
Exploring the Biofilm Formation Capacity in S. pseudintermedius and Coagulase-Negative Staphylococci Species

 ,
,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Design and Bacterial Isolates
2.2. Biofilm Formation Assay
Biofilm Biomass Quantification
2.3. Confocal Laser Scanning Microscopy (CLSM)
2.4. Effect of Antibiotics on 24 h-Old Biofilms
Effect of Antibiotics on Metabolic Activity
2.5. Statistical Analysis
3. Results
3.1. Biofilm Formation
3.2. Antimicrobial Resistance and Biofilm Formation
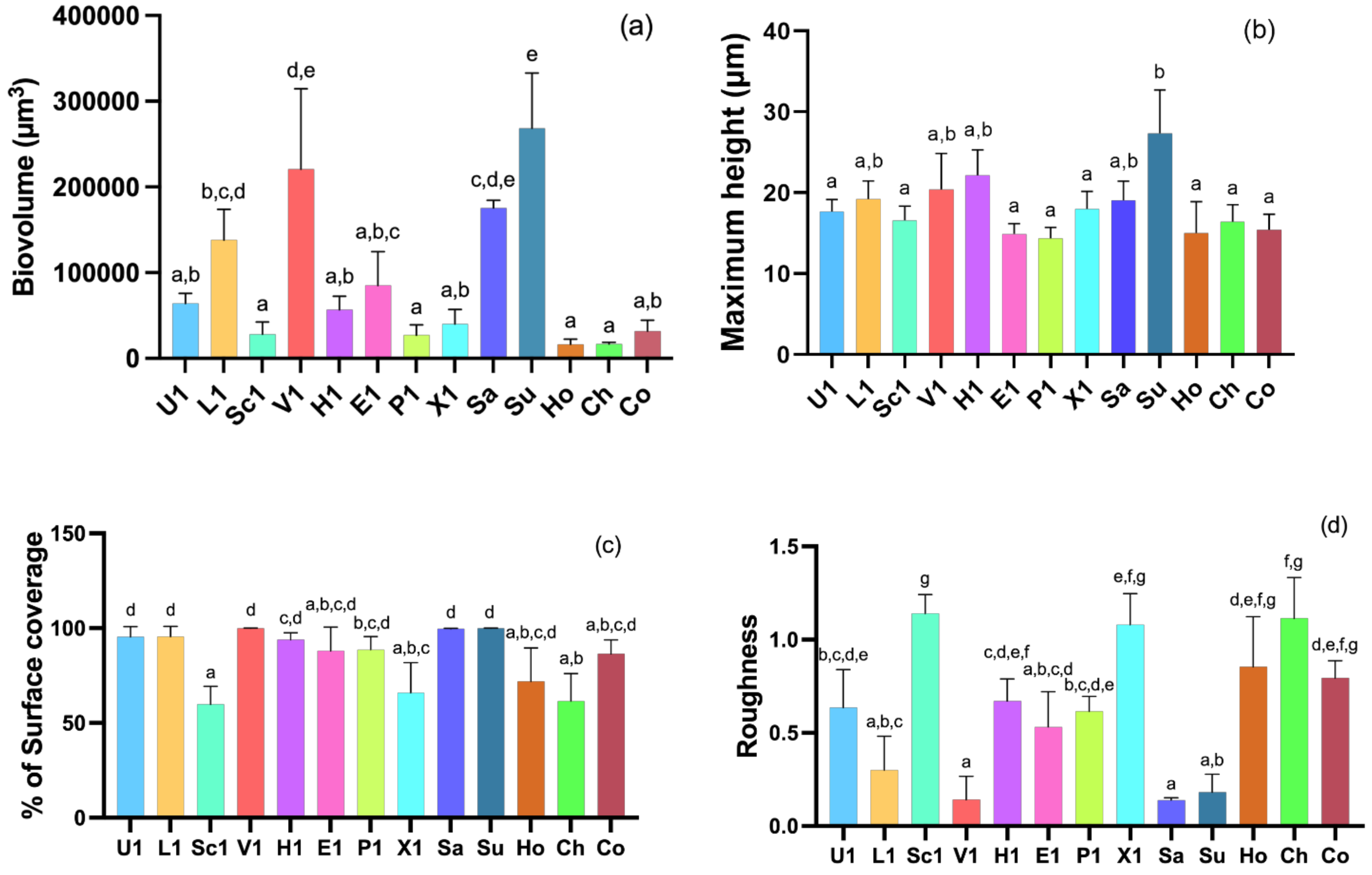
3.3. CLSM Analysis
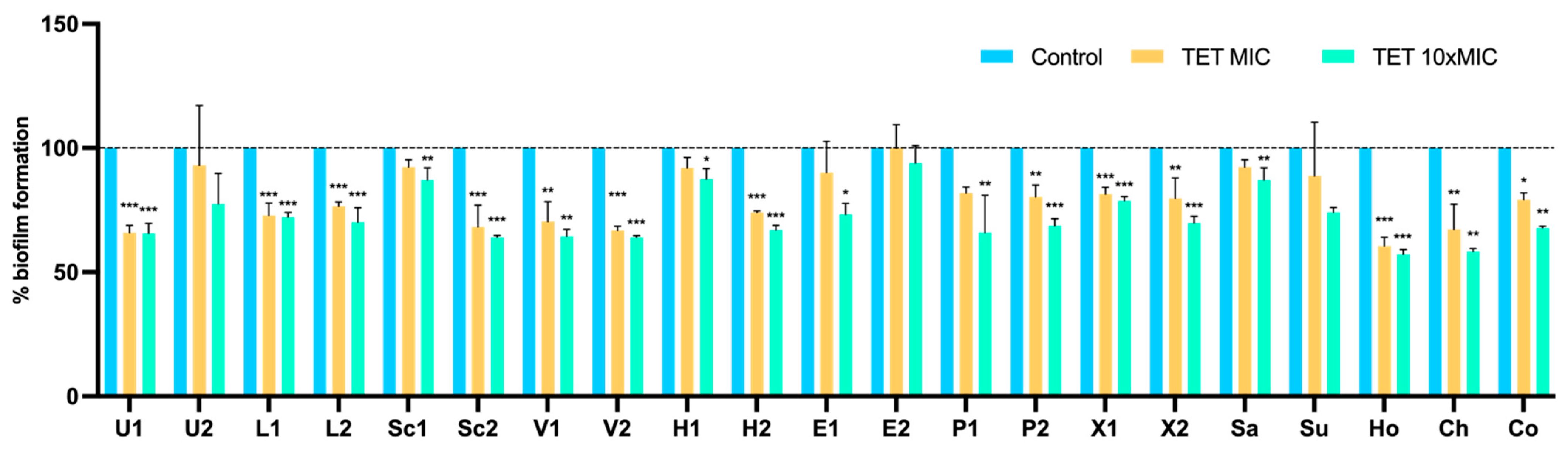
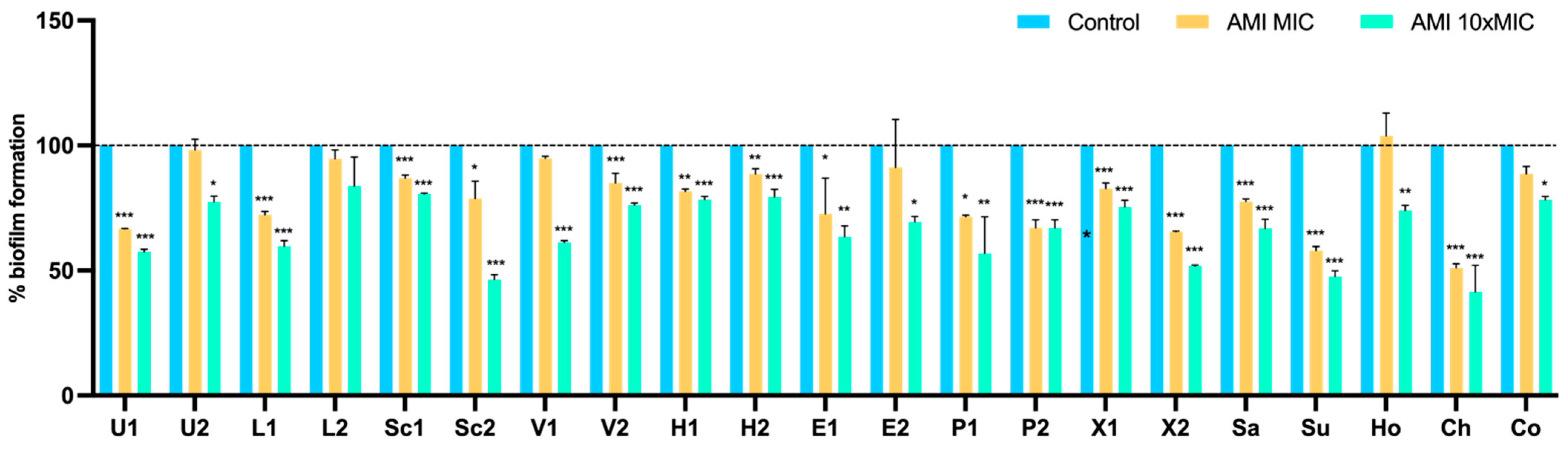
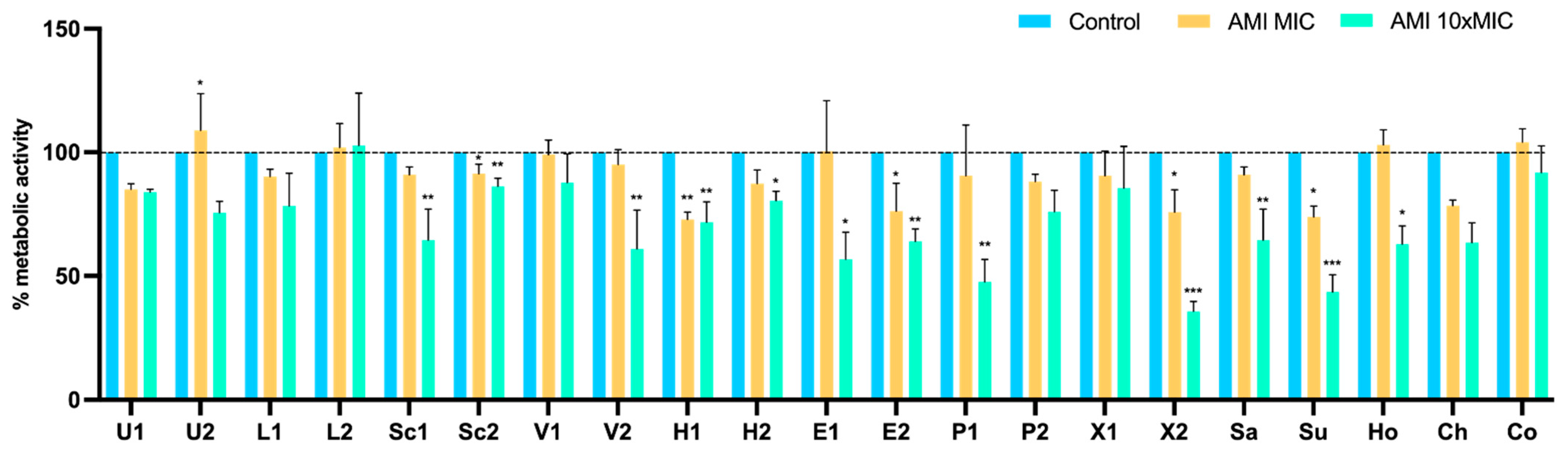
3.4. Effect of Antimicrobials on 24 h-Old Biofilms
Metabolic Activity
4. Discussion
4.1. Biofilm Formation
4.2. Antimicrobial Resistance and Biofilm Formation
4.3. Effect of Antimicrobials on 24 h-Old Biofilms
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heilbronner, S. Staphylococcus lugdunensis. Trends Microbiol. 2021, 29, 1143–1145. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-negative staphylococci. Clin. Microbiol. Rev. 2014, 27, 870–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Moein, K.A.; Zaher, H.M. The Nasal Carriage of Coagulase-Negative Staphylococci Among Animals and Its Public Health Implication. Vector Borne Zoonotic Dis. 2020, 20, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Kloos, W.E.; Bannerman, T.L. Update on clinical significance of coagulase-negative staphylococci. Clin. Microbiol. Rev. 1994, 7, 117–140. [Google Scholar] [CrossRef]
- Heilmann, C.; Ziebuhr, W.; Becker, K. Are coagulase-negative staphylococci virulent? Clin. Microbiol. Infect. 2019, 25, 1071–1080. [Google Scholar] [CrossRef]
- Michels, R.; Last, K.; Becker, S.L.; Papan, C. Update on Coagulase-Negative Staphylococci—What the Clinician Should Know. Microorg. 2021, 9, 830. [Google Scholar] [CrossRef]
- Saber, H.; Jasni, A.S.; Jamaluddin, T.Z.M.T.; Ibrahim, R. A Review of Staphylococcal Cassette Chromosome mec (SCCmec) Types in Coagulase-Negative Staphylococci (CoNS) Species. Malays. J. Med. Sci. 2017, 24, 7–18. [Google Scholar] [CrossRef]
- Asante, J.; Hetsa, B.A.; Amoako, D.G.; Abia, A.L.K.; Bester, L.A.; Essack, S.Y. Multidrug-resistant coagulase-negative staphylococci isolated from bloodstream in the uMgungundlovu district of KwaZulu-Natal Province in South Africa: Emerging pathogens. Antibiotics 2021, 10, 198. [Google Scholar] [CrossRef]
- Chon, J.-W.; Lee, U.J.; Bensen, R.; West, S.; Paredes, A.; Lim, J.; Khan, S.; Hart, M.E.; Phillips, K.S.; Sung, K. Virulence Characteristics of mecA-Positive Multidrug-Resistant Clinical Coagulase-Negative Staphylococci. Microorg. 2020, 8, 659. [Google Scholar] [CrossRef]
- Michalik, M.; Nowakiewicz, A.; Trościańczyk, A.; Kowalski, C.; Podbielska-Kubera, A. Multidrug resistant coagulase-negative Staphylococcus spp. isolated from cases of chronic rhinosinusitis in humans. Study from Poland. Acta Microbiol. Immunol. Hung. 2022, 69, 68–76. [Google Scholar] [CrossRef]
- Silva, V.; Caniça, M.; Ferreira, E.; Vieira-Pinto, M.; Saraiva, C.; Pereira, J.E.; Capelo, J.L.; Igrejas, G.; Poeta, P. Multidrug-Resistant Methicillin-Resistant Coagulase-Negative Staphylococci in Healthy Poultry Slaughtered for Human Consumption. Antibiot. 2022, 11, 365. [Google Scholar] [CrossRef]
- Kizerwetter-Świda, M.; Chrobak-Chmiel, D.; Rzewuska, M. High-level mupirocin resistance in methicillin-resistant staphylococci isolated from dogs and cats. BMC Vet. Res. 2019, 15, 238. [Google Scholar] [CrossRef]
- Pexara, A.; Solomakos, N.; Govaris, A. Occurrence, antibiotic resistance and enteroxigenicity of Staphylococcus spp. in tonsils of slaughtered pigs in Greece. Lett. Appl. Microbiol. 2020, 71, 394–399. [Google Scholar] [CrossRef]
- Marincola, G.; Liong, O.; Schoen, C.; Abouelfetouh, A.; Hamdy, A.; Wencker, F.D.R.; Marciniak, T.; Becker, K.; Köck, R.; Ziebuhr, W. Antimicrobial resistance profiles of coagulase-negative staphylococci in community-based healthy individuals in Germany. Front. Public Heal. 2021, 9, 796. [Google Scholar] [CrossRef]
- Schilcher, K.; Horswill, A.R. Staphylococcal Biofilm Development: Structure, Regulation, and Treatment Strategies. Microbiol. Mol. Biol. Rev. 2020, 84, e00026-19. [Google Scholar] [CrossRef]
- Argemi, X.; Hansmann, Y.; Prola, K.; Prévost, G. Coagulase-negative staphylococci pathogenomics. Int. J. Mol. Sci. 2019, 20, 1215. [Google Scholar] [CrossRef] [Green Version]
- Silva, V.; Capelo, J.L.; Igrejas, G.; Poeta, P. Molecular Mechanisms of Antimicrobial Resistance in Staphylococcus aureus Biofilms BT—Emerging Modalities in Mitigation of Antimicrobial Resistance. In; Akhtar, N., Singh, K.S., Prerna, G.D., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 291–314. ISBN 978-3-030-84126-3. [Google Scholar]
- Geoghegan, J.A.; Foster, T.J. Cell wall-anchored surface proteins of Staphylococcus aureus: Many proteins, multiple functions. In Staphylococcus Aureus; Springer: Cham, Switzerland, 2015; pp. 95–120. [Google Scholar]
- Wojtyczka, R.D.; Orlewska, K.; Kępa, M.; Idzik, D.; Dziedzic, A.; Mularz, T.; Krawczyk, M.; Miklasińska, M.; Wąsik, T.J. Biofilm formation and antimicrobial susceptibility of Staphylococcus epidermidis strains from a hospital environment. Int. J. Environ. Res. Public Health 2014, 11, 4619–4633. [Google Scholar] [CrossRef] [Green Version]
- Folliero, V.; Franci, G.; Dell’Annunziata, F.; Giugliano, R.; Foglia, F.; Sperlongano, R.; De Filippis, A.; Finamore, E.; Galdiero, M. Evaluation of Antibiotic Resistance and Biofilm Production among Clinical Strain Isolated from Medical Devices. Int. J. Microbiol. 2021, 2021, 9033278. [Google Scholar] [CrossRef]
- Govan, J.R.; Deretic, V. Microbial pathogenesis in cystic fibrosis: Mucoid Pseudomonas aeruginosa and Burkholderia cepacia. Microbiol. Rev. 1996, 60, 539–574. [Google Scholar] [CrossRef]
- Singh, S.; Singh, S.K.; Chowdhury, I.; Singh, R. Understanding the Mechanism of Bacterial Biofilms Resistance to Antimicrobial Agents. Open Microbiol. J. 2017, 11, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Silva, V.; Oliveira, A.; Manageiro, V.; Caniça, M.; Contente, D.; Capita, R.; Alonso-Calleja, C.; Carvalho, I.; Capelo, J.L.; Igrejas, G.; et al. Clonal Diversity and Antimicrobial Resistance of Methicillin-Resistant Staphylococcus pseudintermedius Isolated from Canine Pyoderma. Microorg. 2021, 9, 482. [Google Scholar] [CrossRef]
- Silva, V.; Lopes, A.F.; Soeiro, V.; Caniça, M.; Manageiro, V.; Pereira, J.E.; Maltez, L.; Capelo, J.L.; Igrejas, G.; Poeta, P. Nocturnal Birds of Prey as Carriers of Staphylococcus aureus and Other Staphylococci: Diversity, Antimicrobial Resistance and Clonal Lineages. Antibiot. 2022, 11, 240. [Google Scholar] [CrossRef]
- Silva, V.; Alfarela, C.; Caniça, M.; Manageiro, V.; Nóvoa, M.; Leiva, B.; Kress, M.; Capelo, J.L.; Poeta, P.; Igrejas, G. A One Health Approach Molecular Analysis of Staphylococcus aureus Reveals Distinct Lineages in Isolates from Miranda Donkeys (Equus asinus) and Their Handlers. Antibiot. 2022, 11, 374. [Google Scholar] [CrossRef]
- Silva, V.; Pereira, J.E.; Maltez, L.; Ferreira, E.; Manageiro, V.; Caniça, M.; Capelo, J.L.; Igrejas, G.; Poeta, P. Diversity of methicillin-resistant staphylococci among wild Lepus granatensis: First detection of mecA-MRSA in hares. FEMS Microbiol. Ecol. 2020, 96, fiz204. [Google Scholar] [CrossRef]
- Santos, V.; Gomes, A.; Ruiz-Ripa, L.; Mama, O.M.; Sabença, C.; Sousa, M.; Silva, V.; Sousa, T.; Vieira-Pinto, M.; Igrejas, G.; et al. Methicillin-Resistant Staphylococcus aureus CC398 in Purulent Lesions of Piglets and Fattening Pigs in Portugal. Microb. Drug Resist. 2020, 26, 850–856. [Google Scholar] [CrossRef]
- Silva, V.; de Sousa, T.; Gómez, P.; Sabença, C.; Vieira-Pinto, M.; Capita, R.; Alonso-Calleja, C.; Torres, C.; Capelo, J.L.; Igrejas, G.; et al. Livestock-Associated Methicillin-Resistant Staphylococcus aureus (MRSA) in Purulent Subcutaneous Lesions of Farm Rabbits. Foods 2020, 9, 439. [Google Scholar] [CrossRef] [Green Version]
- Silva, V.; Vieira-Pinto, M.; Saraiva, C.; Manageiro, V.; Reis, L.; Ferreira, E.; Caniça, M.; Capelo, J.L.; Igrejas, G.; Poeta, P. Prevalence and Characteristics of Multidrug-Resistant Livestock-Associated Methicillin-Resistant Staphylococcus aureus (LA-MRSA) CC398 Isolated from Quails (Coturnix Coturnix Japonica) Slaughtered for Human Consumption. Animals 2021, 11, 2038. [Google Scholar] [CrossRef]
- Silva, V.; Gabriel, S.I.; Borrego, S.B.; Tejedor-Junco, M.T.; Manageiro, V.; Ferreira, E.; Reis, L.; Caniça, M.; Capelo, J.L.; Igrejas, G.; et al. Antimicrobial Resistance and Genetic Lineages of Staphylococcus aureus from Wild Rodents: First Report of mecC-Positive Methicillin-Resistant S. aureus (MRSA) in Portugal. Animals 2021, 11, 1537. [Google Scholar] [CrossRef]
- Oniciuc, E.-A.; Cerca, N.; Nicolau, A.I. Compositional Analysis of Biofilms Formed by Staphylococcus aureus Isolated from Food Sources. Front. Microbiol. 2016, 7, 390. [Google Scholar] [CrossRef] [Green Version]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 2008, 72, 157–165. [Google Scholar] [CrossRef] [Green Version]
- González-Machado, C.; Capita, R.; Riesco-Peláez, F.; Alonso-Calleja, C. Visualization and quantification of the cellular and extracellular components of Salmonella Agona biofilms at different stages of development. PLoS ONE 2018, 13, e0200011. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Both, A.; Weißelberg, S.; Heilmann, C.; Rohde, H. Emergence of coagulase-negative staphylococci. Expert Rev. Anti. Infect. Ther. 2020, 18, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Morgenstern, M.; Post, V.; Erichsen, C.; Hungerer, S.; Bühren, V.; Militz, M.; Richards, R.G.; Moriarty, T.F. Biofilm formation increases treatment failure in Staphylococcus epidermidis device-related osteomyelitis of the lower extremity in human patients. J. Orthop. Res. 2016, 34, 1905–1913. [Google Scholar] [CrossRef] [PubMed]
- Bino, E.; Lauková, A.; Ščerbová, J.; Kubašová, I.; Kandričáková, A.; Strompfová, V.; Miltko, R.; Belzecki, G. Fecal coagulase-negative staphylococci from horses, their species variability, and biofilm formation. Folia Microbiol. 2019, 64, 719–726. [Google Scholar] [CrossRef]
- De Martino, L.; Lucido, M.; Mallardo, K.; Facello, B.; Mallardo, M.; Iovane, G.; Pagnini, U.; Tufano, M.A.; Catalanotti, P. Methicillin-Resistant Staphylococci Isolated from Healthy Horses and Horse Personnel in Italy. J. Vet. Diagnostic Investig. 2010, 22, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Ugwu, C.C.; Gomez-Sanz, E.; Agbo, I.C.; Torres, C.; Chah, K.F. Characterization of mannitol-fermenting methicillin-resistant staphylococci isolated from pigs in Nigeria. Braz. J. Microbiol. 2015, 46, 885–892. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Agidi, S.; LeJeune, J.T. Diversity of staphylococcal cassette chromosome in coagulase-negative staphylococci from animal sources. J. Appl. Microbiol. 2009, 107, 1375–1383. [Google Scholar] [CrossRef]
- Nemeghaire, S.; Vanderhaeghen, W.; Argudín, M.A.; Haesebrouck, F.; Butaye, P. Characterization of methicillin-resistant Staphylococcus sciuri isolates from industrially raised pigs, cattle and broiler chickens. J. Antimicrob. Chemother. 2014, 69, 2928–2934. [Google Scholar] [CrossRef] [Green Version]
- Kalai, S.; Roychoudhury, P.; Dutta, T.K.; Subudhi, P.K.; Chakraborty, S.; Barman, N.N.; Sen, A. Multidrug resistant staphylococci isolated from pigs with exudative epidermitis in North eastern Region of India. Lett. Appl. Microbiol. 2021, 72, 535–541. [Google Scholar] [CrossRef]
- Singh, A.; Walker, M.; Rousseau, J.; Weese, J.S. Characterization of the biofilm forming ability of Staphylococcus pseudintermedius from dogs. BMC Vet. Res. 2013, 9, 93. [Google Scholar] [CrossRef] [Green Version]
- Osland, A.M.; Vestby, L.K.; Fanuelsen, H.; Slettemeås, J.S.; Sunde, M. Clonal diversity and biofilm-forming ability of methicillin-resistant Staphylococcus pseudintermedius. J. Antimicrob. Chemother. 2012, 67, 841–848. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, L.; Brito, C.I.; de Oliveira, A.; Pereira, V.C.; da Cunha de Souza, M.d.L.R. Staphylococcus epidermidis and Staphylococcus haemolyticus: Detection of biofilm genes and biofilm formation in blood culture isolates from patients in a Brazilian teaching hospital. Diagn. Microbiol. Infect. Dis. 2016, 86, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, L.B.; Bhattarai, N.R.; Khanal, B. Comparative evaluation of methods for the detection of biofilm formation in coagulase-negative staphylococci and correlation with antibiogram. Infect. Drug Resist. 2018, 11, 607. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, Y.D.N.; Lamarche, D.; Chever, P.; Haine, D.; Messier, S.; Jacques, M. Characterization of the ability of coagulase-negative staphylococci isolated from the milk of Canadian farms to form biofilms. J. Dairy Sci. 2013, 96, 234–246. [Google Scholar] [CrossRef] [Green Version]
- Cabrera-Contreras, R.; Morelos-Ramírez, R.; Galicia-Camacho, A.N.; Meléndez-Herrada, E. Antibiotic resistance and biofilm production in Staphylococcus epidermidis strains, isolated from a tertiary care hospital in Mexico City. Int. Sch. Res. Not. 2013, 2013, 918921. [Google Scholar] [CrossRef] [Green Version]
- Singhai, M.; Malik, A.; Shahid, M.; Malik, M.A.; Goyal, R. A study on device-related infections with special reference to biofilm production and antibiotic resistance. J. Glob. Infect. Dis. 2012, 4, 193–198. [Google Scholar] [CrossRef]
- Farajzadeh Sheikh, A.; Asareh Zadegan Dezfuli, A.; Navidifar, T.; Fard, S.S.; Dehdashtian, M. Association between biofilm formation, structure and antibiotic resistance in Staphylococcus epidermidis isolated from neonatal septicemia in southwest Iran. Infect. Drug Resist. 2019, 12, 1771–1782. [Google Scholar] [CrossRef] [Green Version]
- Koksal, F.; Yasar, H.; Samasti, M. Antibiotic resistance patterns of coagulase-negative staphylococcus strains isolated from blood cultures of septicemic patients in Turkey. Microbiol. Res. 2009, 164, 404–410. [Google Scholar] [CrossRef]
- Jimi, S.; Miyazaki, M.; Takata, T.; Ohjimi, H.; Akita, S.; Hara, S. Increased drug resistance of meticillin-resistant Staphylococcus aureus biofilms formed on a mouse dermal chip model. J. Med. Microbiol. 2017, 66, 542. [Google Scholar] [CrossRef]
- Shah, M.S.; Qureshi, S.; Kashoo, Z.; Farooq, S.; Wani, S.A.; Hussain, M.I.; Banday, M.S.; Khan, A.A.; Gull, B.; Habib, A.; et al. Methicillin resistance genes and in vitro biofilm formation among Staphylococcus aureus isolates from bovine mastitis in India. Comp. Immunol. Microbiol. Infect. Dis. 2019, 64, 117–124. [Google Scholar] [CrossRef]
- Sun, X.; Lin, Z.; Hu, X.; Yao, W.; Bai, B.; Wang, H.; Li, D.; Chen, Z.; Cheng, H.; Pan, W.; et al. Biofilm formation in erythromycin-resistant Staphylococcus aureus and the relationship with antimicrobial susceptibility and molecular characteristics. Microb. Pathog. 2018, 124, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Francisco, M.S.; Rossi, C.C.; Brito, M.A.V.P.; Laport, M.S.; Barros, E.M.; Giambiagi-deMarval, M. Characterization of biofilms and antimicrobial resistance of coagulase-negative Staphylococcus species involved with subclinical mastitis. J. Dairy Res. 2021, 88, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Brady, A.J.; Laverty, G.; Gilpin, D.F.; Kearney, P.; Tunney, M. Antibiotic susceptibility of planktonic- and biofilm-grown staphylococci isolated from implant-associated infections: Should MBEC and nature of biofilm formation replace MIC? J. Med. Microbiol. 2017, 66, 461–469. [Google Scholar] [CrossRef] [Green Version]
- Flemming, K.; Klingenberg, C.; Cavanagh, J.P.; Sletteng, M.; Stensen, W.; Svendsen, J.S.; Flægstad, T. High in vitro antimicrobial activity of synthetic antimicrobial peptidomimetics against staphylococcal biofilms. J. Antimicrob. Chemother. 2009, 63, 136–145. [Google Scholar] [CrossRef]
- Mendoza-Olazarán, S.; Morfín-Otero, R.; Villarreal-Treviño, L.; Rodríguez-Noriega, E.; Llaca-Díaz, J.; Camacho-Ortiz, A.; González, G.M.; Casillas-Vega, N.; Garza-González, E. Antibiotic Susceptibility of Biofilm Cells and Molecular Characterisation of Staphylococcus hominis Isolates from Blood. PLoS ONE 2015, 10, e0144684. [Google Scholar] [CrossRef]
- Al-Ouqaili, M.T.S. Biofilm antimicrobial susceptibility pattern for selected antimicrobial agents against planktonic and sessile cells of clinical isolates of staphylococci using MICs, BICs and MBECs. Asian J Pharm 2018, 12. [Google Scholar]
- Qu, Y.; Daley, A.J.; Istivan, T.S.; Garland, S.M.; Deighton, M.A. Antibiotic susceptibility of coagulase-negative staphylococci isolated from very low birth weight babies: Comprehensive comparisons of bacteria at different stages of biofilm formation. Ann. Clin. Microbiol. Antimicrob. 2010, 9, 16. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, A.; Cataneli Pereira, V.; Pinheiro, L.; Moraes Riboli, D.F.; Benini Martins, K.; Ribeiro de Souza da Cunha, M.D. Antimicrobial Resistance Profile of Planktonic and Biofilm Cells of Staphylococcus aureus and Coagulase-Negative Staphylococci. Int. J. Mol. Sci. 2016, 17, 1423. [Google Scholar] [CrossRef] [Green Version]
- Walker, M.; Singh, A.; Nazarali, A.; Gibson, T.W.G.; Rousseau, J.; Weese, J.S. Evaluation of the impact of methicillin-resistant Staphylococcus pseudintermedius biofilm formation on antimicrobial susceptibility. Vet. Surg. 2016, 45, 968–971. [Google Scholar] [CrossRef]
- Glinka, M.; Wojnowski, W.; Wasik, A. Determination of aminoglycoside antibiotics: Current status and future trends. TrAC Trends Anal. Chem. 2020, 131, 116034. [Google Scholar] [CrossRef]
- Silva, V.; Almeida, L.; Gaio, V.; Cerca, N.; Manageiro, V.; Caniça, M.; Capelo, J.L.; Igrejas, G.; Poeta, P. Biofilm Formation of Multidrug-Resistant MRSA Strains Isolated from Different Types of Human Infections. Pathogens 2021, 10, 970. [Google Scholar] [CrossRef]
- Ryder, V.J.; Chopra, I.; O’Neill, A.J. Increased mutability of Staphylococci in biofilms as a consequence of oxidative stress. PLoS ONE 2012, 7, e47695. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic | Resistant M ± SD | Susceptible M ± SD | p |
|---|---|---|---|
| Penicillin | 123.469 ± 38.554 | 116.446± 28.214 | 0.262 |
| Cefoxitin | 134.263 ± 47.678 | 115.547 ± 26.949 | <0.001 |
| Ciprofloxacin | 112.162 ± 22.167 | 123.607 ± 37.029 | 0.016 |
| Gentamycin | 121.581 ± 43.431 | 122.230 ± 35.592 | 0.462 |
| Tobramycin | 121.041 ± 37.159 | 122.504 ± 36.848 | 0.398 |
| Kanamycin | 116.791 ± 38.061 | 123.691 ± 36.453 | 0.120 |
| Erythromycin | 127.907 ± 43.901 | 116.695 ± 27.816 | 0.11 |
| Clindamycin | 127.057 ± 42.489 | 116.182 ± 27.673 | 0.014 |
| Tetracycline | 131.639 ± 44.944 | 114.379 ± 26.382 | <0.001 |
| Chloramphenicol | 129.674 ± 42.052 | 116.704 ± 27.575 | 0.039 |
| Fusidic acid | 140.587 ± 48.318 | 113.390 ± 25.542 | <0.001 |
| Trimethoprim- sulfamethoxazole | 103.112 ± 13.315 | 125.025 ± 37.153 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, V.; Correia, E.; Pereira, J.E.; González-Machado, C.; Capita, R.; Alonso-Calleja, C.; Igrejas, G.; Poeta, P. Exploring the Biofilm Formation Capacity in S. pseudintermedius and Coagulase-Negative Staphylococci Species. Pathogens 2022, 11, 689. https://doi.org/10.3390/pathogens11060689
Silva V, Correia E, Pereira JE, González-Machado C, Capita R, Alonso-Calleja C, Igrejas G, Poeta P. Exploring the Biofilm Formation Capacity in S. pseudintermedius and Coagulase-Negative Staphylococci Species. Pathogens. 2022; 11(6):689. https://doi.org/10.3390/pathogens11060689
Chicago/Turabian StyleSilva, Vanessa, Elisete Correia, José Eduardo Pereira, Camino González-Machado, Rosa Capita, Carlos Alonso-Calleja, Gilberto Igrejas, and Patrícia Poeta. 2022. "Exploring the Biofilm Formation Capacity in S. pseudintermedius and Coagulase-Negative Staphylococci Species" Pathogens 11, no. 6: 689. https://doi.org/10.3390/pathogens11060689