
Polymyxin B in Combination with Glycerol Monolaurate Exerts Synergistic Killing against Gram-Negative Pathogens

 and
and
Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Susceptibility of PMNB and GML
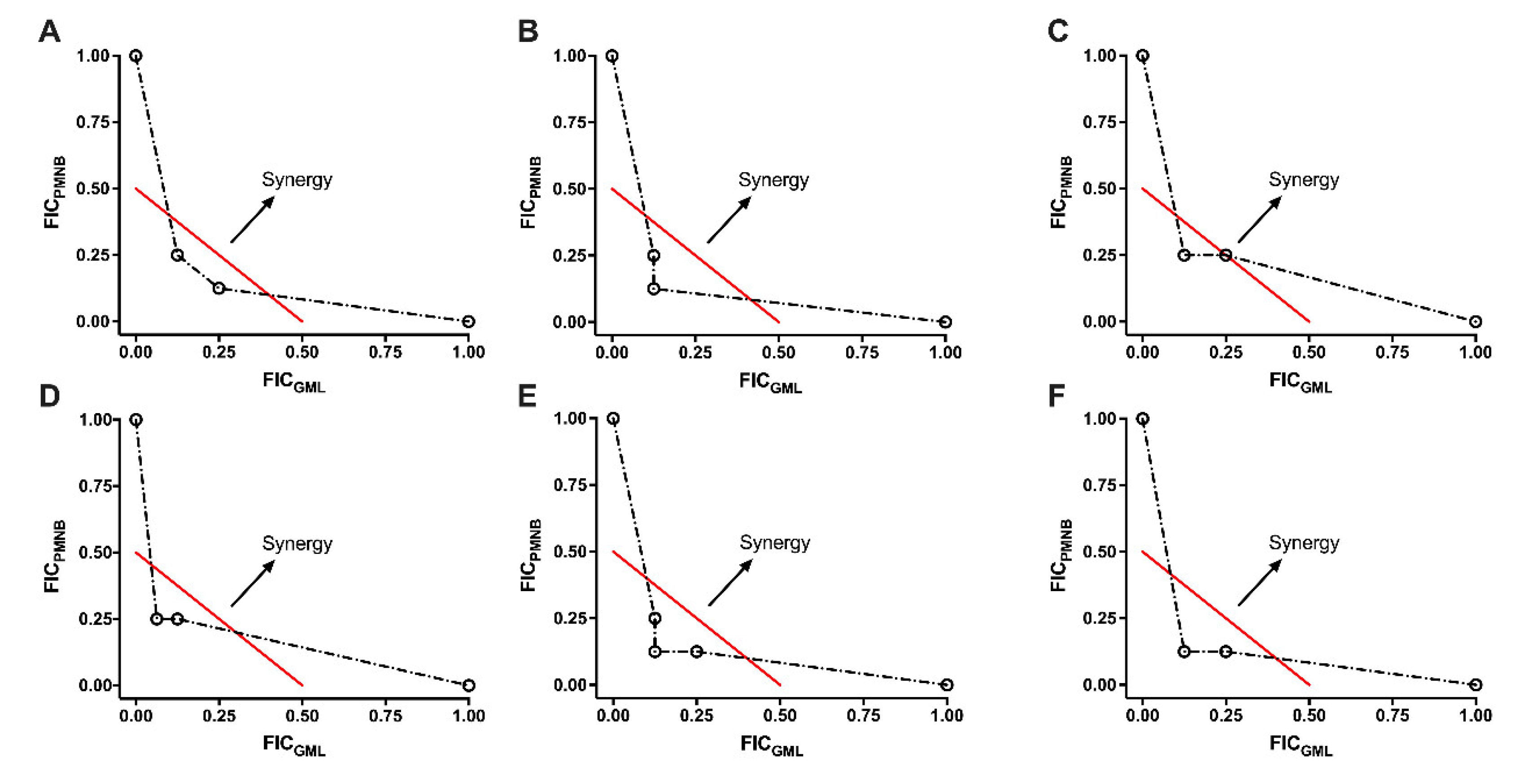
2.2. Synergistic Effect of PMNB Alongside GML against Gram-Negative Pathogens
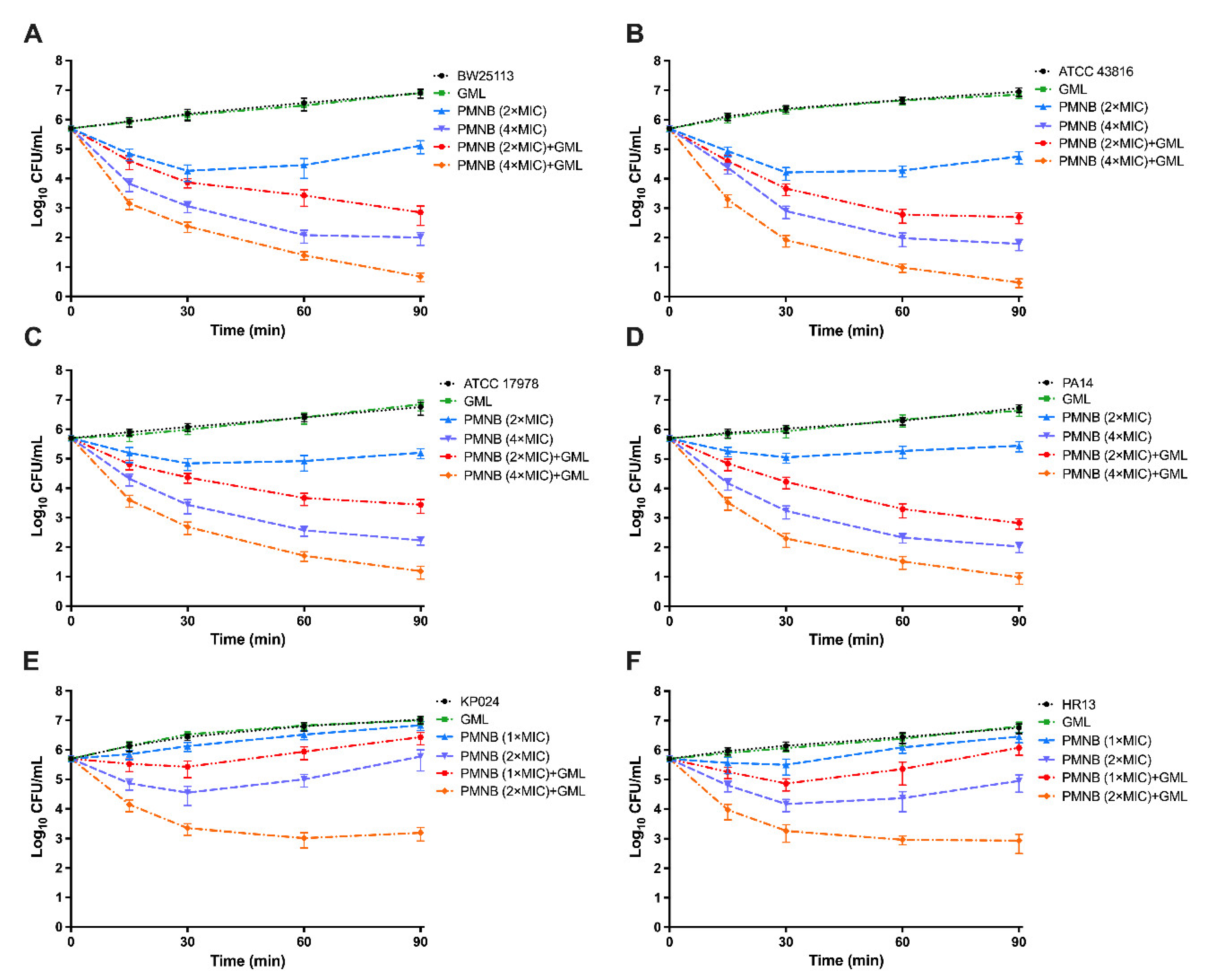
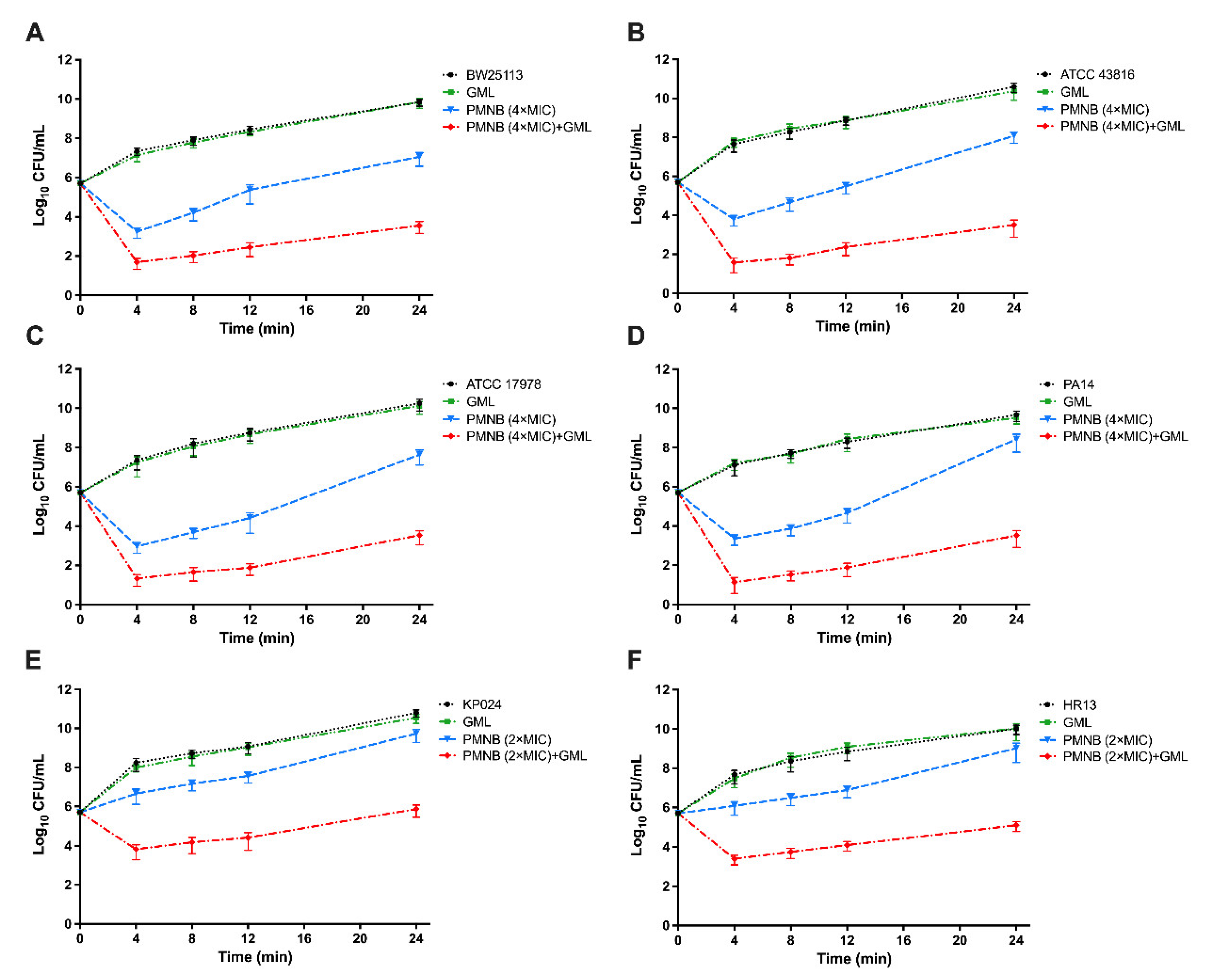
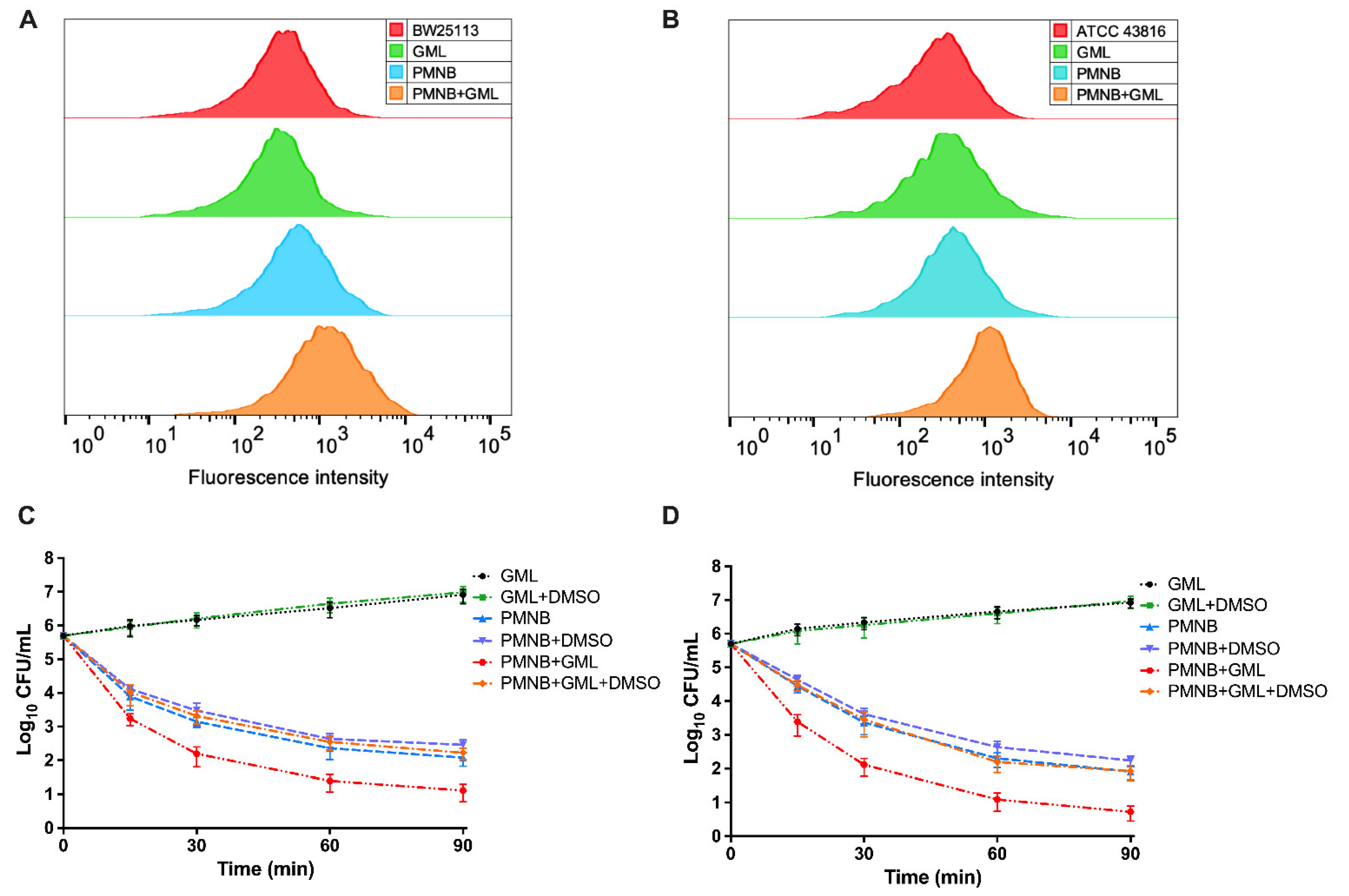
2.3. Antimicrobial Killing of PMNB in Combination with GML
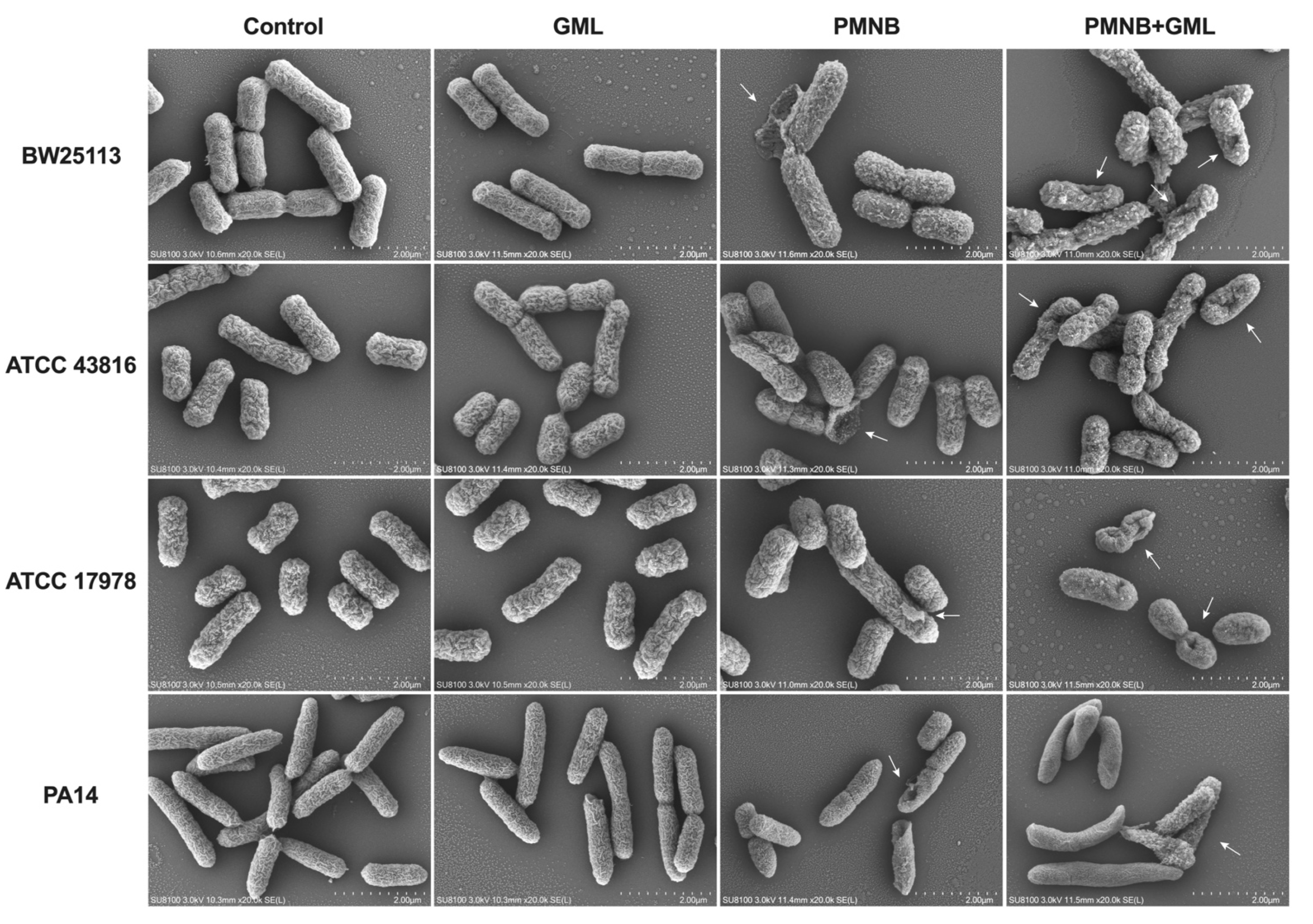
2.4. Combined PMNB and GML Treatment Aggravates Morphological Damage
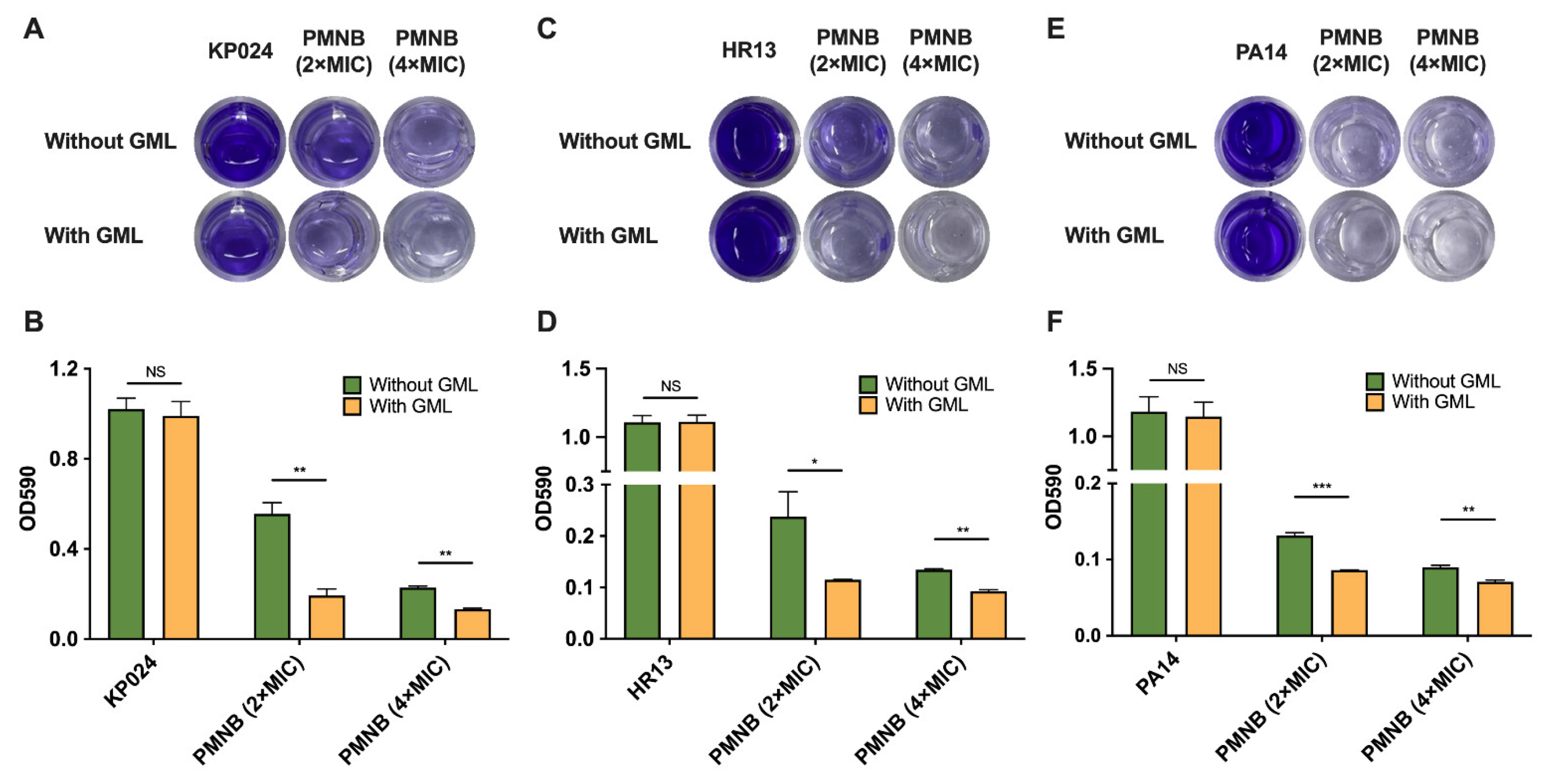
2.5. PMNB Synergized with GML against Biofilm Formation
2.6. PMNB–GML Combination Stimulates ROS Production
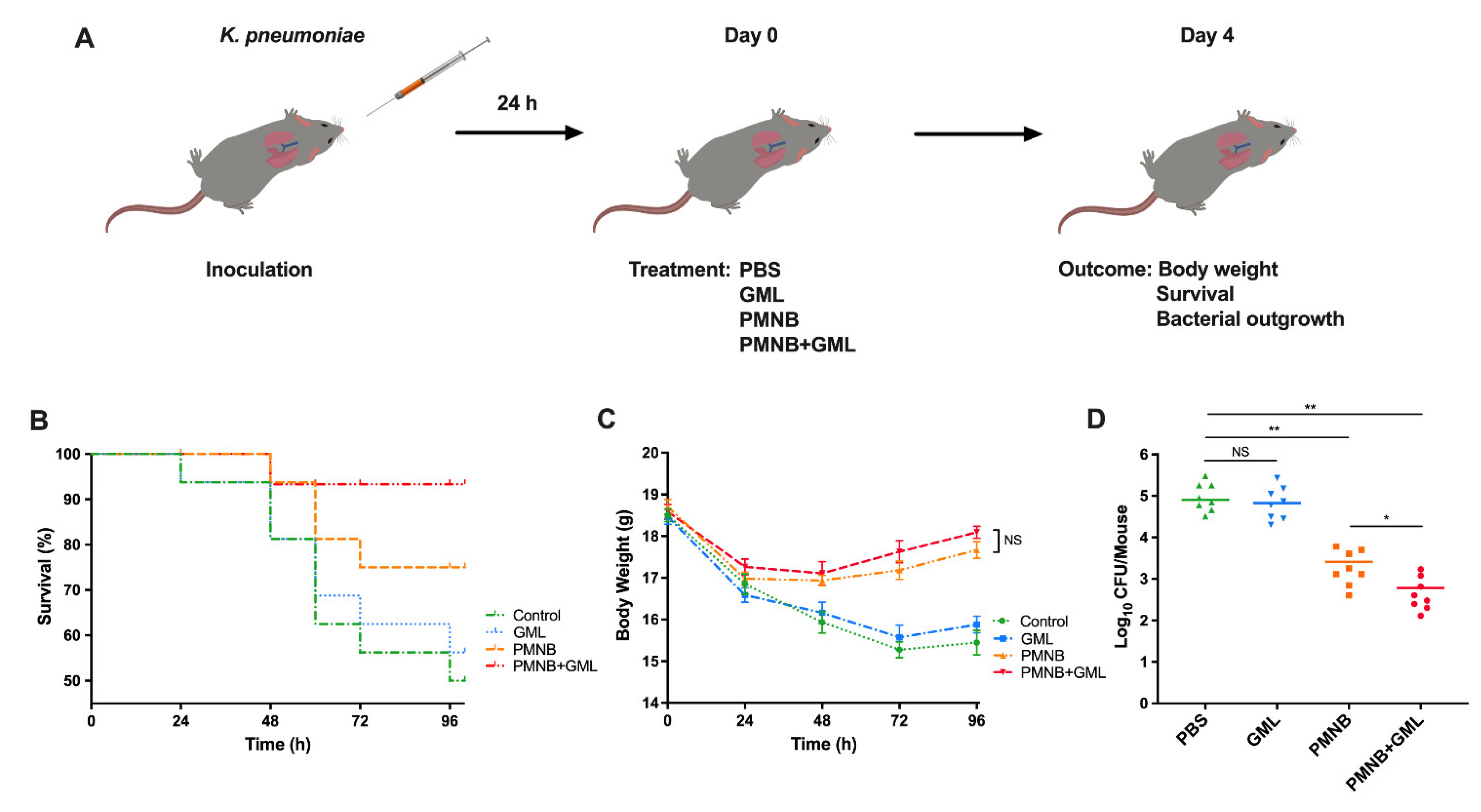
2.7. PMNB–GML Combination Contributed to Clearance of K. pneumoniae from Lung Infection
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates, Drugs, and Mice
4.2. Antimicrobial Susceptibility Testing
4.3. Checkerboard Assays
4.4. Time–Kill Studies
4.5. Cellular Morphology Study
4.6. Crystal Violet Biofilm Assay
4.7. ROS Assay
4.8. Murine Lung Infection Model
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Telhig, S.; Ben Said, L.; Zirah, S.; Fliss, I.; Rebuffat, S. Bacteriocins to Thwart Bacterial Resistance in Gram Negative Bacteria. Front. Microbiol. 2020, 11, 586433. [Google Scholar] [CrossRef] [PubMed]
- Willemse-Erix, D.; Bakker-Schut, T.; Slagboom-Bax, F.; Jachtenberg, J.W.; Lemmens-den Toom, N.; Papagiannitsis, C.C.; Kuntaman, K.; Puppels, G.; van Belkum, A.; Severin, J.A.; et al. Rapid typing of extended-spectrum β-lactamase- and carbapenemase-producing Escherichia coli and Klebsiella pneumoniae isolates by use of SpectraCell RA. J. Clin. Microbiol. 2012, 50, 1370–1375. [Google Scholar] [CrossRef] [PubMed]
- Elliott, A.G.; Huang, J.X.; Neve, S.; Zuegg, J.; Edwards, I.A.; Cain, A.K.; Boinett, C.J.; Barquist, L.; Lundberg, C.V.; Steen, J.; et al. An amphipathic peptide with antibiotic activity against multidrug-resistant Gram-negative bacteria. Nat. Commun. 2020, 11, 3184. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed Ahmed, M.; Zhong, L.L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.B. Colistin and its role in the Era of antibiotic resistance: An extended review (2000–2019). Emerg. Microbes. Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef]
- Fu, Y.Y.; Zhang, L.; Yang, Y.; Liu, C.W.; He, Y.N.; Li, P.; Yu, X. Synergistic antibacterial effect of ultrasound microbubbles combined with chitosan-modified polymyxin B-loaded liposomes on biofilm-producing Acinetobacter baumannii. Int. J. Nanomed. 2019, 14, 1805–1815. [Google Scholar] [CrossRef]
- Nang, S.C.; Azad, M.; Velkov, T.; Zhou, Q.T.; Li, J. Rescuing the Last-Line Polymyxins: Achievements and Challenges. Pharmacol. Rev. 2021, 73, 679–728. [Google Scholar] [CrossRef]
- Feng, Y. Transferability of MCR-1/2 Polymyxin Resistance: Complex Dissemination and Genetic Mechanism. ACS Infect. Dis. 2018, 4, 291–300. [Google Scholar] [CrossRef]
- Zhang, H.; Srinivas, S.; Xu, Y.; Wei, W.; Feng, Y. Genetic and Biochemical Mechanisms for Bacterial Lipid A Modifiers Associated with Polymyxin Resistance. Trends Biochem. Sci. 2019, 44, 973–988. [Google Scholar] [CrossRef]
- Dubashynskaya, N.V.; Raik, S.V.; Dubrovskii, Y.A.; Shcherbakova, E.S.; Demyanova, E.V.; Shasherina, A.Y.; Anufrikov, Y.A.; Poshina, D.N.; Dobrodumov, A.V.; Skorik, Y.A. Hyaluronan/colistin polyelectrolyte complexes: Promising antiinfective drug delivery systems. Int. J. Biol. Macromol. 2021, 187, 157–165. [Google Scholar] [CrossRef]
- Dubashynskaya, N.V.; Bokatyi, A.N.; Gasilova, E.R.; Dobrodumov, A.V.; Dubrovskii, Y.A.; Knyazeva, E.S.; Nashchekina, Y.A.; Demyanova, E.V.; Skorik, Y.A. Hyaluronan-colistin conjugates: Synthesis, characterization, and prospects for medical applications. Int. J. Biol. Macromol. 2022, 215, 243–252. [Google Scholar] [CrossRef]
- Dubashynskaya, N.V.; Raik, S.V.; Dubrovskii, Y.A.; Demyanova, E.V.; Shcherbakova, E.S.; Poshina, D.N.; Shasherina, A.Y.; Anufrikov, Y.A.; Skorik, Y.A. Hyaluronan/diethylaminoethyl chitosan polyelectrolyte complexes as carriers for improved colistin delivery. Int. J. Mol. Sci. 2021, 22, 8381. [Google Scholar] [CrossRef]
- Otto, R.G.; van Gorp, E.; Kloezen, W.; Meletiadis, J.; van den Berg, S.; Mouton, J.W. An alternative strategy for combination therapy: Interactions between polymyxin B and non-antibiotics. Int. J. Antimicrob. Agents 2019, 53, 34–39. [Google Scholar] [CrossRef]
- Ghattamaneni, N.; Panchal, S.K.; Brown, L. Nutraceuticals in rodent models as potential treatments for human Inflammatory Bowel Disease. Pharmacol. Res. 2018, 132, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Kabara, J.J.; Swieczkowski, D.M.; Conley, A.J.; Truant, J.P. Fatty acids and derivatives as antimicrobial agents. Antimicrob. Agents Chemother. 1972, 2, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Jiang, Z.; Cai, H.; Li, Y.; Mo, Q.; Deng, L.; Zhong, H.; Liu, T.; Zhang, H.; Kang, J.X.; et al. Modulation of the Gut Microbiota during High-Dose Glycerol Monolaurate-Mediated Amelioration of Obesity in Mice Fed a High-Fat Diet. mBio 2020, 11, e00190-20. [Google Scholar] [CrossRef]
- Hess, D.J.; Henry-Stanley, M.J.; Wells, C.L. Antibacterial synergy of glycerol monolaurate and aminoglycosides in Staphylococcus aureus biofilms. Antimicrob. Agents Chemother. 2014, 58, 6970–6973. [Google Scholar] [CrossRef]
- Schlievert, P.M.; Kilgore, S.H.; Kaus, G.M.; Ho, T.D.; Ellermeier, C.D. Glycerol Monolaurate (GML) and a Nonaqueous Five-Percent GML Gel Kill Bacillus and Clostridium Spores. mSphere 2018, 3, e00597-18. [Google Scholar] [CrossRef]
- Zavascki, A.P.; Nation, R.L. Nephrotoxicity of Polymyxins: Is There Any Difference between Colistimethate and Polymyxin B? Antimicrob. Agents Chemother. 2017, 61, e02319-16. [Google Scholar] [CrossRef]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [PubMed]
- Kohanski, M.A.; Dwyer, D.J.; Hayete, B.; Lawrence, C.A.; Collins, J.J. A common mechanism of cellular death induced by bactericidal antibiotics. Cell 2007, 130, 797–810. [Google Scholar] [CrossRef]
- Van Acker, H.; Coenye, T. The Role of Reactive Oxygen Species in Antibiotic-Mediated Killing of Bacteria. Trends Microbiol. 2017, 25, 456–466. [Google Scholar] [CrossRef]
- GBD 2016 Lower Respiratory Infections Collaborators. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of lower respiratory infections in 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1191–1210. [Google Scholar] [CrossRef]
- Wenzler, E.; Fraidenburg, D.R.; Scardina, T.; Danziger, L.H. Inhaled Antibiotics for Gram-Negative Respiratory Infections. Clin. Microbiol. Rev. 2016, 29, 581–632. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo-Troyano, A.; Sibila, O. The respiratory threat posed by multidrug resistant Gram-negative bacteria. Respirology 2017, 22, 1288–1299. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Liu, Y.; Huang, X.; Ding, S.; Wang, Y.; Shen, J.; Zhu, K. A broad-spectrum antibiotic adjuvant reverses multidrug-resistant Gram-negative pathogens. Nat. Microbiol. 2020, 5, 1040–1050. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, G.; Yang, L.; Wu, H.; Song, Z.; Wang, H.; Høiby, N.; Ulrich, M.; Molin, S.; Riethmüller, J.; Döring, G. Colistin-tobramycin combinations are superior to monotherapy concerning the killing of biofilm Pseudomonas aeruginosa. J. Infect. Dis. 2010, 202, 1585–1592. [Google Scholar] [CrossRef] [PubMed]
- Aye, S.M.; Galani, I.; Yu, H.; Wang, J.; Chen, K.; Wickremasinghe, H.; Karaiskos, I.; Bergen, P.J.; Zhao, J.; Velkov, T.; et al. Polymyxin Triple Combinations against Polymyxin-Resistant, Multidrug-Resistant, KPC-Producing Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2020, 64, e00246-20. [Google Scholar] [CrossRef]
- Welch, J.L.; Xiang, J.; Okeoma, C.M.; Schlievert, P.M.; Stapleton, J.T. Glycerol Monolaurate, an Analogue to a Factor Secreted by Lactobacillus, Is Virucidal against Enveloped Viruses, Including HIV-1. mBio 2020, 11, e00686-20. [Google Scholar] [CrossRef]
- Thompson, J.M.; Miller, R.J.; Ashbaugh, A.G.; Dillen, C.A.; Pickett, J.E.; Wang, Y.; Ortines, R.V.; Sterling, R.S.; Francis, K.P.; Bernthal, N.M.; et al. Mouse model of Gram-negative prosthetic joint infection reveals therapeutic targets. JCI Insight 2018, 3, e121737. [Google Scholar] [CrossRef]
- Yin, Q.; Wu, S.; Wu, L.; Wang, Z.; Mu, Y.; Zhang, R.; Dong, C.; Zhou, B.; Zhao, B.; Zheng, J.; et al. A novel in silico antimicrobial peptide DP7 combats MDR Pseudomonas aeruginosa and related biofilm infections. J. Antimicrob. Chemother. 2020, 75, 3248–3259. [Google Scholar] [CrossRef]
- Kim, B.O.; Jang, H.J.; Chung, I.Y.; Bae, H.W.; Kim, E.S.; Cho, Y.H. Nitrate Respiration Promotes Polymyxin B Resistance in Pseudomonas aeruginosa. Antioxid. Redox. Signal. 2021, 34, 442–451. [Google Scholar] [CrossRef]
- Keren, I.; Wu, Y.; Inocencio, J.; Mulcahy, L.R.; Lewis, K. Killing by bactericidal antibiotics does not depend on reactive oxygen species. Science 2013, 339, 1213–1216. [Google Scholar] [CrossRef]
- Cilloniz, C.; Martin-Loeches, I.; Garcia-Vidal, C.; San Jose, A.; Torres, A. Microbial Etiology of Pneumonia: Epidemiology, Diagnosis and Resistance Patterns. Int. J. Mol. Sci. 2016, 17, 2120. [Google Scholar] [CrossRef]
- Kumar, V. Pulmonary Innate Immune Response Determines the Outcome of Inflammation During Pneumonia and Sepsis-Associated Acute Lung Injury. Front. Immunol. 2020, 11, 1722. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: The Keio collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef] [PubMed]
- Rahme, L.G.; Stevens, E.J.; Wolfort, S.F.; Shao, J.; Tompkins, R.G.; Ausubel, F.M. Common virulence factors for bacterial pathogenicity in plants and animals. Science 1995, 268, 1899–1902. [Google Scholar] [CrossRef] [PubMed]
- Bakker-Woudenberg, I.A.; van den Berg, J.C.; Vree, T.B.; Baars, A.M.; Michel, M.F. Relevance of serum protein binding of cefoxitin and cefazolin to their activities against Klebsiella pneumoniae pneumonia in rats. Antimicrob. Agents Chemother. 1985, 28, 654–659. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 32nd ed.; CLSI supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022. [Google Scholar]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 12.0, 2022. Available online: http://www.eucast.org (accessed on 1 January 2022).
- Tran, T.B.; Bergen, P.J.; Creek, D.J.; Velkov, T.; Li, J. Novel polymyxin combination with antineoplastic mitotane improved the bacterial killing against polymyxin-resistant multidrug-resistant Gram-negative pathogens. Front. Microbiol. 2018, 9, 721. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, Q.; Wang, R.; Wang, H.; Wong, Y.T.; Wang, M.; Hao, Q.; Yan, A.; Kao, R.Y.; Ho, P.L.; et al. Resensitizing carbapenem- and colistin-resistant bacteria to antibiotics using auranofin. Nat. Commun. 2020, 11, 5263. [Google Scholar] [CrossRef]
- Chen, H.; Yang, N.; Yang, Y.; Zheng, Y.; Xu, M.; Zhang, H.; Liu, Y.; Shen, W.; Li, J. Doxofylline Protects Gram-Negative Pathogens against Antibiotic-Mediated Killing. ACS Infect. Dis. 2021, 7, 3241–3253. [Google Scholar] [CrossRef]
- Xiao, Y.; Xu, M.; Lv, N.; Cheng, C.; Huang, P.; Li, J.; Hu, Y.; Sun, M. Dual stimuli-responsive metal-organic framework-based nanosystem for synergistic photothermal/pharmacological antibacterial therapy. Acta Biomater. 2021, 122, 291–305. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.L.; Sequeira, R.P.; Clarke, T.B. The microbiota protects against respiratory infection via GM-CSF signaling. Nat. Commun. 2017, 8, 1512. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.J.; Jang, A.R.; Park, J.Y.; Ahn, J.H.; Lee, T.S.; Kim, D.Y.; Lee, M.S.; Hwang, S.; Jeong, Y.J.; Park, J.H. IL-10 Protects Mice from the Lung Infection of Acinetobacter baumannii and Contributes to Bacterial Clearance by Regulating STAT3-Mediated MARCO Expression in Macrophages. Front. Immunol. 2020, 11, 270. [Google Scholar] [CrossRef] [PubMed]
- Gaurav, A.; Kothari, A.; Omar, B.J.; Pathania, R. Assessment of polymyxin B–doxycycline in combination against Pseudomonas aeruginosa in vitro and in a mouse model of acute pneumonia. Int. J. Antimicrob. Agents 2020, 56, 106022. [Google Scholar] [CrossRef] [PubMed]
- Mo, Q.; Liu, T.; Fu, A.; Ruan, S.; Zhong, H.; Tang, J.; Zhao, M.; Li, Y.; Zhu, S.; Cai, H.; et al. Novel Gut Microbiota Patterns Involved in the Attenuation of Dextran Sodium Sulfate-Induced Mouse Colitis Mediated by Glycerol Monolaurate via Inducing Anti-inflammatory Responses. mBio 2021, 12, e0214821. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Isolate | Agent | MIC (μg/mL) | MICIn combination/MICsingly | FICI | Outcome | |
|---|---|---|---|---|---|---|
| Singly | In Combination | |||||
| E. coli BW25113 | PMNB | 0.5 | 0.125 | 0.25 | <0.375 | Synergy |
| GML | >4000 | 500 | <0.125 | |||
| K. pneumoniae ATCC 43816 | PMNB | 1 | 0.125 | 0.125 | <0.25 | Synergy |
| GML | >4000 | 500 | <0.125 | |||
| K. pneumoniae KP024 | PMNB | 8 | 1 | 0.125 | <0.25 | Synergy |
| GML | >4000 | 500 | <0.125 | |||
| A. baumannii ATCC 17978 | PMNB | 0.5 | 0.125 | 0.25 | <0.375 | Synergy |
| GML | >4000 | 500 | <0.125 | |||
| A. baumannii HR13 | PMNB | 4 | 0.5 | 0.125 | <0.25 | Synergy |
| GML | >4000 | 500 | <0.125 | |||
| P. aeruginosa PA14 | PMNB | 0.5 | 0.125 | 0.25 | <0.3125 | Synergy |
| GML | >4000 | 250 | <0.0625 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Yang, N.; Ding, Y.; Li, J.; Liu, Y.; Chen, H.; Li, J. Polymyxin B in Combination with Glycerol Monolaurate Exerts Synergistic Killing against Gram-Negative Pathogens. Pathogens 2022, 11, 874. https://doi.org/10.3390/pathogens11080874
Zheng Y, Yang N, Ding Y, Li J, Liu Y, Chen H, Li J. Polymyxin B in Combination with Glycerol Monolaurate Exerts Synergistic Killing against Gram-Negative Pathogens. Pathogens. 2022; 11(8):874. https://doi.org/10.3390/pathogens11080874
Chicago/Turabian StyleZheng, Yun, Ning Yang, Yuting Ding, Jiajia Li, Yanyan Liu, Haoran Chen, and Jiabin Li. 2022. "Polymyxin B in Combination with Glycerol Monolaurate Exerts Synergistic Killing against Gram-Negative Pathogens" Pathogens 11, no. 8: 874. https://doi.org/10.3390/pathogens11080874