

Assessing Red Fox (Vulpes vulpes) Demographics to Monitor Wildlife Diseases: A Spotlight on Echinococcus multilocularis

 , , , ,
, , , ,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
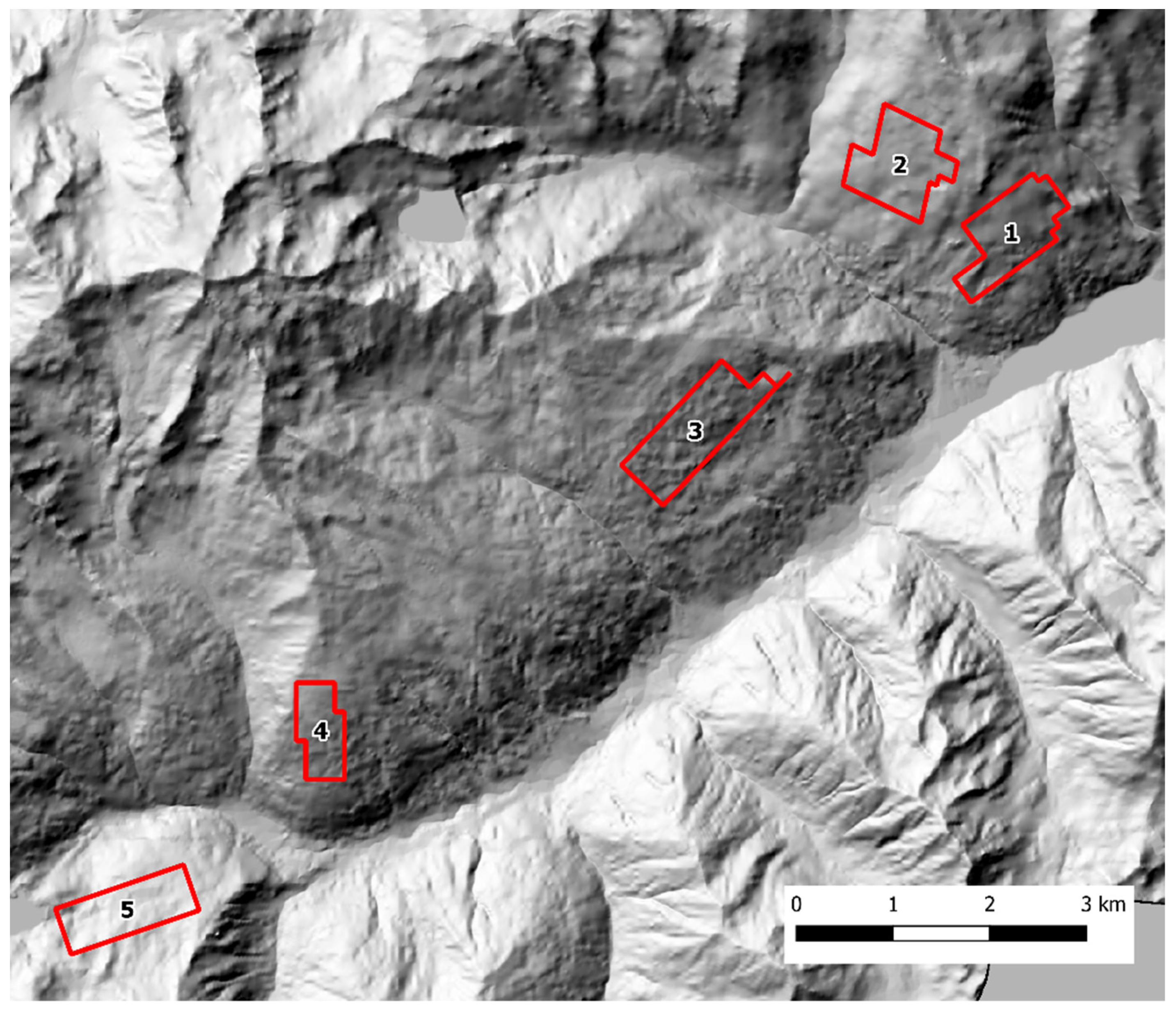
2. Materials and Methods
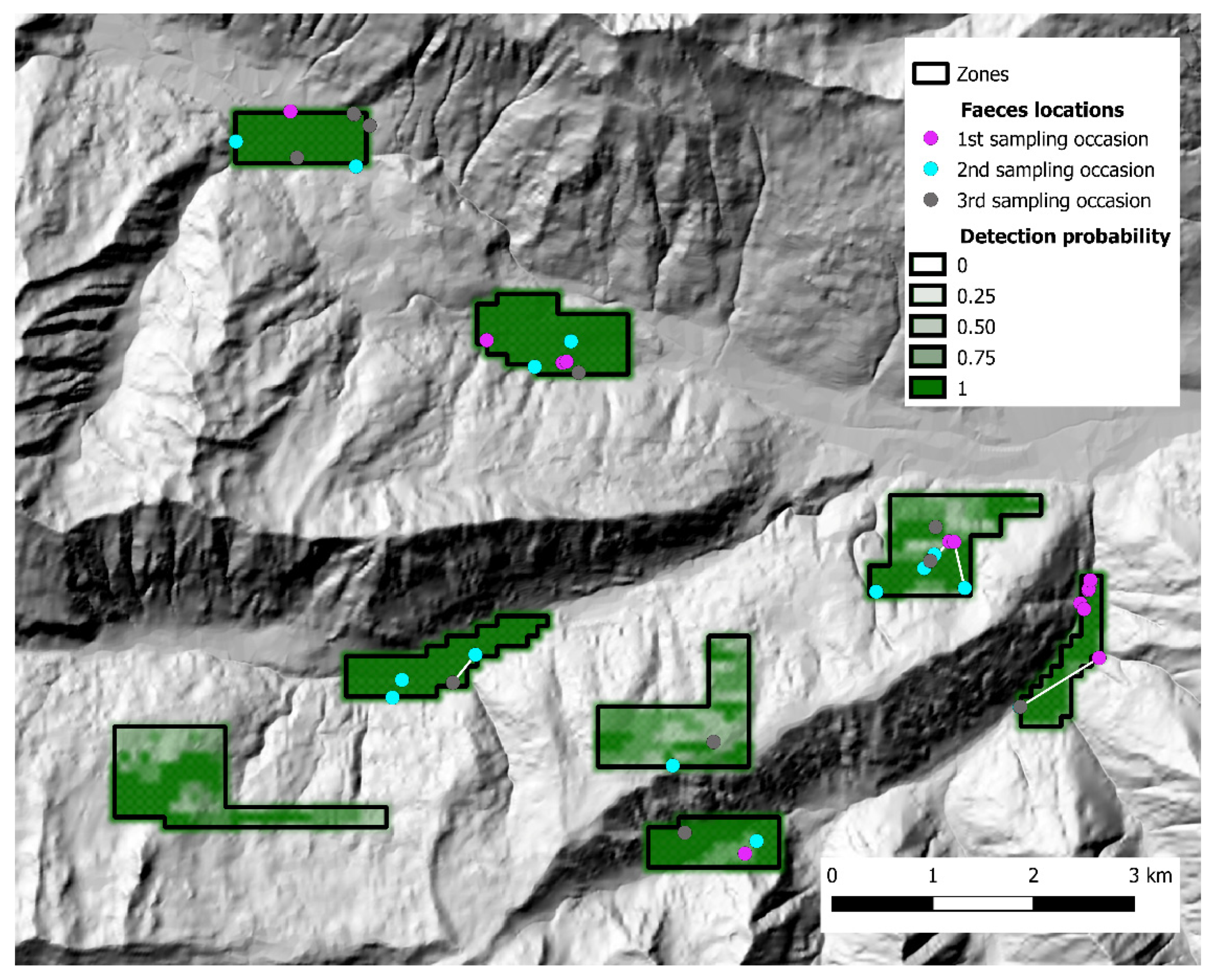
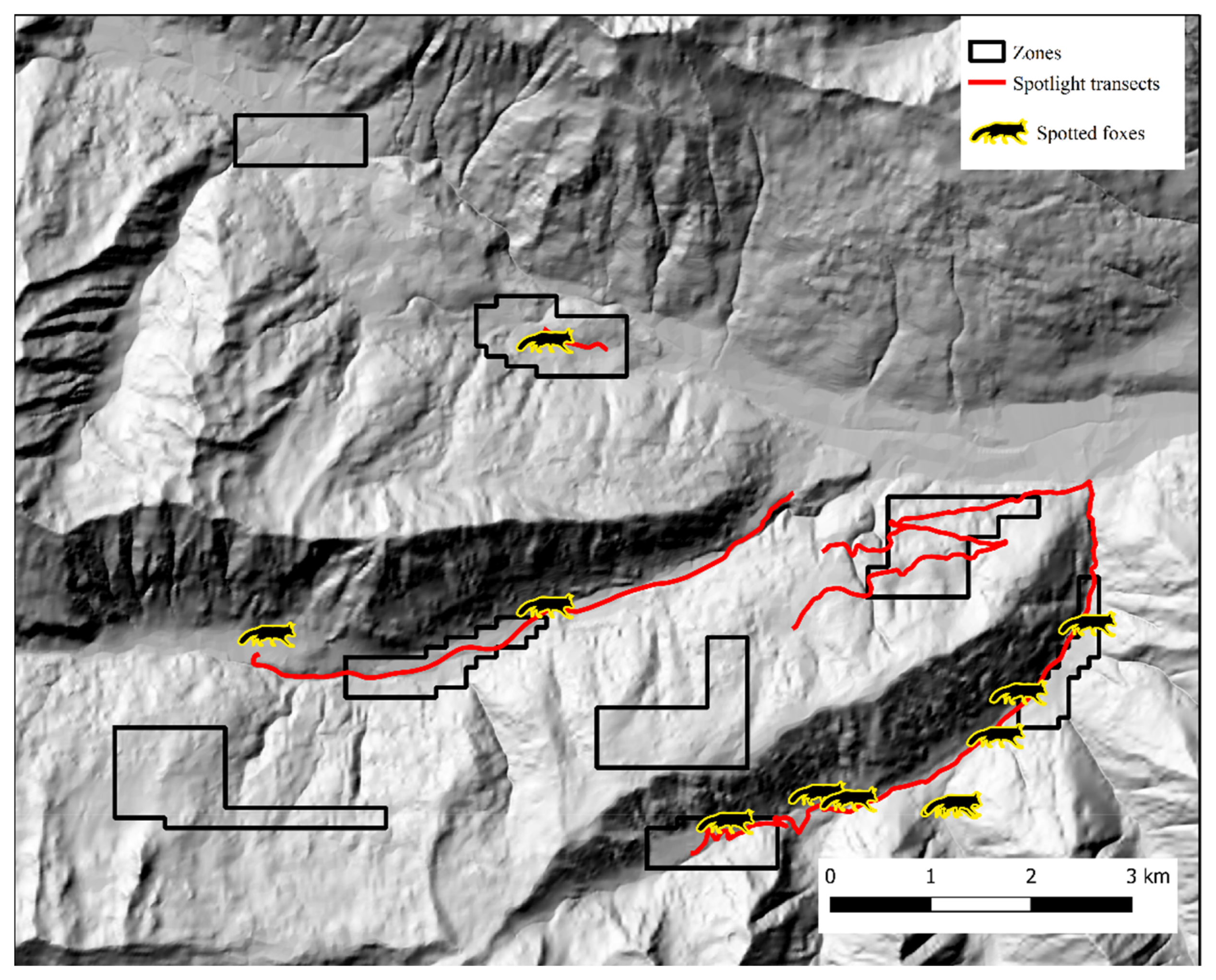
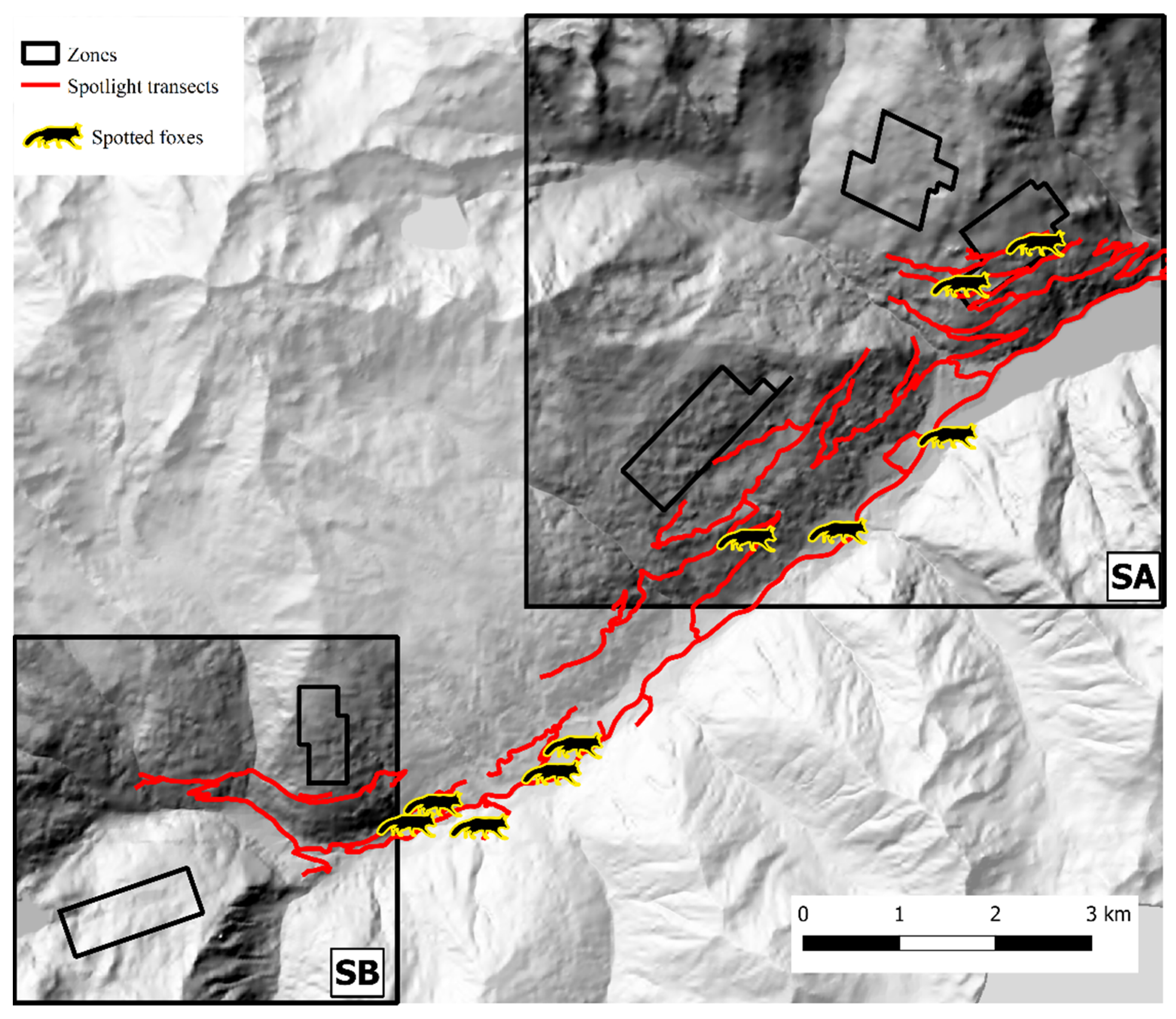
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paolucci, P.; Bon, M. Mammiferi Terrestri d’Italia: Riconoscimento, Ecologia e Tricologia; WBA Books: Verona, Italy, 2022; ISBN 978-88-943738-4-4. [Google Scholar]
- Wandeler, A. Epidemiology and Ecology of Fox Rabies in Europe. In Historical Perspective of Rabies in Europe and the Mediterranean Basin: A Testament to Rabies; King, A.A., Ed.; World Organisation for Animal Health: Paris, France, 2004; pp. 201–214. [Google Scholar]
- Pozio, E. Trichinella and Trichinellosis in Europe. Vet. Glas. 2019, 73, 65–84. [Google Scholar] [CrossRef] [Green Version]
- Woolsey, I.D.; Miller, A.L. Echinococcus Granulosus Sensu Lato and Echinococcus Multilocularis: A Review. Res. Vet. Sci. 2021, 135, 517–522. [Google Scholar] [CrossRef]
- Beltrán, F.; Delibes, M.; Rau, J.R. Methods of Censusing Red Fox (Vulpes vulpes) Populations. Hystrix 1991, 3, 199–214. [Google Scholar] [CrossRef]
- Saunders, G.; Coman, B.; Kinnear, J.; Braysher, M. Managing Vertebrate Pests: Foxes; Australian Government Publ. Service: Sydney, Australia, 1996. [Google Scholar]
- Sadlier, L.M.J.; Webbon, C.C.; Baker, P.J.; Harris, S. Methods of Monitoring Red Foxes Vulpes vulpes and Badgers Meles Meles: Are Field Signs the Answer? Mammal Rev. 2004, 34, 75–98. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, J.; Chandler, R.; Descalzo, J.; Mateo, R.; Ferreras, P. Corrigendum to: Generalized Spatial Mark–Resight Models with Incomplete Identification: An Application to Red Fox Density Estimates. Ecol. Evol. 2019, 9, 4739–4748. [Google Scholar] [CrossRef] [PubMed]
- Wegge, P.; Bakke, B.B.; Odden, M.; Rolstad, J. DNA from Scats Combined with Capture–Recapture Modeling: A Promising Tool for Estimating the Density of Red Foxes—A Pilot Study in a Boreal Forest in Southeast Norway. Mammal Res. 2019, 64, 147–154. [Google Scholar] [CrossRef]
- Lombardi, J.V.; Comer, C.E.; Scognamillo, D.G.; Conway, W.C. Coyote, Fox, and Bobcat Response to Anthropogenic and Natural Landscape Features in a Small Urban Area. Urban Ecosyst. 2017, 20, 1239–1248. [Google Scholar] [CrossRef]
- Ruette, S.; Stahl, P.; Albaret, M. Applying Distance-Sampling Methods to Spotlight Counts of Red Foxes. J. Appl. Ecol. 2003, 40, 32–43. [Google Scholar] [CrossRef]
- Pandolfil, M.; Santolini, R.; Bonacoscia, M. Spotlight Census of the Red Fox (Vulpes vulpes) and the Domestic Cat (Felis catus) in Three Sample Areas of the Marches Region (Central Italy). Hystrix 1991, 3, 221–224. [Google Scholar] [CrossRef]
- Scott, D.M.; Baker, R.; Charman, N.; Karlsson, H.; Yarnell, R.W.; Mill, A.C.; Smith, G.C.; Tolhurst, B.A. A Citizen Science Based Survey Method for Estimating the Density of Urban Carnivores. PLoS ONE 2018, 13, e0197445. [Google Scholar] [CrossRef]
- König, A.; Romig, T. Fox Tapeworm Echinococcus Multilocularis, an Underestimated Threat: A Model for Estimating Risk of Contact. Wildl. Biol. 2010, 16, 258–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soulsbury, C.D.; Iossa, G.; Baker, P.J.; Cole, N.C.; Funk, S.M.; Harris, S. The Impact of Sarcoptic Mange Sarcoptes Scabiei on the British Fox Vulpes vulpes Population. Mammal Rev. 2007, 37, 278–296. [Google Scholar] [CrossRef] [Green Version]
- Harris, S.; Rayner, J.M.V. Urban Fox (Vulpes vulpes) Population Estimates and Habitat Requirements in Several British Cities. J. Anim. Ecol. 1986, 55, 575. [Google Scholar] [CrossRef]
- Harris, S. Distribution, Habitat Utilization and Age Structure of a Suburban Fox (Vulpes vulpes) Population. Mammal Rev. 1977, 7, 25–38. [Google Scholar] [CrossRef]
- Obber, F.; Capello, K.; Mulatti, P.; Lorenzetto, M.; Vendrami, S.; Citterio, C.V. Exploring the Use of Red Fox (Vulpes vulpes) Counts during Deer Censuses as a Tool to Evaluate the Fox Population Trend in the Framework of Disease Surveillance. Hystrix Ital. J. Mammal. 2018, 29, 53–59. [Google Scholar] [CrossRef]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L. Distance Sampling; Springer: Dordrecht, The Netherlands, 1993; ISBN 978-0-412-42670-4. [Google Scholar]
- Shaikenov, B.S. Distribution and Ecology of Echinococcus Multilocularis in Central Asia. Parasitol. Int. 2006, 55, S213–S219. [Google Scholar] [CrossRef]
- Citterio, C.V.; Obber, F.; Trevisiol, K.; Dellamaria, D.; Celva, R.; Bregoli, M.; Ormelli, S.; Sgubin, S.; Bonato, P.; Da Rold, G.; et al. Echinococcus Multilocularis and Other Cestodes in Red Foxes (Vulpes vulpes) of Northeast Italy, 2012–2018. Parasites Vectors 2021, 14, 29. [Google Scholar] [CrossRef]
- Panel on Animal Health and Welfare. Echinococcus Multilocularis Infection in Animals. EFSA J. 2015, 13, 4373. [Google Scholar] [CrossRef] [Green Version]
- Boué, F.; Boes, J.; Boireau, P.; Claes, M.; Cook, A.J.C.; Dorny, P.; Enemark, H.; van der Giessen, J.; Hunt, K.R.; Howell, M.; et al. Development of Harmonised Schemes for the Monitoring and Reporting of Echinococcus in Animals and Foodstuffs in the European Union. EFSA Support. Publ. 2017, 7, 36E. [Google Scholar] [CrossRef]
- Eckert, J.; Deplazes, P. Biological, Epidemiological, and Clinical Aspects of Echinococcosis, a Zoonosis of Increasing Concern. Clin. Microbiol. Rev. 2004, 17, 107. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control Echinococcosis (ECDC). Annual Epidemiological Report for 2020; European Centre for Disease Prevention and Control Echinococcosis: Stockholm, Sweden, 2022. [Google Scholar]
- Hegglin, D.; Deplazes, P. Control of Echinococcus Multilocularis: Strategies, Feasibility and Cost-Benefit Analyses. Int. J. Parasitol. 2013, 43, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Hegglin, D.; Bontadina, F.; Gloor, S.; Romig, T.; Deplazes, P.; Kern, P. Survey of Public Knowledge about Echinococcus Multilocularis in Four European Countries: Need for Proactive Information. BMC Public Health 2008, 8, 247. [Google Scholar] [CrossRef] [Green Version]
- Manfredi, M.T.; Genchi, C.; Deplazes, P.; Trevisiol, K.; Fraquelli, C. Echinococcus Multilocularis Infection in Red Foxes in Italy. Vet. Rec. 2002, 150, 757. [Google Scholar] [CrossRef]
- Manfredi, M.; Di Cerbo, A.R.; Trevisiol, K. An Updating on the Epidemiological Situation of Echinococcus Multilocularis in Trentino Alto Adige (Northern Italy). Parassitologia 2004, 46, 431–433. [Google Scholar] [PubMed]
- Di Cerbo, A.R.; Manfredi, M.T.; Trevisiol, K.; Bregoli, M.; Ferrari, N.; Pirinesi, F.; Bazzoli, S. Intestinal Helminth Communities of the Red Fox (Vulpes vulpes L.) in the Italian Alps. Acta Parasitol. 2008, 53, 302–311. [Google Scholar] [CrossRef]
- Casulli, A.; Manfredi, M.T.; La Rosa, G.; Di Cerbo, A.R.; Dinkel, A.; Romig, T.; Deplazes, P.; Genchi, C.; Pozio, E. Echinococcus Multilocularis in Red Foxes (Vulpes vulpes) of the Italian Alpine Region: Is There a Focus of Autochthonous Transmission? Int. J. Parasitol. 2005, 35, 1079–1083. [Google Scholar] [CrossRef]
- Casulli, A.; Bart, J.M.; Knapp, J.; La Rosa, G.; Dusher, G.; Gottstein, B.; Di Cerbo, A.; Manfredi, M.T.; Genchi, C.; Piarroux, R.; et al. Multi-Locus Microsatellite Analysis Supports the Hypothesis of an Autochthonous Focus of Echinococcus Multilocularis in Northern Italy. Int. J. Parasitol. 2009, 39, 837–842. [Google Scholar] [CrossRef]
- Obber, F.; Celva, R.; Da Rold, G.; Trevisiol, K.; Ravagnan, S.; Danesi, P.; Cenni, L.; Rossi, C.; Bonato, P.; Capello, K.; et al. A Highly Endemic Area of Echinococcus Multilocularis Identified through a Comparative Re-Assessment of Prevalence in the Red Fox (Vulpes vulpes), Alto Adige (Italy: 2019–2020). PLoS ONE 2022, 17, e0268045. [Google Scholar] [CrossRef]
- Massolo, A.; Valli, D.; Wassermann, M.; Cavallero, S.; D’Amelio, S.; Meriggi, A.; Torretta, E.; Serafini, M.; Casulli, A.; Zambon, L.; et al. Unexpected Echinococcus Multilocularis Infections in Shepherd Dogs and Wolves in South-Western Italian Alps: A New Endemic Area? Int. J. Parasitol. Parasites Wildl. 2018, 7, 309–316. [Google Scholar] [CrossRef]
- Kéry, M.; Gardner, B.; Stoeckle, T.; Weber, D.; Royle, J.A. Use of Spatial Capture-Recapture Modeling and DNA Data to Estimate Densities of Elusive Animals. Conserv. Biol. 2011, 25, 356–364. [Google Scholar] [CrossRef]
- Spehar, S.N.; Loken, B.; Rayadin, Y.; Royle, J.A. Comparing Spatial Capture-Recapture Modeling and Nest Count Methods to Estimate Orangutan Densities in the Wehea Forest, East Kalimantan, Indonesia. Biol. Conserv. 2015, 191, 185–193. [Google Scholar] [CrossRef]
- Gimenez, O.; Gatti, S.; Duchamp, C.; Germain, E.; Laurent, A.; Zimmermann, F.; Marboutin, E. Spatial Density Estimates of Eurasian Lynx (Lynx Lynx) in the French Jura and Vosges Mountains. Ecol. Evol. 2019, 9, 11707–11715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsø, L.K.; Dupont, P.; Rød-Eriksen, L.; Andersskog, I.P.Ø.; Ulvund, K.R.; Flagstad, Ø.; Bischof, R.; Eide, N.E. Estimating Red Fox Density Using Non-Invasive Genetic Sampling and Spatial Capture–Recapture Modelling. Oecologia 2022, 198, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Tarquini, S.; Isola, I.; Battistini, A. TINITALY, a Digital Elevation Model of Italy with a 10 Meters Cell Size (Version 1.0); Istituto Nazionale di Geofisica e Vulcanologia (INGV): Rome, Italy, 2007. [CrossRef]
- Borchers, D.L.; Efford, M.G. Spatially Explicit Maximum Likelihood Methods for Capture-Recapture Studies. Biometrics 2008, 64, 377–385. [Google Scholar] [CrossRef]
- Royle, J.A.; Kéry, M.; Guélat, J. Spatial Capture-Recapture Models for Search-Encounter Data. Methods Ecol. Evol. 2011, 2, 602–611. [Google Scholar] [CrossRef]
- Molinari, P.; Jobin, A.M.; Breitenmoser, U. Predatori in Azione: Manuale Di Identificazione Delle Predazioni e Di Altri Segni Di Presenza Dei Grandi Mammiferi Carnivori; Wildvet Projects: Stampa, Switzerland, 2000; ISBN 9788890052705. [Google Scholar]
- Lang, Z.; Reiczigel, J. Confidence Limits for Prevalence of Disease Adjusted for Estimated Sensitivity and Specificity. Prev. Vet. Med. 2014, 113, 13–22. [Google Scholar] [CrossRef]
- Zecchin, B.; De Nardi, M.; Nouvellet, P.; Vernesi, C.; Babbucci, M.; Crestanello, B.; Bagó, Z.; Bedeković, T.; Hostnik, P.; Milani, A.; et al. Genetic and Spatial Characterization of the Red Fox (Vulpes vulpes) Population in the Area Stretching between the Eastern and Dinaric Alps and Its Relationship with Rabies and Canine Distemper Dynamics. PLoS ONE 2019, 14, e0213515. [Google Scholar] [CrossRef] [Green Version]
- Taberlet, P.; Griffin, S.; Goossens, B.; Questiau, S.; Manceau, V.; Escaravage, N.; Waits, L.P.; Bouvet, J. Reliable Genotyping of Samples with Very Low DNA Quantities Using PCR. Nucleic Acids Res. 1996, 24, 3189–3194. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research-an Update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. MICRO-CHECKER: Software for Identifying and Correcting Genotyping Errors in Microsatellite Data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Lunn, D.; Spiegelhalter, D.; Thomas, A.; Best, N. The BUGS Project: Evolution, Critique and Future Directions. Stat. Med. 2009, 28, 3049–3067. [Google Scholar] [CrossRef] [PubMed]
- Gilks, W.R.; Thomas, A.; Spiegelhalter, D.J. A Language and Program for Complex Bayesian Modelling. J. R. Stat. Soc. Ser. D (Stat.) 1994, 43, 169. [Google Scholar] [CrossRef] [Green Version]
- Gompertz, B. On the Nature of the Function Expressive of the Law of Human Mortality, and on a New Mode of Determining the Value of Life Contingencies. Philos. Trans. R. Soc. Lond. 1825, 115, 513–583. [Google Scholar]
- Tanner, M.A.; Wong, W.H. The Calculation of Posterior Distributions by Data Augmentation. J. Am. Stat. Assoc. 1987, 82, 528–540. [Google Scholar] [CrossRef]
- Royle, J.A.; Dorazio, R.M.; Link, W.A. Analysis of Multinomial Models with Unknown Index Using Data Augmentation. J. Comput. Graph. Stat. 2007, 16, 67–85. [Google Scholar] [CrossRef]
- Cavallini, P. Variation in the Social System of the Red Fox. Ethol. Ecol. Evol. 1996, 8, 323–342. [Google Scholar] [CrossRef]
- Pandolfi, M.; Forconi, P.; Montecchiari, L. Spatial Behaviour of the Red Fox (Vulpes vulpes) in a Rural Area of Central Italy. Ital. J. Zool. 1997, 64, 351–358. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013; Volume 2. [Google Scholar]
- Sturtz, S.; Ligges, U.; Gelman, A. R2WinBUGS: A Package for Running WinBUGS from R. J. Stat. Softw. 2005, 12, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kery, M. Introduction to WinBUGS for Ecologists; Elsevier Inc.: Amsterdam, The Netherlands, 2010; ISBN 9780123786050. [Google Scholar]
- Arneberg, P. An Ecological Law and Its Macroecological Consequences as Revealed by Studies of Relationships between Host Densities and Parasite Prevalence. Ecography 2001, 24, 352–358. [Google Scholar] [CrossRef]
- Anderson, R.M.; May, R.M. Regulation and Stability of Host-Parasite Population Interactions: I. Regulatory Processes. J. Anim. Ecol. 1978, 47, 219. [Google Scholar] [CrossRef]
- May, R.M.; Anderson, R.M. Regulation and Stability of Host-Parasite Population Interactions: II. Destabilizing Processes. J. Anim. Ecol. 1978, 47, 249. [Google Scholar] [CrossRef]
- Hopkins, S.R.; Fleming-Davies, A.E.; Belden, L.K.; Wojdak, J.M. Systematic Review of Modelling Assumptions and Empirical Evidence: Does Parasite Transmission Increase Nonlinearly with Host Density? Methods Ecol. Evol. 2020, 11, 476–486. [Google Scholar] [CrossRef]
- Dobson, A.P. Models for Multi-Species Parasite-Host Communities. In Parasite Communities: Patterns and Processes; Springer: Dordrecht, The Netherlands, 1990; pp. 261–288. [Google Scholar]
- Lopez, J.E. Parasite Prevalence and the Size of Host Populations: An Experimental Test. J. Parasitol. 2005, 91, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Raoul, F.; Hegglin, D.; Giraudoux, P. Trophic Ecology, Behaviour and Host Population Dynamics in Echinococcus Multilocularis Transmission. Vet. Parasitol. 2015, 213, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Hegglin, D.; Bontadina, F.; Contesse, P.; Gloor, S.; Deplazes, P. Plasticity of Predation Behaviour as a Putative Driving Force for Parasite Life-Cycle Dynamics: The Case of Urban Foxes and Echinococcus Multilocularis Tapeworm. Funct. Ecol. 2007, 21, 552–560. [Google Scholar] [CrossRef]
- Robardet, E.; Giraudoux, P.; Caillot, C.; Boue, F.; Cliquet, F.; Augot, D.; Barrat, J. Infection of Foxes by Echinococcocus Multilocuiaris in Urban and Suburban Areas of Nancy, France: Influence of Feeding Habits and Environment. Parasite 2008, 15, 77–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raoul, F.; Michelat, D.; Ordinaire, M.; Décoté, Y.; Aubert, M.; Delattre, P.; Deplazes, P.; Giraudoux, P. Echinococcus Multilocularis: Secondary Poisoning of Fox Population during a Vole Outbreak Reduces Environmental Contamination in a High Endemicity Area. Int. J. Parasitol. 2003, 33, 945–954. [Google Scholar] [CrossRef]
- Eckert, J.; Gemmell, M.; Meslin, F.; Pawlowski, Z. Who/Oie Manual on Echinococcosis in Humans and Animals: A Public Health Problem of Global Concern: 9789290445227: Medicine & Health Science Books @ Amazon.Com. Available online: http://www.oie.int (accessed on 27 September 2022).
- Porteus, T.A.; Reynolds, J.C.; McAllister, M.K. Population Dynamics of Foxes during Restricted-Area Culling in Britain: Advancing Understanding through State-Space Modelling of Culling Records. PLoS ONE 2019, 14, e0225201. [Google Scholar] [CrossRef] [Green Version]
- Newsome, T.M.; Crowther, M.S.; Dickman, C.R. Rapid Recolonisation by the European Red Fox: How Effective Are Uncoordinated and Isolated Control Programs? Eur. J. Wildl. Res. 2014, 60, 749–757. [Google Scholar] [CrossRef]
- Bino, G.; Dolev, A.; Yosha, D.; Guter, A.; King, R.; Saltz, D.; Kark, S. Abrupt Spatial and Numerical Responses of Overabundant Foxes to a Reduction in Anthropogenic Resources. J. Appl. Ecol. 2010, 47, 1262–1271. [Google Scholar] [CrossRef]
- Jacob, J.; Imholt, C.; Caminero-Saldaña, C.; Couval, G.; Giraudoux, P.; Herrero-Cófreces, S.; Horváth, G.; Luque-Larena, J.J.; Tkadlec, E.; Wymenga, E. Europe-Wide Outbreaks of Common Voles in 2019. J. Pest Sci. 2020, 93, 703–709. [Google Scholar] [CrossRef] [Green Version]
- Gottstein, B.; Stojkovic, M.; Vuitton, D.A.; Millon, L.; Marcinkute, A.; Deplazes, P. Threat of Alveolar Echinococcosis to Public Health—A Challenge for Europe. Trends Parasitol. 2015, 31, 407–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweiger, A.; Ammann, R.W.; Candinas, D.; Clavien, P.A.; Eckert, J.; Gottstein, B.; Halkic, N.; Muellhaupt, B.; Prinz, B.M.; Reichen, J.; et al. Human Alveolar Echinococcosis after Fox Population Increase, Switzerland. Emerg. Infect. Dis. 2007, 13, 878–882. [Google Scholar] [CrossRef] [PubMed]
- Liccioli, S.; Giraudoux, P.; Deplazes, P.; Massolo, A. Wilderness in the “city” Revisited: Different Urbes Shape Transmission of Echinococcus Multilocularis by Altering Predator and Prey Communities. Trends Parasitol. 2015, 31, 297–305. [Google Scholar] [CrossRef]
- Monne, I.; Fusaro, A.; Valastro, V.; Citterio, C.; Pozza, M.D.; Obber, F.; Trevisiol, K.; Cova, M.; De Benedictis, P.; Bregoli, M.; et al. A Distinct CDV Genotype Causing a Major Epidemic in Alpine Wildlife. Vet. Microbiol. 2011, 150, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Capello, K.; Mulatti, P.; Comin, A.; Gagliazzo, L.; Guberti, V.; Citterio, C.; De Benedictis, P.; Lorenzetto, M.; Costanzi, C.; Vio, P.; et al. Impact of Emergency Oral Rabies Vaccination of Foxes in Northeastern Italy, 28 December 2009–20 January 2010: Preliminary Evaluation. Eurosurveillance 2010, 15, 19617. [Google Scholar] [CrossRef]
- Mulatti, P.; Bonfanti, L.; Patregnani, T.; Lorenzetto, M.; Ferrè, N.; Gagliazzo, L.; Casarotto, C.; Ponti, A.M.; Ferri, G.; Marangon, S. 2008–2011 Sylvatic Rabies Epidemic in Italy: Challenges and Experiences. Pathog. Glob. Health 2013, 107, 346–353. [Google Scholar] [CrossRef] [Green Version]
- Nouvellet, P.; Donnelly, C.A.; De Nardi, M.; Rhodes, C.J.; De Benedictis, P.; Citterio, C.; Obber, F.; Lorenzetto, M.; Pozza, M.D.; Cauchemez, S.; et al. Rabies and Canine Distemper Virus Epidemics in the Red Fox Population of Northern Italy (2006–2010). PLoS ONE 2013, 8, e61588. [Google Scholar] [CrossRef]
- Bianco, A.; Zecchin, B.; Fusaro, A.; Schivo, A.; Ormelli, S.; Bregoli, M.; Citterio, C.V.; Obber, F.; Dellamaria, D.; Trevisiol, K.; et al. Two Waves of Canine Distemper Virus Showing Different Spatio-Temporal Dynamics in Alpine Wildlife (2006–2018). Infect. Genet. Evol. 2020, 84, 104359. [Google Scholar] [CrossRef]
- Artois, M.; Langlais, M.; Suppo, C. Simulation of Rabies Control within an Increasing Fox Population. Ecol. Model. 1997, 97, 23–34. [Google Scholar] [CrossRef]
- Breitenmoser, U.; Müller, U.; Kappeler, A.; Zanoni, R.G. The Final Stage of Rabies in Switzerland. Schweiz. Arch. Tierheilkd. 2000, 142, 447–454. [Google Scholar]
- Gloor, S.; Bontadina, F.; Hegglin, D.; Deplazes, P.; Breitenmoser, U. The Rise of Urban Fox Populations in Switzerland. Z. Saugetierkd. 2001, 66, 155–164. [Google Scholar]
- Chautan, M.; Pontier, D.; Artois, M. Role of Rabies in Recent Demographic Changes in Red Fox (Vulpes vulpes) Populations in Europe. Mammalia 2000, 64, 391–410. [Google Scholar] [CrossRef]
- Delcourt, J.; Brochier, B.; Delvaux, D.; Vangeluwe, D.; Poncin, P. Fox Vulpes vulpes Population Trends in Western Europe during and after the Eradication of Rabies. Mammal Rev. 2022, 52, 343–359. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Celva, R.; Crestanello, B.; Obber, F.; Dellamaria, D.; Trevisiol, K.; Bregoli, M.; Cenni, L.; Agreiter, A.; Danesi, P.; Hauffe, H.C.; et al. Assessing Red Fox (Vulpes vulpes) Demographics to Monitor Wildlife Diseases: A Spotlight on Echinococcus multilocularis. Pathogens 2023, 12, 60. https://doi.org/10.3390/pathogens12010060
Celva R, Crestanello B, Obber F, Dellamaria D, Trevisiol K, Bregoli M, Cenni L, Agreiter A, Danesi P, Hauffe HC, et al. Assessing Red Fox (Vulpes vulpes) Demographics to Monitor Wildlife Diseases: A Spotlight on Echinococcus multilocularis. Pathogens. 2023; 12(1):60. https://doi.org/10.3390/pathogens12010060
Chicago/Turabian StyleCelva, Roberto, Barbara Crestanello, Federica Obber, Debora Dellamaria, Karin Trevisiol, Marco Bregoli, Lucia Cenni, Andreas Agreiter, Patrizia Danesi, Heidi Christine Hauffe, and et al. 2023. "Assessing Red Fox (Vulpes vulpes) Demographics to Monitor Wildlife Diseases: A Spotlight on Echinococcus multilocularis" Pathogens 12, no. 1: 60. https://doi.org/10.3390/pathogens12010060