
In Silico Analysis of a Drosophila Parasitoid Venom Peptide Reveals Prevalence of the Cation–Polar–Cation Clip Motif in Knottin Proteins
 , , , and
, , , and 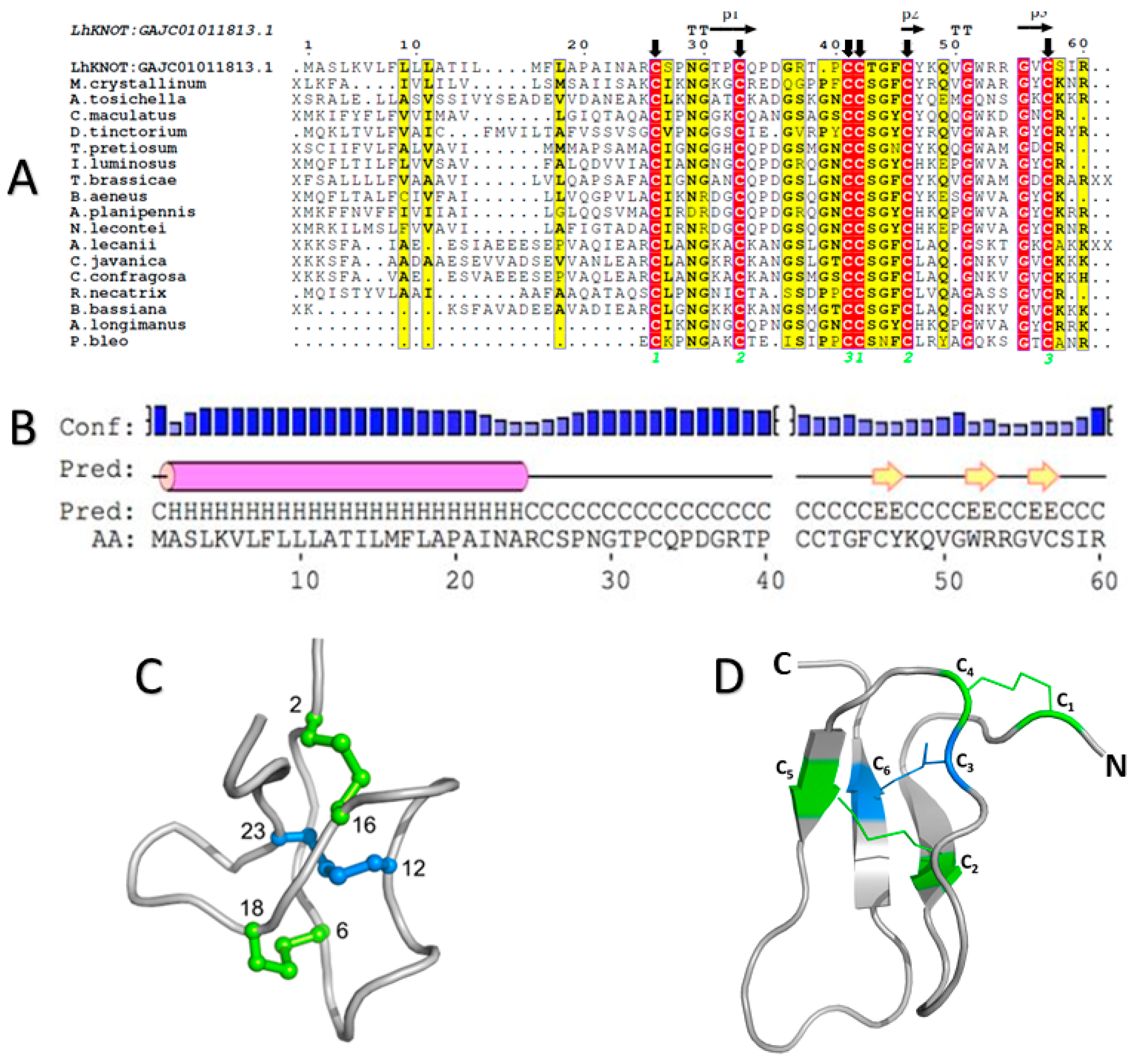{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The LhKNOT Sequence Shows Similarity to Insect and Plant Antimicrobial Peptides
2.2. The Three-Dimensional Structure of LhKNOT
2.3. LhKNOT Shows the Presence of a CPC Clip, Capable of Interacting with Heparin and Other GAGs
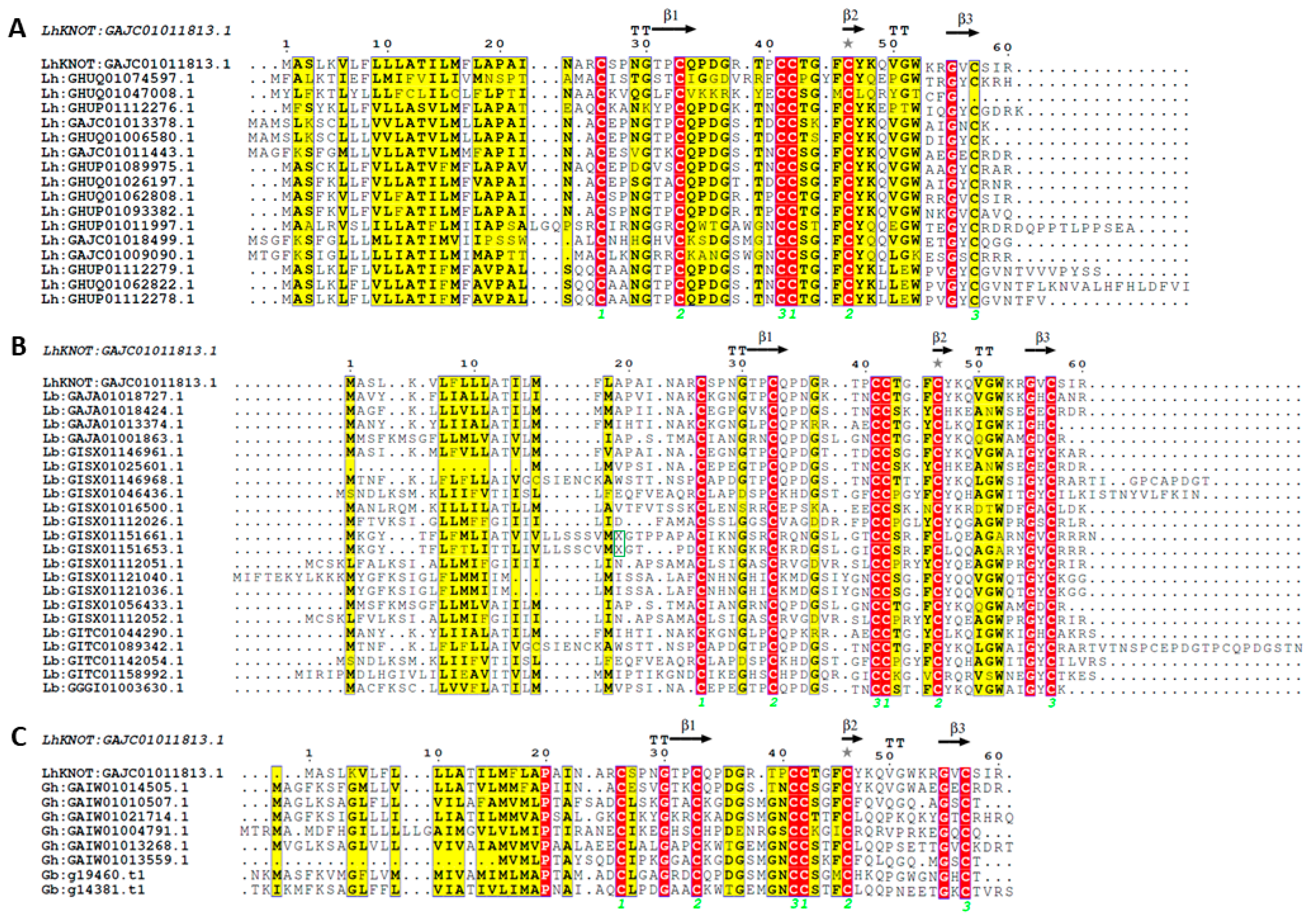
2.4. Other Structural Homologs of LhKNOT Also Contain the CPC Clip Motif
2.5. In Silico Mutational Analysis of LhKNOT CPC Clip Reveals a Secondary CPC Clip
2.6. A CPC Clip Is Also Present in the Antimicrobial Peptide Drosomycin
3. Discussion and Speculation
4. Methods
4.1. Sequence Analysis
4.2. LhKNOT Primary Structure Analysis
4.3. Tertiary Structure Prediction and Evaluation of LhKNOT and Its Homologs
4.4. CPC Clip Motif Identification
4.5. Association of LhKNOT with Other GAGs
4.6. Protein-Heparin Docking Analysis for Mutated CPC Clip in LhKNOT
4.7. Protein-Heparin Docking Analysis in Structural Homologs and Drosomycin
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Penczykowski, R.M.; Laine, A.L.; Koskella, B. Understanding the ecology and evolution of host-parasite interactions across scales. Evol. Appl. 2015, 9, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L.; Fellous, S.; Koella, J.C. Coevolutionary interactions between host and parasite genotypes. Trends Parasitol. 2006, 22, 12–16. [Google Scholar] [CrossRef]
- Quicke, D.L. Parasitic Wasps; Springer: Dordrecht, The Netherlands, 1997. [Google Scholar]
- Poirié, M.; Colinet, D.; Gatti, J. Insights into function and evolution of parasitoid wasp venoms. Curr. Opin. Insect Sci. 2014, 6, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Kraaijeveld, A.R.; Godfray, H.C.J. Evolution of host resistance and parasitoid counter-resistance. Adv. Parasitol. 2009, 70, 257–280. [Google Scholar] [CrossRef] [PubMed]
- Burke, G.R.; Strand, M.R. Systematic analysis of a wasp parasitism arsenal. Mol. Ecol. 2014, 23, 890–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.; Fang, Q.; Wang, L.; Liu, J.; Zhu, Y.; Wang, F.; Li, F.; Werren, J.H.; Ye, G. Insights into the venom composition and evolution of an endoparasitoid wasp by combining proteomic and transcriptomic analyses. Sci. Rep. 2016, 6, 19604. [Google Scholar] [CrossRef] [Green Version]
- Lue, C.; Driskell, A.C.; Leips, J.; Buffington, M.L. Review of the genus Leptopilina (Hymenoptera, Cynipoidea, Figitidae, Eucoilinae) from the Eastern United States, including three newly described species. J. Hymenopt. Res. 2016, 53, 35–76. [Google Scholar] [CrossRef] [Green Version]
- Heavner, M.E.; Hudgins, A.D.; Rajwani, R.; Morales, J.; Govind, S. Harnessing the natural Drosophila-parasitoid model for integrating insect immunity with functional venomics. Curr. Opin. Insect Sci. 2014, 6, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.J.; Kalamarz, M.E.; Paddibhatla, I.; Small, C.; Rajwani, R.; Govind, S. Virulence factors and strategies of Leptopilina spp.: Selective responses in Drosophila hosts. Adv. Parasitol. 2009, 70, 123–145. [Google Scholar] [CrossRef] [Green Version]
- Buchon, N.; Silverman, N.; Cherry, S. Immunity in Drosophila melanogaster—From microbial recognition to whole-organism physiology. Nat. Rev. Immunol. 2014, 14, 796–810. [Google Scholar] [CrossRef]
- Govind, S. Innate immunity in Drosophila: Pathogens and pathways. Insect Sci. 2008, 15, 29–43. [Google Scholar] [CrossRef]
- Kounatidis, I.; Ligoxygakis, P. Drosophila as a model system to unravel the layers of innate immunity to infection. Open Biol. 2012, 2, 120075. [Google Scholar] [CrossRef] [Green Version]
- Schlenke, T.A.; Morales, J.; Govind, S.; Clark, A.G. Contrasting infection strategies in generalist and specialist wasp parasitoids of Drosophila melanogaster. PLoS Pathog. 2007, 3, 1486–1501. [Google Scholar] [CrossRef]
- Qiu, P.; Pan, P.C.; Govind, S. A role for the Drosophila Toll/Cactus pathway in larval hematopoiesis. Development 1998, 125, 1909–1920. [Google Scholar] [CrossRef]
- Gueguen, G.; Kalamarz, M.E.; Ramroop, J.; Uribe, J.; Govind, S. Polydnaviral ankyrin proteins aid parasitic wasp survival by coordinate and selective inhibition of hematopoietic and immune NF-kappa B signaling in insect hosts. PLoS Pathog. 2013, 9, e1003580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louradour, I.; Sharma, A.; Morin-Poulard, I.; Letourneau, M.; Vincent, A.; Crozatier, M.; Vanzo, N. Reactive oxygen species-dependent Toll/NF-kappaB activation in the Drosophila hematopoietic niche confers resistance to wasp parasitism. eLife 2017, 6, e25496. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, R.P.; Melk, J.P.; Govind, S. Genetic analysis of contributions of dorsal group and JAK-Stat92E pathway genes to larval hemocyte concentration and the egg encapsulation response in Drosophila. Genetics 2004, 166, 1343–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, H.; Govind, S. Natural infection of D. melanogaster by virulent parasitic wasps induces apoptotic depletion of hematopoietic precursors. Cell Death Differ. 2002, 9, 1379–1381. [Google Scholar] [CrossRef] [Green Version]
- Rizki, R.M.; Rizki, T.M. Parasitoid virus-like particles destroy Drosophila cellular immunity. Proc. Natl. Acad. Sci. USA 1990, 87, 8388–8392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heavner, M.E.; Ramroop, J.; Gueguen, G.; Ramrattan, G.; Dolios, G.; Scarpati, M.; Kwiat, J.; Bhattacharya, S.; Wang, R.; Singh, S.; et al. Novel Organelles with Elements of Bacterial and Eukaryotic Secretion Systems Weaponize Parasites of Drosophila. Curr Biol. 2017, 27, 2869–2877.e6. [Google Scholar] [CrossRef] [PubMed]
- Ferrarese, R.; Morales, J.; Fimiarz, D.; Webb, B.A.; Govind, S. A supracellular system of actin-lined canals controls biogenesis and release of virulence factors in parasitoid venom glands. J. Exp. Biol. 2009, 212, 2261–2268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, H.; Morales, J.; Govind, S. Identification and immuno-electron microscopy localization of p40, a protein component of immunosuppressive virus-like particles from Leptopilina heterotoma, a virulent parasitoid wasp of Drosophila. J. Gen. Virol. 2006, 87, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Gracy, J.; Chiche, L. Structure and modeling of knottins, a promising molecular scaffold for drug discovery. Curr. Pharm. Des. 2011, 17, 4337–4350. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.H.; Liu, W.; Dai, J.X.; Wang, J.F.; Hu, Z.; Zhang, Y.; Wang, D.C. Solution structure of PAFP-S: A new knottin-type antifungal peptide from the seeds of Phytolacca americana. Biochemistry 2001, 40, 10973–10978. [Google Scholar] [CrossRef]
- Loo, S.; Kam, A.; Xiao, T.; Tam, J.P. Bleogens, Cactus-Derived Anti-Candida Cysteine-Rich Peptides with Three Different Precursor Arrangements. Front. Plant Sci. 2017, 8, 2162. [Google Scholar] [CrossRef] [Green Version]
- Becker, S.; Terlau, H. Toxins from cone snails: Properties, applications and biotechnological production. Appl. Microbiol. Biotechnol. 2008, 79, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reily, M.D.; Holub, K.E.; Gray, W.R.; Norris, T.M.; Adams, M.E. Structure-activity relationships for P-type calcium channel-selective omega-agatoxins. Nat. Struct. Biol. 1994, 1, 853–856. [Google Scholar] [CrossRef]
- Nguyen, K.N.T.; Nguyen, G.K.T.; Nguyen, P.Q.T.; Ang, K.H.; Dedon, P.C.; Tam, J.P. Immunostimulating and Gram-negative-specific antibacterial cyclotides from the butterfly pea (Clitoria ternatea). FEBS J. 2016, 283, 2067–2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbault, F.; Landon, C.; Guenneugues, M.; Meyer, J.P.; Schott, V.; Dimarcq, J.L.; Vovelle, F. Solution structure of Alo-3: A new knottin-type antifungal peptide from the insect Acrocinus longimanus. Biochemistry 2003, 42, 14434–14442. [Google Scholar] [CrossRef] [Green Version]
- Cox, N.; Kintzing, J.R.; Smith, M.; Grant, G.A.; Cochran, J.R. Integrin-Targeting Knottin Peptide-Drug Conjugates Are Potent Inhibitors of Tumor Cell Proliferation. Angew. Chem. Int. Ed. Engl. 2016, 55, 9894–9897. [Google Scholar] [CrossRef]
- Zhang, C.R.; Zhang, S.; Xia, J.; Li, F.F.; Xia, W.Q.; Liu, S.S.; Wang, X.W. The immune strategy and stress response of the Mediterranean species of the Bemisia tabaci complex to an orally delivered bacterial pathogen. PLoS ONE 2014, 9, e94477. [Google Scholar] [CrossRef] [PubMed]
- Shao, F.; Hu, Z.; Xiong, Y.M.; Huang, Q.Z.; Wang, C.G.; Zhu, R.H.; Wang, D.C. A new antifungal peptide from the seeds of Phytolacca americana: Characterization, amino acid sequence and cDNA cloning. Biochim. Biophys. Acta 1999, 1430, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.I.; Smith, R.; O’Donoghue, S.I.; Nilges, M.; Connor, M.; Howden, M.E.; Christie, M.J.; King, G.F. The structure of a novel insecticidal neurotoxin, omega-atracotoxin-HV1, from the venom of an Australian funnel web spider. Nat. Struct. Biol. 1997, 4, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Casale, J.; Crane, J.S. Biochemistry, Glycosaminoglycans. In Anonymous StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2022. [Google Scholar]
- Postic, G.; Gracy, J.; Périn, C.; Chiche, L.; Gelly, J. KNOTTIN: The database of inhibitor cystine knot scaffold after 10 years, toward a systematic structure modeling. Nucleic Acids Res. 2018, 46, D454–D458. [Google Scholar] [CrossRef] [Green Version]
- Soding, J.; Biegert, A.; Lupas, A.N. The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acids Res. 2005, 33 (Suppl. 2), 244. [Google Scholar] [CrossRef] [Green Version]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35, 407. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, D.; Luthy, R.; Bowie, J.U. VERIFY3D: Assessment of protein models with three-dimensional profiles. Methods Enzymol. 1997, 277, 396–404. [Google Scholar] [CrossRef]
- Olechnovič, K.; Venclovas, Č. VoroMQA: Assessment of protein structure quality using interatomic contact areas. Proteins 2017, 85, 1131–1145. [Google Scholar] [CrossRef]
- Pulido, D.; Rebollido-Rios, R.; Valle, J.; Andreu, D.; Boix, E.; Torrent, M. Structural similarities in the CPC clip motif explain peptide-binding promiscuity between glycosaminoglycans and lipopolysaccharides. J. R. Soc. Interface 2017, 14, 20170423. [Google Scholar] [CrossRef] [Green Version]
- Torrent, M.; Nogues, M.V.; Andreu, D.; Boix, E. The “CPC clip motif”: A conserved structural signature for heparin-binding proteins. PLoS ONE 2012, 7, e42692. [Google Scholar] [CrossRef]
- Landon, C.; Sodano, P.; Hetru, C.; Hoffmann, J.; Ptak, M. Solution structure of drosomycin, the first inducible antifungal protein from insects. Protein Sci. 1997, 6, 1878–1884. [Google Scholar] [CrossRef] [Green Version]
- Cornet, B.; Bonmatin, J.M.; Hetru, C.; Hoffmann, J.A.; Ptak, M.; Vovelle, F. Refined three-dimensional solution structure of insect defensin A. Structure 1995, 3, 435–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Illergard, K.; Ardell, D.H.; Elofsson, A. Structure is three to ten times more conserved than sequence—A study of structural response in protein cores. Proteins 2009, 77, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Wywial, E.; Singh, S.M. Identification and structural characterization of FYVE domain-containing proteins of Arabidopsis thaliana. BMC Plant Biol. 2010, 10, 157. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.L.; Singh, S.M. Identification, modeling, and characterization studies of Tetrahymena thermophila myosin FERM domains suggests a conserved core fold but functional differences. Cytoskeleton 2015, 72, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Wiech, E.M.; Cheng, H.P.; Singh, S.M. Molecular modeling and computational analyses suggests that the Sinorhizobium meliloti periplasmic regulator protein ExoR adopts a superhelical fold and is controlled by a unique mechanism of proteolysis. Protein Sci. 2015, 24, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Scarpati, M.; Qi, Y.; Govind, S.; Singh, S. A combined computational strategy of sequence and structural analysis predicts the existence of a functional eicosanoid pathway in Drosophila melanogaster. PLoS ONE 2019, 14, e0211897. [Google Scholar] [CrossRef] [Green Version]
- Barreto, C.; Silva, A.; Wiech, E.; Lopez, A.; San, A.; Singh, S. Proteomic Tools for the Analysis of Cytoskeleton Proteins. Methods Mol. Biol. 2022, 2364, 363–425. [Google Scholar] [CrossRef]
- Raskob, G.E.; Carter, C.J.; Hull, R.D. Heparin therapy for venous thrombosis and pulmonary embolism. Blood Rev. 1988, 2, 251–258. [Google Scholar] [CrossRef]
- Zhang, L. Glycosaminoglycan (GAG) biosynthesis and GAG-binding proteins. Prog. Mol. Biol. Transl. Sci. 2010, 93, 1–17. [Google Scholar] [CrossRef]
- Andersson, E.; Rydengard, V.; Sonesson, A.; Morgelin, M.; Bjorck, L.; Schmidtchen, A. Antimicrobial activities of heparin-binding peptides. Eur. J. Biochem. 2004, 271, 1219–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannuzzi, C.; Irace, G.; Sirangelo, I. The effect of glycosaminoglycans (GAGs) on amyloid aggregation and toxicity. Molecules 2015, 20, 2510–2528. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhu, S. Functional role of charged residues in drosomycin, a Drosophila antifungal peptide. Dev. Comp. Immunol. 2010, 34, 953–958. [Google Scholar] [CrossRef]
- Govind, S.; Nehm, R.H. Innate immunity in fruit flies: A textbook example of genomic recycling. PLoS Biol. 2004, 2, E276. [Google Scholar] [CrossRef] [PubMed]
- Araki, M.; Kurihara, M.; Kinoshita, S.; Awane, R.; Sato, T.; Ohkawa, Y.; Inoue, Y.H. Anti-tumour effects of antimicrobial peptides, components of the innate immune system, against haematopoietic tumours in Drosophila mxc mutants. Dis. Model Mech. 2019, 12, dmm037721. [Google Scholar] [CrossRef] [Green Version]
- Ramroop, J.R.; Heavner, M.E.; Razzak, Z.H.; Govind, S. A parasitoid wasp of Drosophila employs preemptive and reactive strategies to deplete its host’s blood cells. PLoS Pathog. 2021, 17, e1009615. [Google Scholar] [CrossRef]
- Zhu, F.; Li, D.; Chen, K. Structures and functions of invertebrate glycosylation. Open Biol. 2019, 9, 180232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iozzo, R.V.; Schaefer, L. Proteoglycan form and function: A comprehensive nomenclature of proteoglycans. Matrix Biol. 2015, 42, 11–55. [Google Scholar] [CrossRef]
- Alexander, C.; Rietschel, E.T. Bacterial lipopolysaccharides and innate immunity. J. Endotoxin Res. 2001, 7, 167–202. [Google Scholar] [CrossRef]
- Baron, M.J.; Wong, S.L.; Nybakken, K.; Carey, V.J.; Madoff, L.C. Host glycosaminoglycan confers susceptibility to bacterial infection in Drosophila melanogaster. Infect Immun. 2009, 77, 860–866. [Google Scholar] [CrossRef]
- Imler, J.; Bulet, P. Antimicrobial peptides in Drosophila: Structures, activities and gene regulation. Chem Immunol Allerg. 2005, 86, 1–21. [Google Scholar] [CrossRef]
- Hwang, B.; Hwang, J.; Lee, J.; Lee, D.G. Antifungal properties and mode of action of psacotheasin, a novel knottin-type peptide derived from Psacothea hilaris. Biochem. Biophys. Res. Commun. 2010, 400, 352–357. [Google Scholar] [CrossRef]
- De Guillen, K.; Lorrain, C.; Tsan, P.; Barthe, P.; Petre, B.; Saveleva, N.; Rouhier, N.; Duplessis, S.; Padilla, A.; Hecker, A. Structural genomics applied to the rust fungus Melampsora larici-populina reveals two candidate effector proteins adopting cystine knot and NTF2-like protein folds. Sci. Rep. 2019, 9, 18084–18089. [Google Scholar] [CrossRef] [Green Version]
- NCBI Resource Coordinators. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2018, 46, D8–D13. [Google Scholar] [CrossRef] [Green Version]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeira, F.; Pearce, M.; Tivey, A.R.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, 276. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, 29. [Google Scholar] [CrossRef] [Green Version]
- Goecks, J.; Mortimer, N.T.; Mobley, J.A.; Bowersock, G.J.; Taylor, J.; Schlenke, T.A. Integrative approach reveals composition of endoparasitoid wasp venoms. PLoS ONE 2013, 8, e64125. [Google Scholar] [CrossRef] [PubMed]
- Varaldi, J.; Lepetit, D. Deciphering the behaviour manipulation imposed by a virus on its parasitoid host: Insights from a dual transcriptomic approach. Parasitology 2018, 145, 1979–1989. [Google Scholar] [CrossRef]
- Mortimer, N.T.; Goecks, J.; Kacsoh, B.Z.; Mobley, J.A.; Bowersock, G.J.; Taylor, J.; Schlenke, T.A. Parasitoid wasp venom SERCA regulates Drosophila calcium levels and inhibits cellular immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 9427–9432. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Keller, O.; Kollmar, M.; Stanke, M.; Waack, S. A novel hybrid gene prediction method employing protein multiple sequence alignments. Bioinformatics 2011, 27, 757–763. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Poirot, O.; O’Toole, E.; Notredame, C. Tcoffee@ igs: A web server for computing, evaluating and combining multiple sequence alignments. Nucleic Acids Res. 2003, 31, 3503–3506. [Google Scholar] [CrossRef] [PubMed]
- Gouet, P.; Courcelle, E.; Stuart, D.I.; Metoz, F. ESPript: Analysis of multiple sequence alignments in PostScript. Bioinformatics 1999, 15, 305–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchler-Bauer, A.; Lu, S.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; et al. CDD: A Conserved Domain Database for the functional annotation of proteins. Nucleic Acids Res. 2011, 39, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, 222. [Google Scholar] [CrossRef] [Green Version]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [Green Version]
- Kall, L.; Krogh, A.; Sonnhammer, E.L. Advantages of combined transmembrane topology and signal peptide prediction—The Phobius web server. Nucleic Acids Res. 2007, 35, 429. [Google Scholar] [CrossRef] [Green Version]
- Simossis, V.A.; Heringa, J. The influence of gapped positions in multiple sequence alignments on secondary structure prediction methods. Comput. Biol. Chem. 2004, 28, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, B.; Sali, A. Comparative Protein Structure Modeling Using MODELLER. Curr. Protoc. Bioinform. 2014, 47, 5–6. [Google Scholar] [CrossRef] [Green Version]
- Maiti, R.; Van Domselaar, G.H.; Zhang, H.; Wishart, D.S. SuperPose: A simple server for sophisticated structural superposition. Nucleic Acids Res. 2004, 32, 590. [Google Scholar] [CrossRef] [Green Version]
- Shatsky, M.; Nussinov, R.; Wolfson, H.J. A method for simultaneous alignment of multiple protein structures. Proteins 2004, 56, 143–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vajda, S.; Yueh, C.; Beglov, D.; Bohnuud, T.; Mottarella, S.E.; Xia, B.; Hall, D.R.; Kozakov, D. New additions to the ClusPro server motivated by CAPRI. Proteins 2017, 85, 435–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrodinger LLC. The PyMOL Molecular Graphics System; Version 1.8; Schrodinger, LLC: New York, NY, USA, 2015; in press. [Google Scholar]
- Baker, N.A.; Sept, D.; Joseph, S.; Holst, M.J.; McCammon, J.A. Electrostatics of nanosystems: Application to microtubules and the ri-bosome. Proc. Natl. Acad. Sci USA 2001, 98, 10037–10041. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Jabłońska, J.; Pravda, L.; Vařeková, R.S.; Thornton, J.M. PDBsum: Structural summaries of PDB entries. Protein Sci. 2018, 27, 129–134. [Google Scholar] [CrossRef]
- Lippens, G.; Najib, J.; Wodak, S.J.; Tartar, A. NMR sequential assignments and solution structure of chlorotoxin, a small scorpion toxin that blocks chloride channels. Biochemistry 1995, 34, 13–21. [Google Scholar] [CrossRef]
- Takahashi, H.; Kim, J.I.; Min, H.J.; Sato, K.; Swartz, K.J.; Shimada, I. Solution structure of hanatoxin1, a gating modifier of voltage-dependent K(+) channels: Common surface features of gating modifier toxins. J. Mol. Biol. 2000, 297, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Landon, C.; Pajon, A.; Vovelle, F.; Sodano, P. The active site of drosomycin, a small insect antifungal protein, delineated by comparison with the modeled structure of Rs-AFP2, a plant antifungal protein. J. Pept. Res. 2000, 56, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.M.; Alewood, P.F.; Craik, D.J. Solution structure of the sodium channel antagonist conotoxin GS: A new molecular caliper for probing sodium channel geometry. Structure 1997, 5, 571–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, K.J.; Thomas, L.; Lewis, R.J.; Alewood, P.F.; Craik, D.J. A consensus structure for omega-conotoxins with different selectivities for voltage-sensitive calcium channel subtypes: Comparison of MVIIA, SVIB and SNX-202. J. Mol. Biol. 1996, 263, 297–310. [Google Scholar] [CrossRef]
- Scanlon, M.J.; Naranjo, D.; Thomas, L.; Alewood, P.F.; Lewis, R.J.; Craik, D.J. Solution structure and proposed binding mechanism of a novel potassium channel toxin kappa-conotoxin PVIIA. Structure 1997, 5, 1585–1597. [Google Scholar] [CrossRef] [Green Version]
- Fujitani, N.; Kawabata, S.; Osaki, T.; Kumaki, Y.; Demura, M.; Nitta, K. Structure of the antimicrobial peptide tachystatin A. J. Biol. Chem. 2002, 277, 23651–23657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujitani, N.; Kouno, T.; Nakahara, T.; Takaya, K.; Osaki, T.; Kawabata, S. The solution structure of horseshoe crab antimicrobial peptide tachystatin B with an inhibitory cystine-knot motif. J. Pept Sci. 2007, 13, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Bernard, C.; Corzo, G.; Mosbah, A.; Nakajima, T.; Darbon, H. Solution structure of Ptu1, a toxin from the assassin bug Peirates turpis that blocks the voltage-sensitive calcium channel N-type. Biochemistry 2001, 40, 12795–12800. [Google Scholar] [CrossRef]
- Bernard, C.; Corzo, G.; Adachi-Akahane, S.; Foures, G.; Kanemaru, K.; Furukawa, Y. Solution structure of ADO1, a toxin extracted from the saliva of the assassin bug, Agriosphodrus dohrni. Proteins 2004, 54, 195–205. [Google Scholar] [CrossRef]
- Martins, J.C.; Enassar, M.; Willem, R.; Wieruzeski, J.M.; Lippens, G.; Wodak, S.J. Solution structure of the main alpha-amylase inhibitor from amaranth seeds. Eur. J. Biochem. 2001, 268, 2379–2389. [Google Scholar] [CrossRef] [PubMed]
- Janssen, B.J.C.; Schirra, H.J.; Lay, F.T.; Anderson, M.A.; Craik, D.J. Structure of Petunia hybrida defensin 1, a novel plant defensin with five disulfide bonds. Biochemistry 2003, 42, 8214–8222. [Google Scholar] [CrossRef]
- Frenal, K.; Xu, C.; Wolff, N.; Wecker, K.; Gurrola, G.B.; Zhu, S.; Chi, C.-W.; Possani, L.D.; Tytgat, J.; Delepierre, M. Exploring structural features of the interaction between the scorpion toxinCnErg1 and ERG K+ channels. Proteins 2004, 56, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Lomize, A.L.; Maiorov, V.N.; Arsen’ev, A.S. Determination of the spatial structure of insectotoxin 15A from Buthus erpeus by (1)H-NMR spectroscopy data. Bioorg Khim. 1991, 17, 1613–1632. [Google Scholar]
- Pimentel, C.; Choi, S.; Chagot, B.; Guette, C.; Camadro, J.; Darbon, H. Solution structure of PcFK1, a spider peptide active against Plasmodium falciparum. Protein Sci. 2006, 15, 628–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koltay, A.; Daly, N.L.; Gustafson, K.R.; Craik, D.J. Structure of Circulin B and Implications for Antimicrobial Activity of the Cyclotides. Int. J. Pept. Res. Ther. 2005, 11, 99–106. [Google Scholar] [CrossRef]
- Craik, D.J.; Daly, N.L.; Bond, T.; Waine, C. Plant cyclotides: A unique family of cyclic and knotted proteins that defines the cyclic cystine knot structural motif. J. Mol. Biol. 1999, 294, 1327–1336. [Google Scholar] [CrossRef]
- Li, H.; Bowling, J.J.; Fronczek, F.R.; Hong, J.; Jabba, S.V.; Murray, T.F.; Ha, N.-C.; Hamann, M.T.; Jung, J.H. Asteropsin A: An unusual cystine-crosslinked peptide from porifera enhances neuronal Ca2+ influx. Biochim Biophys Acta. 2013, 1830, 2591–2599. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arguelles, J.; Lee, J.; Cardenas, L.V.; Govind, S.; Singh, S. In Silico Analysis of a Drosophila Parasitoid Venom Peptide Reveals Prevalence of the Cation–Polar–Cation Clip Motif in Knottin Proteins. Pathogens 2023, 12, 143. https://doi.org/10.3390/pathogens12010143
Arguelles J, Lee J, Cardenas LV, Govind S, Singh S. In Silico Analysis of a Drosophila Parasitoid Venom Peptide Reveals Prevalence of the Cation–Polar–Cation Clip Motif in Knottin Proteins. Pathogens. 2023; 12(1):143. https://doi.org/10.3390/pathogens12010143
Chicago/Turabian StyleArguelles, Joseph, Jenny Lee, Lady V. Cardenas, Shubha Govind, and Shaneen Singh. 2023. "In Silico Analysis of a Drosophila Parasitoid Venom Peptide Reveals Prevalence of the Cation–Polar–Cation Clip Motif in Knottin Proteins" Pathogens 12, no. 1: 143. https://doi.org/10.3390/pathogens12010143