani, M.; Mintsa, J.; Waku, J.; Rachdi, M.; Doumbo, O.K.; Demongeot, J. Demographic and spatial factors as causes of an epidemic spread, the copule approach. Application to the retro-prediction of the Black Death epidemy of 1346. In Proceedings of the 2010 IEEE 24th International Conference on Advanced Information Networking and Applications Workshops, Perth, WA, Australia, 20–23 April 2010; pp. 751–758. [Google Scholar]
- Gaudart, J.; Ghassani, M.; Mintsa, J.; Rachdi, M.; Waku, J.; Demongeot, J. Demography and Diffusion in epidemics: Malaria and Black Death spread. Acta Biotheor. 2010, 58, 277–305. [Google Scholar] [CrossRef]
- Demongeot, J.; Gaudart, J.; Lontos, A.; Promayon, E.; Mintsa, J.; Rachdi, M. Least diffusion zones in morphogenesis and epidemiology. Int. Bifurc. Chaos 2012, 22, 1250028. [Google Scholar] [CrossRef] [Green Version]
- Demongeot, J.; Gaudart, J.; Mintsa, J.; Rachdi, M. Demography in epidemics modelling. Commun. Pure Appl. Anal. 2012, 11, 61–82. [Google Scholar] [CrossRef]
- Redlinger, R. Existence theorem for semilinear parabolic systems with functionals. Nonlinear Anal. 1984, 8, 667682. [Google Scholar] [CrossRef]
- Cooper, I.; Mondal, A.; Antonopoulos, C.G. A SIR model assumption for the spread of COVID-19 in different communities. Chaos, Solitons Fractals 2020, 139, 110057. [Google Scholar] [CrossRef] [PubMed]
- Keeling, M.J.; Woolhouse, M.E.; Shaw, D.J.; Matthews, L.; Chase-Topping, M.; Haydon, D.T.; Cornell, S.J.; Kappey, J.; Wilesmith, J.; Grenfell, B.T. Dynamics of the 2001 UK foot and mouth epidemic stochastic dispers alina heterogeneous landscape. Science 2001, 294, 813–817. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.L.; Lucey, B.; Waller, L.A.; Childs, J.E.; Real, L.A. Predicting the spatial dynamics of rabies epidemics on heterogeneous landscapes. Nature 2002, 99, 3668–3672. [Google Scholar] [CrossRef] [Green Version]
- Wang, T. Dynamics of an epidemic model with spatial diffusion. Phys. Stat. Mech. Its Appl. 2014, 409, 119–129. [Google Scholar] [CrossRef]
- Rajapaksha, R.N.U.; Wijesinghe, M.S.D.; Jayasooriya, K.S.P.; Weerasinghe, W.P.C.; Gunawardana, B.I. An Extended Susceptible-Exposed-Infected-Recovered (SEIR) Model with Vaccination for Predicting the COVID-19 Pandemic in Sri Lanka. medRxiv 2021. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, R.; Kuniya, T. A reaction–diffusion Susceptible–Vaccinated–Infected–Recovered model in a spatially heterogeneous environment with Dirichlet boundary condition. Math. Comput. Simul. 2021, 190, 848–865. [Google Scholar] [CrossRef]
- Li, R.; Song, Y.; Wang, H.; Jiang, G.; Xiao, M. Reactive-diffusion epidemic model on human mobility networks: Analysis and applications to COVID-19 in China. Phys. A 2022, 609, 128337. [Google Scholar] [CrossRef]
- Mehdaoul, M.; Alaoul, A.L.; Tilloua, M. Optimal control for a multi-group reaction–diffusion SIR model with heterogeneous incidence rates. Int. J. Dynam. Control 2022. [Google Scholar] [CrossRef]
- Tu, Y.; Meng, X.; Gao, S.; Hayat, T.; Hobiny, A. Dynamics and strategies evaluations of a novel reaction-diffusion COVID-19 model with direct and aerosol transmission. J. Frankl. Inst. 2022, 359, 10058–10097. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhao, X.Q. Basic reproduction numbers for reaction diffusion epidemic models. SIAM J. Appl. Dyn. Syst. 2012, 11, 1652–1673. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, T. Global stability of the positive equilibrium of a mathematical model for unstirred membrane reactors. Bull. Korean Math. Soc. 2017, 54, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Lasalle, J. The Stability of Dynamical Systems; SIAM: Philadelphia, PA, USA, 1976. [Google Scholar]
- Abioye, A.I.; Peter, O.J.; Ogunseye, H.A.; Oguntolu, F.A.; Oshinubi, K.; Ibrahim, A.A.; Khan, I. Mathematical model of COVID-19 in Nigeria with optimal control. Results Phys. 2021, 28, 104598. [Google Scholar] [CrossRef]
- Ogunrinde, R.B.; Nwajeri, U.K.; Fadugba, S.E.; Ogunrinde, R.R.; Oshinubi, K. Dynamic model of COVID-19 and citizens reaction using fractional derivative. Alex. Eng. J. 2021, 60, 2001–2012. [Google Scholar] [CrossRef]
- Vaziry, A.; Kolokolnikov, T.; Kevrekidis, P.G. Modelling of spatial infection spread through heterogeneous population: From lattice to partial differential equation models. R. Soc. Open Sci. 2022, 9, 220064. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Biological Signification of Parameters |
|---|---|
| Number of incoming susceptible per day | |
| Mortality rate of susceptible per day | |
| Contact between S and I | |
| Contact between V and I | |
| Vaccination rate | |
| Mortality rate of exposed population per day | |
| Infection rate | |
| Mortality rate of infected population per day | |
| Mortality rate of recovered population per day | |
| Recovery rate | |
| Mortality rate of vaccinated population per day |
| Parameters | Values | References |
|---|---|---|
| 350 | Estimated | |
| [26] | ||
| [26] | ||
| [26] | ||
| [26] | ||
| [26] | ||
| [26] | ||
| [26] | ||
| [26] | ||
| [26] | ||
| [26] | ||
| Estimated |
| Parameters | Values | References |
|---|---|---|
| 15,000 | Estimated | |
| [26] | ||
| [26] | ||
| Estimated | ||
| [27] | ||
| Estimated | ||
| [26] | ||
| [26] | ||
| [26] | ||
| [26] | ||
| [26] | ||
| [26] |
| Parameter | Sensitivity Index |
|---|---|
| 0.019 | |
| 1 | |
| 0.99 | |
| 0.00187597 | |
| −0.99727 | |
| −0.00085543 | |
| −0.00004068 | |
| −0.00183557 | |
| −0.3579566 | |
| −0.642 | |
| −0.019 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kammegne, B.; Oshinubi, K.; Babasola, O.; Peter, O.J.; Longe, O.B.; Ogunrinde, R.B.; Titiloye, E.O.; Abah, R.T.; Demongeot, J. Mathematical Modelling of the Spatial Distribution of a COVID-19 Outbreak with Vaccination Using Diffusion Equation. Pathogens 2023, 12, 88. https://doi.org/10.3390/pathogens12010088
Kammegne B, Oshinubi K, Babasola O, Peter OJ, Longe OB, Ogunrinde RB, Titiloye EO, Abah RT, Demongeot J. Mathematical Modelling of the Spatial Distribution of a COVID-19 Outbreak with Vaccination Using Diffusion Equation. Pathogens. 2023; 12(1):88. https://doi.org/10.3390/pathogens12010088
Chicago/Turabian StyleKammegne, Brice, Kayode Oshinubi, Oluwatosin Babasola, Olumuyiwa James Peter, Olumide Babatope Longe, Roseline Bosede Ogunrinde, Emmanuel Olurotimi Titiloye, Roseline Toyin Abah, and Jacques Demongeot. 2023. "Mathematical Modelling of the Spatial Distribution of a COVID-19 Outbreak with Vaccination Using Diffusion Equation" Pathogens 12, no. 1: 88. https://doi.org/10.3390/pathogens12010088