
Adaptive Evolution Compensated for the Plasmid Fitness Costs Brought by Specific Genetic Conflicts
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmid
2.2. Growth Curve Assay
2.3. Competition Experiments
2.4. Experimental Evolution
2.5. Whole-Genome Sequencing and Bioinformatic Analysis
2.6. Statistical Analysis
3. Results
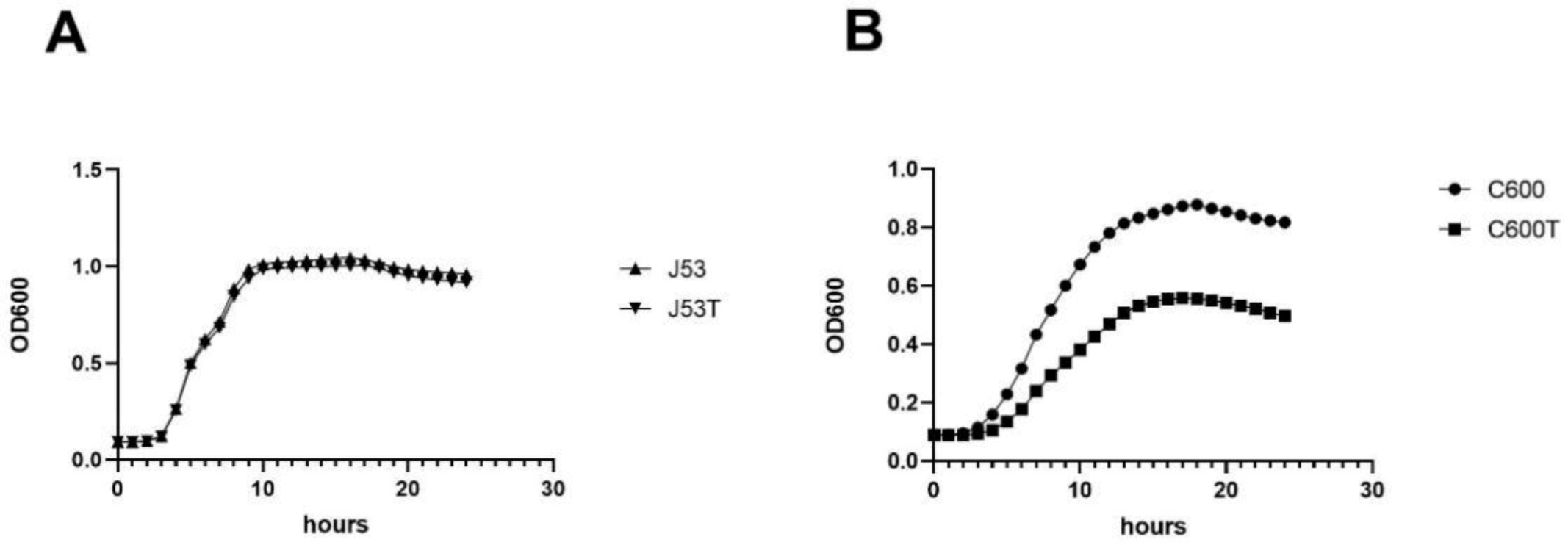
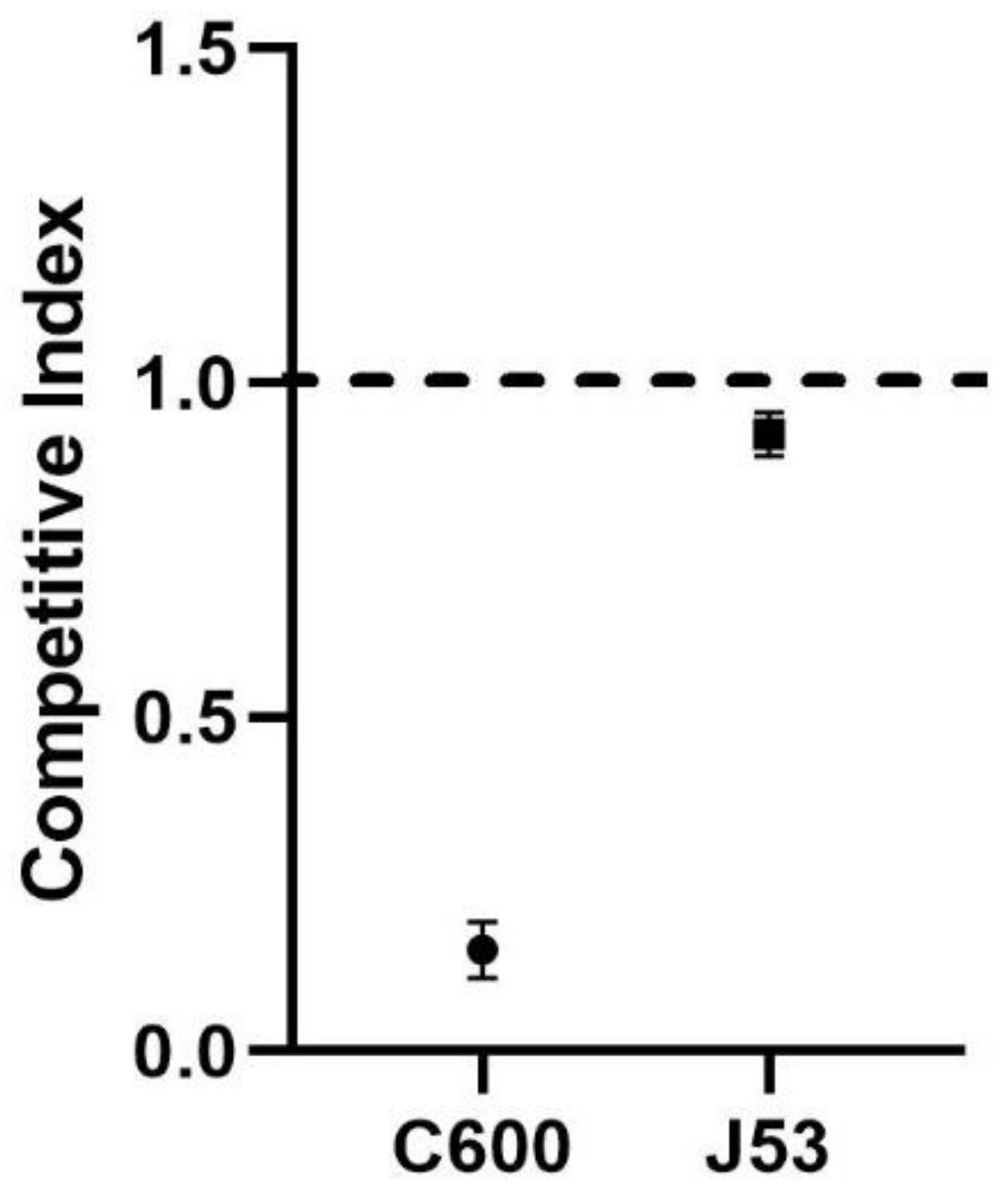
3.1. Fitness Cost of Carrying IncX3 Plasmid
3.2. Comparative Analysis of E. coli C600 and E. coli C600/pHN330
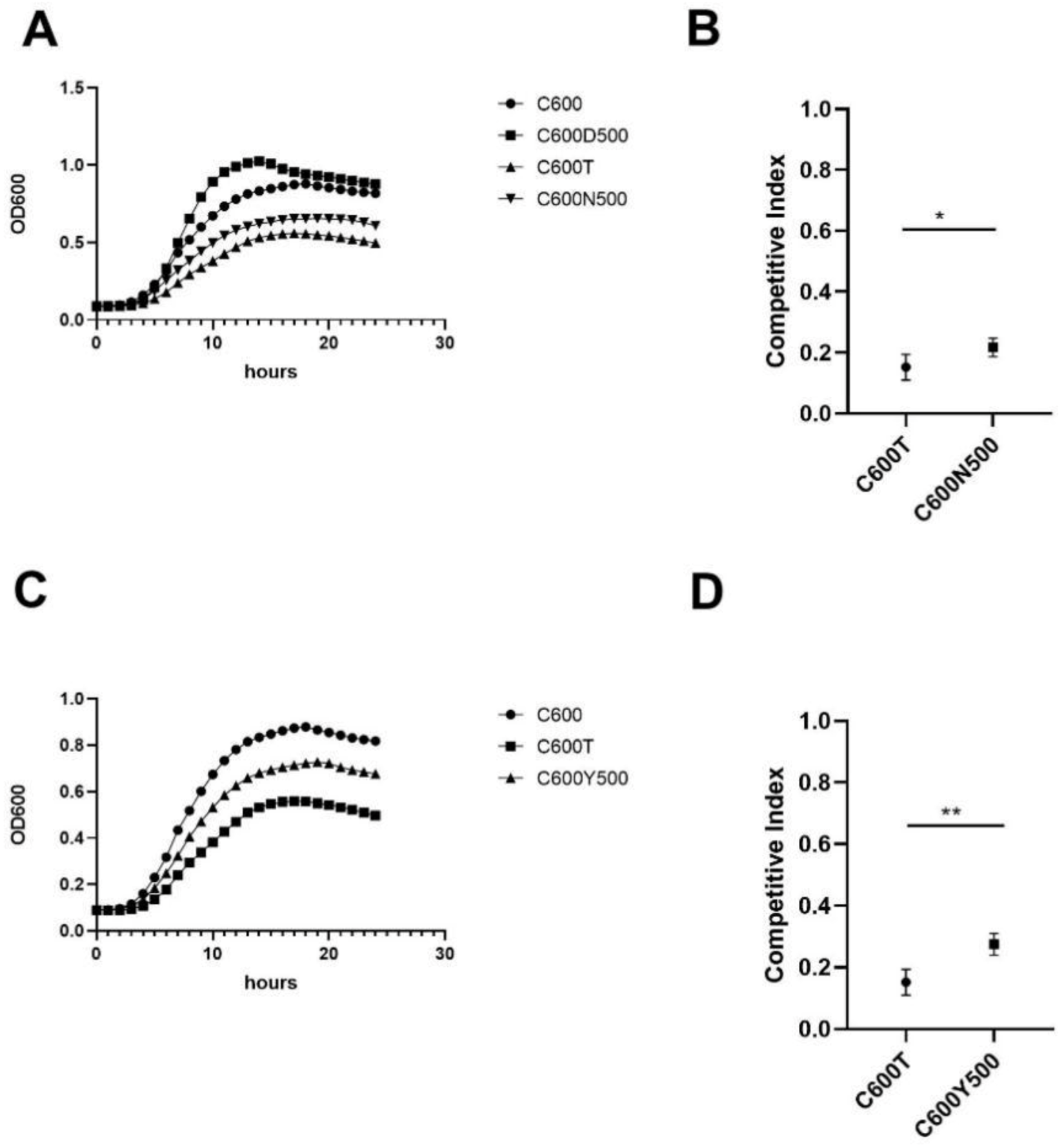
3.3. IncX3 Plasmid Maintenance in E. coli C600 after 500 Passages
3.4. Fitness Cost of Progeny Strains Decreased Compared to C600T Strain
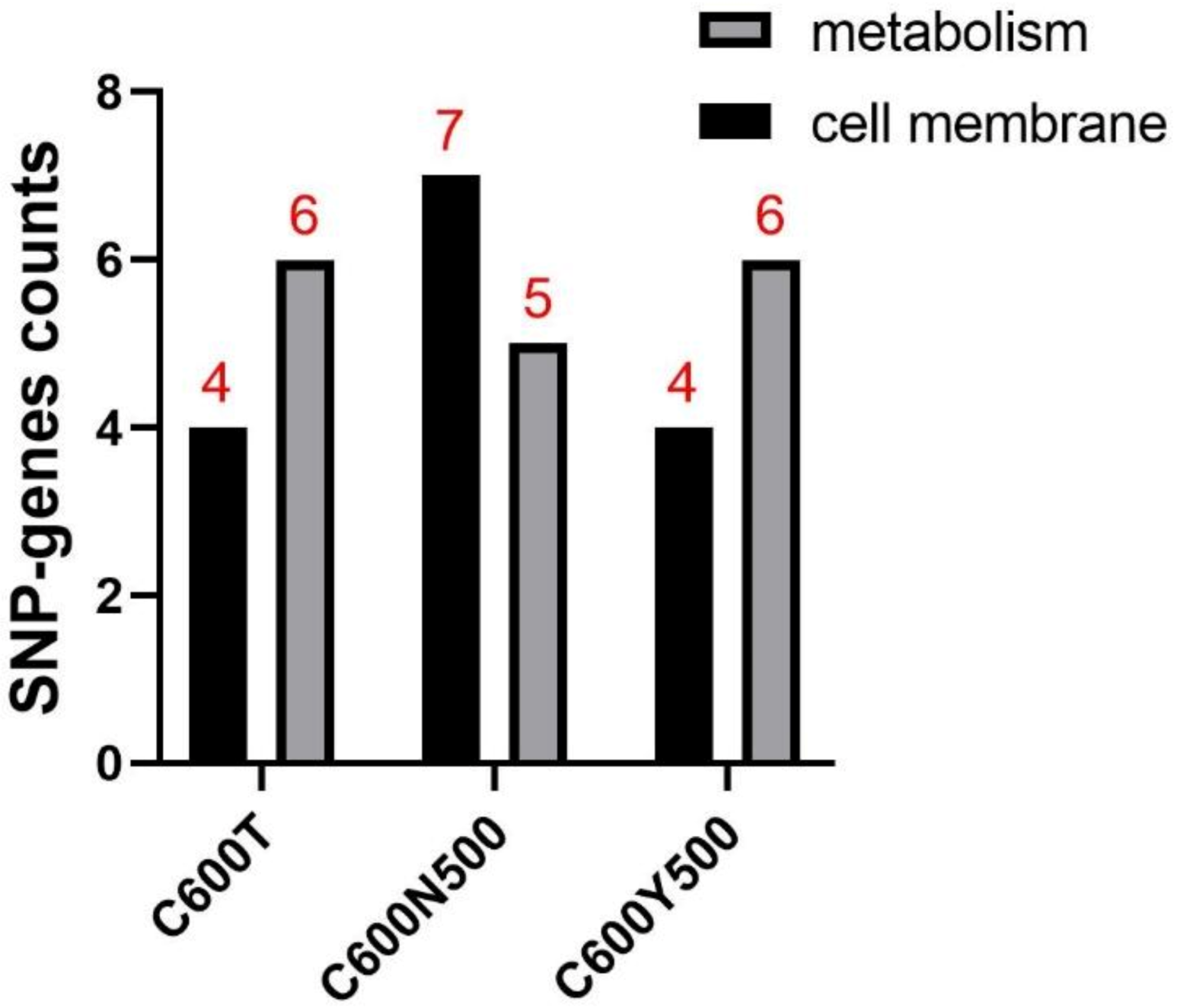
3.5. SNPs Occurring in the CDS Region Reduced the Fitness Cost of C600T
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046–5054. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Berglund, B.; Zou, H.; Zhou, Z.; Xia, H.; Zhao, L.; Nilsson, L.E.; Li, X. Dissemination of bla(NDM-5) via IncX3 plasmids in carbapenem-resistant Enterobacteriaceae among humans and in the environment in an intensive vegetable cultivation area in eastern China. Environ. Pollut. 2021, 273, 116370. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Feng, Y.; Tang, G.; Qiao, F.; McNally, A.; Zong, Z. NDM Metallo-β-Lactamases and Their Bacterial Producers in Health Care Settings. Clin. Microbiol. Rev. 2019, 32, e00115-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- San Millan, A.; MacLean, R.C. Fitness Costs of Plasmids: A Limit to Plasmid Transmission. Microbiol. Spectr. 2017, 5, 5.5.02. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltrus, D.A. Exploring the costs of horizontal gene transfer. Trends Ecol. Evol. 2013, 28, 489–495. [Google Scholar] [CrossRef]
- Zünd, P.; Lebek, G. Generation time-prolonging R plasmids: Correlation between increases in the generation time of Escherichia coli caused by R plasmids and their molecular size. Plasmid 1980, 3, 65–69. [Google Scholar] [CrossRef]
- Harrison, E.; Brockhurst, M.A. Plasmid-mediated horizontal gene transfer is a coevolutionary process. Trends Microbiol. 2012, 20, 262–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baomo, L.; Lili, S.; Moran, R.A.; van Schaik, W.; Chao, Z. Temperature-Regulated IncX3 Plasmid Characteristics and the Role of Plasmid-Encoded H-NS in Thermoregulation. Front. Microbiol. 2021, 12, 765492. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Guo, Y.; Liu, N.; Wang, J.; Li, F.; Yao, L.; Zhuo, C. In silico Evolution and Comparative Genomic Analysis of IncX3 Plasmids Isolated From China Over Ten Years. Front. Microbiol. 2021, 12, 725391. [Google Scholar] [CrossRef]
- Hammer-Dedet, F.; Aujoulat, F.; Jumas-Bilak, E.; Licznar-Fajardo, P. Persistence and Dissemination Capacities of a blaNDM-5-Harboring IncX-3 Plasmid in Escherichia coli Isolated from an Urban River in Montpellier, France. Antibiotics 2022, 11, 196. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liao, K.; Gao, H.; Wang, Q.; Wang, X.; Li, H.; Wang, R.; Wang, H. Decreased Fitness and Virulence in ST10 Escherichia coli Harboring blaNDM-5 and mcr-1 against a ST4981 Strain with blaNDM-5. Front. Cell Infect. Microbiol. 2017, 7, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, T.; Wei, R.; Zhang, L.; Sun, L.; Pang, M.; Wang, R.; Wang, Y. Characterization of NDM-5-positive extensively resistant Escherichia coli isolates from dairy cows. Vet. Microbiol. 2017, 207, 153–158. [Google Scholar] [CrossRef]
- Ma, T.; Fu, J.; Xie, N.; Ma, S.; Lei, L.; Zhai, W.; Shen, Y.; Sun, C.; Wang, S.; Shen, Z.; et al. Fitness Cost of blaNDM-5-Carrying p3R-IncX3 Plasmids in Wild-Type NDM-Free Enterobacteriaceae. Microorganisms 2020, 8, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.P.J.; Wright, R.C.T.; Harrison, E.; Muddiman, K.J.; Wood, A.J.; Paterson, S.; Brockhurst, M.A. Plasmid fitness costs are caused by specific genetic conflicts enabling resolution by compensatory mutation. PLoS Biol. 2021, 19, e3001225. [Google Scholar] [CrossRef]
- Haudiquet, M.; de Sousa, J.M.; Touchon, M.; Rocha, E.P.C. Selfish, promiscuous and sometimes useful: How mobile genetic elements drive horizontal gene transfer in microbial populations. Philos. Trans. R Soc. Lond. B Biol. Sci. 2022, 377, 20210234. [Google Scholar] [CrossRef]
- Mc Ginty, S.; Rankin, D.J. The evolution of conflict resolution between plasmids and their bacterial hosts. Evolution 2012, 66, 1662–1670. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.F.; Fang, L.X.; Chang, M.X.; Cheng, M.; Zhang, H.; Long, T.F.; Li, Q.; Lian, X.L.; Sun, J.; Liao, X.P.; et al. A Trade-Off for Maintenance of Multidrug-Resistant IncHI2 Plasmids in Salmonella enterica Serovar Typhimurium through Adaptive Evolution. mSystems 2022, 7, e0024822. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H. Selfish genetic elements, genetic conflict, and evolutionary innovation. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 2), 10863–10870. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Shui, L.; Zhou, K.; Jiang, Y.; Li, X.; Guan, J.; Li, Q.; Zhuo, C. Impact of Plasmid-Encoded H-NS-like Protein on blaNDM-1-Bearing IncX3 Plasmid in Escherichia coli. J. Infect. Dis. 2020, 221, S229–S236. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance standards for antimicrobial susceptibility testing. CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2021. [Google Scholar]
- Yang, S.; Lopez, C.R.; Zechiedrich, E.L. Quorum sensing and multidrug transporters in Escherichia coli. Proc. Natl. Acad. Sci. USA 2006, 103, 2386–2391. [Google Scholar] [CrossRef]
- Liu, D.; Liu, Z.S.; Hu, P.; Cai, L.; Fu, B.Q.; Li, Y.S.; Lu, S.Y.; Liu, N.N.; Ma, X.L.; Chi, D.; et al. Characterization of surface antigen protein 1 (SurA1) from Acinetobacter baumannii and its role in virulence and fitness. Vet. Microbiol. 2016, 186, 126–138. [Google Scholar] [CrossRef]
- Trinetta, V.; Magossi, G.; Allard, M.W.; Tallent, S.M.; Brown, E.W.; Lomonaco, S. Characterization of Salmonella enterica Isolates from Selected U.S. Swine Feed Mills by Whole-Genome Sequencing. Foodborne Pathog. Dis. 2020, 17, 126–136. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olawoye, I.B.; Frost, S.D.W.; Happi, C.T. The Bacteria Genome Pipeline (BAGEP): An automated, scalable workflow for bacteria genomes with Snakemake. PeerJ 2020, 8, e10121. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckner, M.M.C.; Saw, H.T.H.; Osagie, R.N.; McNally, A.; Ricci, V.; Wand, M.E.; Woodford, N.; Ivens, A.; Webber, M.A.; Piddock, L.J.V. Clinically Relevant Plasmid-Host Interactions Indicate that Transcriptional and Not Genomic Modifications Ameliorate Fitness Costs of Klebsiella pneumoniae Carbapenemase-Carrying Plasmids. mBio 2018, 9, e02303-17. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Guo, Z.W.; Zhi, C.P.; Yang, T.; Zhao, J.J.; Chen, X.J.; Zeng, L.; Lv, L.C.; Zeng, Z.L.; Liu, J.H. Impact of plasmid-borne oqxAB on the development of fluoroquinolone resistance and bacterial fitness in Escherichia coli. J. Antimicrob. Chemother. 2017, 72, 1293–1302. [Google Scholar] [CrossRef] [Green Version]
- Murarka, A.; Clomburg, J.M.; Moran, S.; Shanks, J.V.; Gonzalez, R. Metabolic analysis of wild-type Escherichia coli and a pyruvate dehydrogenase complex (PDHC)-deficient derivative reveals the role of PDHC in the fermentative metabolism of glucose. J. Biol. Chem. 2010, 285, 31548–31558. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, R. Analysis of melibiose mutants deficient in alpha-galactosidase and thiomethylgalactoside permease II in Escherichia coli K-12. J. Bacteriol. 1968, 96, 462–471. [Google Scholar] [CrossRef]
- Lengeler, J. Mutations affecting transport of the hexitols D-mannitol, D-glucitol, and galactitol in Escherichia coli K-12: Isolation and mapping. J. Bacteriol. 1975, 124, 26–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denger, K.; Weiss, M.; Felux, A.K.; Schneider, A.; Mayer, C.; Spiteller, D.; Huhn, T.; Cook, A.M.; Schleheck, D. Sulphoglycolysis in Escherichia coli K-12 closes a gap in the biogeochemical sulphur cycle. Nature 2014, 507, 114–117. [Google Scholar] [CrossRef] [Green Version]
- Voulhoux, R.; Bos, M.P.; Geurtsen, J.; Mols, M.; Tommassen, J. Role of a highly conserved bacterial protein in outer membrane protein assembly. Science 2003, 299, 262–265. [Google Scholar] [CrossRef]
- Rodríguez-Alonso, R.; Létoquart, J.; Nguyen, V.S.; Louis, G.; Calabrese, A.N.; Iorga, B.I.; Radford, S.E.; Cho, S.H.; Remaut, H.; Collet, J.F. Structural insight into the formation of lipoprotein-β-barrel complexes. Nat. Chem. Biol. 2020, 16, 1019–1025. [Google Scholar] [CrossRef]
- Fiedler, W.; Rotering, H. Properties of Escherichia coli mutants lacking membrane-derived oligosaccharides. J. Biol. Chem. 1988, 263, 14684–14689. [Google Scholar] [CrossRef]
- Bossemeyer, D.; Borchard, A.; Dosch, D.C.; Helmer, G.C.; Epstein, W.; Booth, I.R.; Bakker, E.P. K+-transport protein TrkA of Escherichia coli is a peripheral membrane protein that requires other trk gene products for attachment to the cytoplasmic membrane. J. Biol. Chem. 1989, 264, 16403–16410. [Google Scholar] [CrossRef] [PubMed]
- San Millan, A.; Toll-Riera, M.; Qi, Q.; Betts, A.; Hopkinson, R.J.; McCullagh, J.; MacLean, R.C. Integrative analysis of fitness and metabolic effects of plasmids in Pseudomonas aeruginosa PAO1. ISME J. 2018, 12, 3014–3024. [Google Scholar] [CrossRef] [PubMed]
- Subbiah, M.; Top, E.M.; Shah, D.H.; Call, D.R. Selection pressure required for long-term persistence of blaCMY-2-positive IncA/C plasmids. Appl. Environ. Microbiol. 2011, 77, 4486–4493. [Google Scholar] [CrossRef] [Green Version]
- Lang, K.S.; Johnson, T.J. Transcriptome modulations due to A/C2 plasmid acquisition. Plasmid 2015, 80, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Loftie-Eaton, W.; Bashford, K.; Quinn, H.; Dong, K.; Millstein, J.; Hunter, S.; Thomason, M.K.; Merrikh, H.; Ponciano, J.M.; Top, E.M. Compensatory mutations improve general permissiveness to antibiotic resistance plasmids. Nat. Ecol. Evol. 2017, 1, 1354–1363. [Google Scholar] [CrossRef]
- Noinaj, N.; Gumbart, J.C.; Buchanan, S.K. The beta-barrel assembly machinery in motion. Nat. Rev. Microbiol. 2017, 15, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhu, X.; Zhang, J.; Lin, Y.; You, X.; Chen, M.; Wang, Y.; Zhu, N.; Si, S. Identification of a Compound That Inhibits the Growth of Gram-Negative Bacteria by Blocking BamA-BamD Interaction. Front. Microbiol. 2020, 11, 1252. [Google Scholar] [CrossRef]
- Storek, K.M.; Auerbach, M.R.; Shi, H.; Garcia, N.K.; Sun, D.; Nickerson, N.N.; Vij, R.; Lin, Z.; Chiang, N.; Schneider, K.; et al. Monoclonal antibody targeting the β-barrel assembly machine of Escherichia coli is bactericidal. Proc. Natl. Acad. Sci. USA 2018, 115, 3692–3697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noinaj, N.; Kuszak, A.J.; Gumbart, J.C.; Lukacik, P.; Chang, H.; Easley, N.C.; Lithgow, T.; Buchanan, S.K. Structural insight into the biogenesis of β-barrel membrane proteins. Nature 2013, 501, 385–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.; Malinverni, J.; Ruiz, N.; Kim, S.; Silhavy, T.J.; Kahne, D. Identification of a multicomponent complex required for outer membrane biogenesis in Escherichia coli. Cell 2005, 121, 235–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gawel, D.; Hamilton, M.D.; Schaaper, R.M. A novel mutator of Escherichia coli carrying a defect in the dgt gene, encoding a dGTP triphosphohydrolase. J. Bacteriol. 2008, 190, 6931–6939. [Google Scholar] [CrossRef] [Green Version]
- Thongsomboon, W.; Serra, D.O.; Possling, A.; Hadjineophytou, C.; Hengge, R.; Cegelski, L. Phosphoethanolamine cellulose: A naturally produced chemically modified cellulose. Science 2018, 359, 334–338. [Google Scholar] [CrossRef] [Green Version]
- Sherman, D.J.; Xie, R.; Taylor, R.J.; George, A.H.; Okuda, S.; Foster, P.J.; Needleman, D.J.; Kahne, D. Lipopolysaccharide is transported to the cell surface by a membrane-to-membrane protein bridge. Science 2018, 359, 798–801. [Google Scholar] [CrossRef] [Green Version]
- Brabetz, W.; Müller-Loennies, S.; Holst, O.; Brade, H. Deletion of the heptosyltransferase genes rfaC and rfaF in Escherichia coli K-12 results in an Re-type lipopolysaccharide with a high degree of 2-aminoethanol phosphate substitution. Eur. J. Biochem. 1997, 247, 716–724. [Google Scholar] [CrossRef]
- Cote, J.M.; Taylor, E.A. The Glycosyltransferases of LPS Core: A Review of Four Heptosyltransferase Enzymes in Context. Int. J. Mol. Sci. 2017, 18, 2256. [Google Scholar] [CrossRef]
- Kruse, T.; Bork-Jensen, J.; Gerdes, K. The morphogenetic MreBCD proteins of Escherichia coli form an essential membrane-bound complex. Mol. Microbiol. 2005, 55, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Eger, E.; Schwabe, M.; Schulig, L.; Hübner, N.O.; Bohnert, J.A.; Bornscheuer, U.T.; Heiden, S.E.; Müller, J.U.; Adnan, F.; Becker, K.; et al. Extensively Drug-Resistant Klebsiella pneumoniae Counteracts Fitness and Virulence Costs That Accompanied Ceftazidime-Avibactam Resistance Acquisition. Microbiol. Spectr. 2022, 10, e0014822. [Google Scholar] [CrossRef] [PubMed]
- Harrison, E.; Guymer, D.; Spiers, A.J.; Paterson, S.; Brockhurst, M.A. Parallel compensatory evolution stabilizes plasmids across the parasitism-mutualism continuum. Curr. Biol. 2015, 25, 2034–2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; de Bruijn, I.; van der Voort, M.; Loper, J.E.; Raaijmakers, J.M. The Gac regulon of Pseudomonas fluorescens SBW25. Environ. Microbiol. Rep. 2013, 5, 608–619. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Feng, Y.; Zong, Z. Fitness cost of a mcr-1-carrying IncHI2 plasmid. PLoS One 2018, 13, e0209706. [Google Scholar] [CrossRef] [Green Version]
- Rangarajan, E.S.; Li, Y.; Ajamian, E.; Iannuzzi, P.; Kernaghan, S.D.; Fraser, M.E.; Cygler, M.; Matte, A. Crystallographic trapping of the glutamyl-CoA thioester intermediate of family I CoA transferases. J. Biol. Chem. 2005, 280, 42919–42928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mironov, A.; Seregina, T.; Shatalin, K.; Nagornykh, M.; Shakulov, R.; Nudler, E. CydDC functions as a cytoplasmic cystine reductase to sensitize Escherichia coli to oxidative stress and aminoglycosides. Proc. Natl. Acad. Sci. USA 2020, 117, 23565–23570. [Google Scholar] [CrossRef]
- Ohtsu, I.; Wiriyathanawudhiwong, N.; Morigasaki, S.; Nakatani, T.; Kadokura, H.; Takagi, H. The L-cysteine/L-cystine shuttle system provides reducing equivalents to the periplasm in Escherichia coli. J. Biol. Chem. 2010, 285, 17479–17487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clugston, S.L.; Barnard, J.F.; Kinach, R.; Miedema, D.; Ruman, R.; Daub, E.; Honek, J.F. Overproduction and characterization of a dimeric non-zinc glyoxalase I from Escherichia coli: Evidence for optimal activation by nickel ions. Biochemistry 1998, 37, 8754–8763. [Google Scholar] [CrossRef] [PubMed]
- MacLean, M.J.; Ness, L.S.; Ferguson, G.P.; Booth, I.R. The role of glyoxalase I in the detoxification of methylglyoxal and in the activation of the KefB K+ efflux system in Escherichia coli. Mol. Microbiol. 1998, 27, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bao, W.; Guo, N.; Chen, H.; Cheng, W.; Jin, K.; Shen, F.; Xu, J.; Zhang, Q.; Wang, C.; et al. Antimicrobial activity of the imipenem/rifampicin combination against clinical isolates of Acinetobacter baumannii grown in planktonic and biofilm cultures. World J. Microbiol. Biotechnol. 2014, 30, 3015–3025. [Google Scholar] [CrossRef] [PubMed]
- Kohanski, M.A.; Dwyer, D.J.; Hayete, B.; Lawrence, C.A.; Collins, J.J. A common mechanism of cellular death induced by bactericidal antibiotics. Cell 2007, 130, 797–810. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Plasmid Replicon | Description |
|---|---|---|
| C600 | None | model E. coli strain |
| C600T | IncX3 | C600 transformant |
| C600N500 | IncX3 | 500th passage of C600T without antibiotics |
| C600Y500 | IncX3 | 500th passage of C600T with 2 mg/L imipenem |
| C600D500 | None | 500th passage of C600 |
| J53 | None | model E. coli strain |
| J53T | IncX3 | J53 transformant |
| Strain | MIC (mg/L) | ||||||
|---|---|---|---|---|---|---|---|
| MEM | IPM | FEP | CAZ | ATM | AMK | LEV | |
| C600 | 0.125 | 0.25 | 0.125 | 1 | 0.25 | 0.125 | 0.125 |
| C600T | 64 (512-fold) a | 64 (256-fold) a | 32 (256-fold) a | 128 (128-fold) a | 8 (32-fold) a | 0.125 | 0.125 |
| C600N500 | 32 | 32 | 32 | 128 | 8 | 0.125 | 0.125 |
| C600Y500 | 16 | 32 | 32 | 128 | 8 | 0.125 | 0.125 |
| J53 | 0.125 | 0.125 | 0.125 | 0.125 | 0.5 | 4 | 0.125 |
| J53T | 4 (32-fold) a | 16 (128-fold) a | 64 (512-fold) a | 64 (512-fold) a | 128 (512-fold) a | 4 | 0.125 |
| Strain | SNP Counts | ||
|---|---|---|---|
| Total SNPs | CDS Count | Intergenic Sequence Count | |
| J53T | 8 | 0 | 8 |
| C600T | 29 | 11 | 18 |
| Strain | Non-Synonymous SNP Gene |
|---|---|
| J53T | None |
| C600T | ptrA, lpdA, bamA, melA, gatA, mdoB, yihV, mntH, trkA, and ydfK |
| Strain | SNP Counts | ||
|---|---|---|---|
| Total SNPs | CDS Count | Intergenic Sequence Count | |
| C600N500 | 28 | 17 | 11 |
| C600Y500 | 27 | 11 | 16 |
| Strain | Non-Synonymous SNP Gene |
|---|---|
| C600N500 | gudD, araC, dgt, bamA, rfaF, lptF, ycjX, bcsG, pepQ, mshA, sfmD, paaJ, mreC, and tfaE |
| C600Y500 | gloA, ydiF, fliY, hycF, caiA, bamA, rlmB, sbp, acnA, and yghQ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Wang, J.; Jiang, Y.; Guo, Y.; Liu, N.; Xiao, S.; Yao, L.; Li, J.; Zhuo, C.; He, N.; et al. Adaptive Evolution Compensated for the Plasmid Fitness Costs Brought by Specific Genetic Conflicts. Pathogens 2023, 12, 137. https://doi.org/10.3390/pathogens12010137
Li F, Wang J, Jiang Y, Guo Y, Liu N, Xiao S, Yao L, Li J, Zhuo C, He N, et al. Adaptive Evolution Compensated for the Plasmid Fitness Costs Brought by Specific Genetic Conflicts. Pathogens. 2023; 12(1):137. https://doi.org/10.3390/pathogens12010137
Chicago/Turabian StyleLi, Feifeng, Jiong Wang, Ying Jiang, Yingyi Guo, Ningjing Liu, Shunian Xiao, Likang Yao, Jiahui Li, Chuyue Zhuo, Nanhao He, and et al. 2023. "Adaptive Evolution Compensated for the Plasmid Fitness Costs Brought by Specific Genetic Conflicts" Pathogens 12, no. 1: 137. https://doi.org/10.3390/pathogens12010137