
Absence of Polymorphisms in Codons 167, 198 and 200 of All Seven β-Tubulin Isotypes of Benzimidazole Susceptible and Resistant Parascaris spp. Specimens from Australia



Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasite Collection
2.2. DNA Isolation
2.3. Identification of β-Tubulin Genes in the Parascaris univalens and Ascaris suum Genomes
2.4. Phylogenetic Analysis
2.5. Comparison of Expression Levels between β-Tubulin Paralogs in Parascaris univalens and Ascaris suum
2.6. PCRs and Sequencing
3. Results
3.1. Nomenclature of Parascaris univalens and Ascaris suum β-Tubulin Paralogs
3.2. Phylogenetic Analysis of the β-Tubulin Repertoire of Ascaris sp. and Parascaris univalens
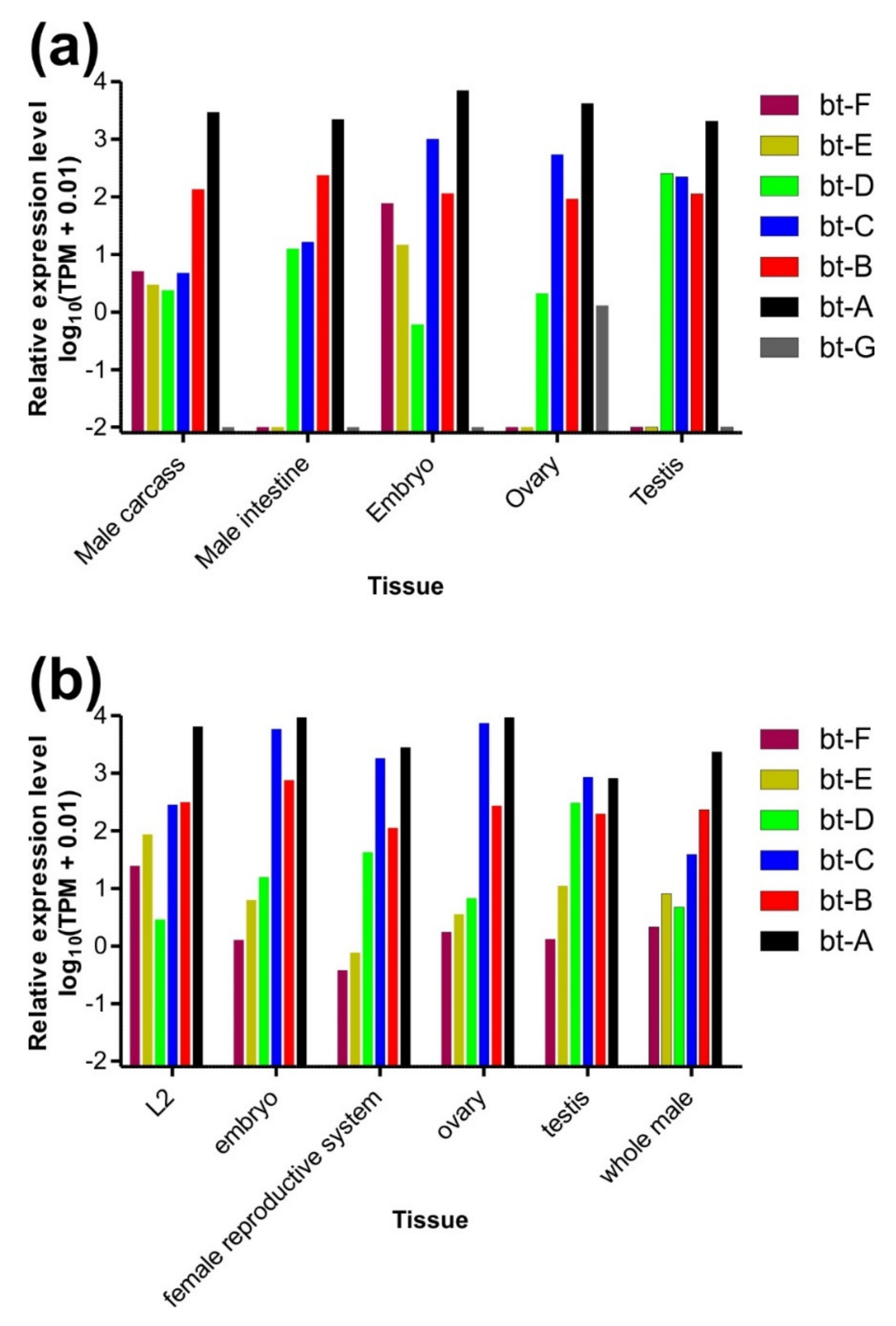
3.3. Expression Levels of β-Tubulin Paralogs in Parascaris univalens and Ascaris suum
3.4. Genotyping of Parascaris spp. Specimens from Australia
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Slocombe, D.O.J. Pathogenesis of helminths in equines. Vet. Parasitol. 1985, 18, 139–153. [Google Scholar] [PubMed]
- Hautala, K.; Näreaho, A.; Kauppinen, O.; Nielsen, M.K.; Sukura, A.; Rajala-Schultz, P.J. Risk factors for equine intestinal parasite infections and reduced efficacy of pyrantel embonate against Parascaris sp. Vet. Parasitol. 2019, 273, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Saeed, M.A.; Beveridge, I.; Abbas, G.; Beasley, A.; Bauquier, J.; Wilkes, E.; Jacobson, C.; Hughes, K.J.; El-Hage, C.; O’Handley, R.; et al. Systematic review of gastrointestinal nematodes of horses from Australia. Parasit. Vectors 2019, 12, 188. [Google Scholar] [CrossRef] [PubMed]
- Misuno, E.; Clark, C.R.; Anderson, S.L.; Jenkins, E.; Wagner, B.; Dembek, K.; Petrie, L. Characteristics of parasitic egg shedding over a 1-year period in foals and their dams in 2 farms in central Saskatchewan. Can. Vet. J. 2018, 59, 284–292. [Google Scholar] [PubMed]
- Nielsen, M.K.; Wang, J.; Davis, R.; Bellaw, J.L.; Lyons, E.T.; Lear, T.L.; Goday, C. Parascaris univalens—A victim of large-scale misidentification? Parasitol. Res. 2014, 113, 4485–4490. [Google Scholar] [CrossRef]
- Bullini, L.; Nascetti, G.; Carrè, S.; Rumore, F.; Biocca, E. Ricerche cariologiche ed elettroforetiche su Parascaris univalens e Parascaris equorum. Acc. Naz. Lincei Rend. Cl. Sc. Fis. Mat. Nat. 1978, 65, 133–141. [Google Scholar]
- Von Samson-Himmelstjerna, G.; Janssen, I.J.I.; Ramünke, S.; Goday, C.; Borges, F.d.A.; Koudela, B.; Niedźwiedź, A.; Tomczuk, K.; Studzińska, M.B.; Kornas, S.; et al. Very low intraspecific sequence variation in selected nuclear and mitochondrial Parascaris univalens genes. Infect. Genet. Evol. 2021, 95, 105035. [Google Scholar] [CrossRef]
- Pimpinelli, S.; Goday, C. Unusual kinetochores and chromatin diminution in Parascaris. Trends Genet. 1989, 5, 310–315. [Google Scholar] [CrossRef]
- Matthews, J.B. Anthelmintic resistance in equine nematodes. Int. J. Parasitol. Drugs drug Resist. 2014, 4, 310–315. [Google Scholar] [CrossRef] [Green Version]
- Peregrine, A.S.; Molento, M.B.; Kaplan, R.M.; Nielsen, M.K. Anthelmintic resistance in important parasites of horses: Does it really matter? Vet. Parasitol. 2014, 201, 1–8. [Google Scholar] [CrossRef]
- Von Samson-Himmelstjerna, G. Anthelmintic resistance in equine parasites - detection, potential clinical relevance and implications for control. Vet. Parasitol. 2012, 185, 2–8. [Google Scholar] [CrossRef]
- Raza, A.; Qamar, A.G.; Hayat, K.; Ashraf, S.; Williams, A.R. Anthelmintic resistance and novel control options in equine gastrointestinal nematodes. Parasitology 2019, 146, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Craig, T.M.; Diamond, P.L.; Ferwerda, N.S.; Thompson, J.A. Evidence of Ivermectin Resistance by Parascaris equorum on a Texas Horse Farm. J. Equine Vet. Sci. 2007, 27, 67–71. [Google Scholar] [CrossRef]
- Alanazi, A.D.; Mukbel, R.M.; Alyousif, M.S.; AlShehri, Z.S.; Alanazi, I.O.; Al-Mohammed, H.I. A field study on the anthelmintic resistance of Parascaris spp. in Arab foals in the Riyadh region, Saudi Arabia. Vet. Q. 2017, 37, 200–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, S.K.; Woodgate, R.G.; Gough, S.; Heller, J.; Sangster, N.C.; Hughes, K.J. The efficacy of ivermectin, pyrantel and fenbendazole against Parascaris equorum infection in foals on farms in Australia. Vet. Parasitol. 2014, 205, 575–580. [Google Scholar] [CrossRef]
- Martin, F.; Halvarsson, P.; Delhomme, N.; Höglund, J.; Tydén, E. Exploring the β-tubulin gene family in a benzimidazole-resistant Parascaris univalens population. Int. J. Parasitol. Drugs Drug Resist. 2021, 17, 84–91. [Google Scholar] [CrossRef]
- Kotze, A.C.; Hunt, P.W.; Skuce, P.; von Samson-Himmelstjerna, G.; Martin, R.J.; Sager, H.; Krücken, J.; Hodgkinson, J.; Lespine, A.; Jex, A.R.; et al. Recent advances in candidate-gene and whole-genome approaches to the discovery of anthelmintic resistance markers and the description of drug/receptor interactions. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 164–184. [Google Scholar] [CrossRef] [Green Version]
- Ridley, R.K.; Dryden, M.W.; Gabbert, N.H.; Schoning, P. Epidemiology and control of helminth parasites in Greyhound breeding farms. Compend. Contin. Educ. Pract. Vet. 1994, 16, 585–599. [Google Scholar]
- Collins, J.B.; Jordan, B.; Baldwin, L.; Hebron, C.; Paras, K.; Vidyashankar, A.N.; Kaplan, R.M. Resistance to fenbendazole in Ascaridia dissimilis, an important nematode parasite of turkeys. Poult. Sci. 2019, 98, 5412–5415. [Google Scholar] [CrossRef]
- Krücken, J.; Fraundorfer, K.; Mugisha, J.C.; Ramünke, S.; Sifft, K.C.; Geus, D.; Habarugira, F.; Ndoli, J.; Sendegeya, A.; Mukampunga, C.; et al. Reduced efficacy of albendazole against Ascaris lumbricoides in Rwandan schoolchildren. Int. J. Parasitol. Drugs Drug Resist. 2017, 7, 262–271. [Google Scholar] [CrossRef]
- Kwa, M.S.G.; Veenstra, J.G.; Van Dijk, M.; Roos, M.H. β-Tubulin Genes from the Parasitic Nematode—Modulate Drug Resistance in Caenorhabditis elegans. J. Mol. Biol. 1995, 246, 500–510. [Google Scholar] [CrossRef]
- Kwa, M.S.G.; Veenstra, J.G.; Roos, M.H. Benzimidazole resistance in Haemonchus contortus is correlated with a conserved mutation at amino acid 200 in β-tubulin isotype 1. Mol. Biochem. Parasitol. 1994, 63, 299–303. [Google Scholar] [CrossRef]
- Pape, M.; Posedi, J.; Failing, K.; Schnieder, T.; von Samson-Himmelstjerna, G. Analysis of the beta-tubulin codon 200 genotype distribution in a benzimidazole-susceptible and -resistant cyathostome population. Parasitology 2003, 127, S0031182003003317. [Google Scholar] [CrossRef] [PubMed]
- Prichard, R. Genetic variability following selection of Haemonchus contortus with anthelmintics. Trends Parasitol. 2001, 17, 445–453. [Google Scholar] [CrossRef]
- Ghisi, M.; Kaminsky, R.; Mäser, P. Phenotyping and genotyping of Haemonchus contortus isolates reveals a new putative candidate mutation for benzimidazole resistance in nematodes. Vet. Parasitol. 2007, 144, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Valladares, M.; Valderas-García, E.; Gandasegui, J.; Skuce, P.; Morrison, A.; Castilla Gómez de Agüero, V.; Cambra-Pellejà, M.; Balaña-Fouce, R.; Rojo-Vázquez, F.A. Teladorsagia circumcincta beta tubulin: The presence of the E198L polymorphism on its own is associated with benzimidazole resistance. Parasit. Vectors 2020, 13, 453. [Google Scholar] [CrossRef] [PubMed]
- Mohammedsalih, K.M.; Krücken, J.; Bashar, A.; Juma, F.-R.; Abdalmalaik, A.A.H.; Khalafalla, A.; Abakar, A.; Coles, G.; von Samson-Himmelstjerna, G. Susceptible trichostrongyloid species mask presence of benzimidazole-resistant Haemonchus contortus in cattle. Parasit. Vectors 2021, 14, 101. [Google Scholar] [CrossRef]
- Mohammedsalih, K.M.; Krücken, J.; Khalafalla, A.; Bashar, A.; Juma, F.-R.; Abakar, A.; Abdalmalaik, A.A.H.; Coles, G.; von Samson-Himmelstjerna, G. New codon 198 β-tubulin polymorphisms in highly benzimidazole resistant Haemonchus contortus from goats in three different states in Sudan. Parasit. Vectors 2020, 13, 114. [Google Scholar] [CrossRef] [Green Version]
- Avramenko, R.W.; Redman, E.M.; Melville, L.; Bartley, Y.; Wit, J.; Queiroz, C.; Bartley, D.J.; Gilleard, J.S. Deep amplicon sequencing as a powerful new tool to screen for sequence polymorphisms associated with anthelmintic resistance in parasitic nematode populations. Int. J. Parasitol. 2019, 49, 13–26. [Google Scholar] [CrossRef]
- Redman, E.; Whitelaw, F.; Tait, A.; Burgess, C.; Bartley, Y.; Skuce, P.J.; Jackson, F.; Gilleard, J.S. The emergence of resistance to the benzimidazole anthlemintics in parasitic nematodes of livestock is characterised by multiple independent hard and soft selective sweeps. PLoS Negl. Trop. Dis. 2015, 9, e0003494. [Google Scholar] [CrossRef] [Green Version]
- Baltrušis, P.; Halvarsson, P.; Höglund, J. Exploring benzimidazole resistance in Haemonchus contortus by next generation sequencing and droplet digital PCR. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Fávero, F.C.; Dos Santos, L.B.; de Araújo, F.R.; Ramünke, S.; Krücken, J.; von Samson-Himmelstjerna, G.; Borges, F.d.A. Haemonchus sp. in beef cattle in Brazil: Species composition and frequency of benzimidazole resistance alleles. Prev. Vet. Med. 2020, 185, 105162. [Google Scholar] [CrossRef] [PubMed]
- Ramünke, S.; Melville, L.; Rinaldi, L.; Hertzberg, H.; de Waal, T.; von Samson-Himmelstjerna, G.; Cringoli, G.; Mavrot, F.; Skuce, P.; Krücken, J.; et al. Benzimidazole resistance survey for Haemonchus, Teladorsagia and Trichostrongylus in three European countries using pyrosequencing including the development of new assays for Trichostrongylus. Int. J. Parasitol. Drugs drug Resist. 2016, 6, 230–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteban-Ballesteros, M.; Rojo-Vázquez, F.A.; Skuce, P.J.; Melville, L.; González-Lanza, C.; Martínez-Valladares, M. Quantification of resistant alleles in the β-tubulin gene of field strains of gastrointestinal nematodes and their relation with the faecal egg count reduction test. BMC Vet. Res. 2017, 13, 71. [Google Scholar] [CrossRef] [Green Version]
- Ademola, I.O.; Krücken, J.; Ramünke, S.; Demeler, J.; von Samson-Himmelstjerna, G. Absence of detectable benzimidazole-resistance associated alleles in Haemonchus placei in cattle in Nigeria revealed by pyrosequencing of β-tubulin isotype 1. Parasitol. Res. 2015, 114, 1997–2001. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Valladares, M.; Donnan, A.; Geldhof, P.; Jackson, F.; Rojo-Vázquez, F.-A.; Skuce, P. Pyrosequencing analysis of the beta-tubulin gene in Spanish Teladorsagia circumcincta field isolates. Vet. Parasitol. 2012, 184, 371–376. [Google Scholar] [CrossRef]
- Barrère, V.; Alvarez, L.; Suarez, G.; Ceballos, L.; Moreno, L.; Lanusse, C.; Prichard, R.K. Relationship between increased albendazole systemic exposure and changes in single nucleotide polymorphisms on the β-tubulin isotype 1 encoding gene in Haemonchus contortus. Vet. Parasitol. 2012, 186, 344–349. [Google Scholar] [CrossRef]
- Chaudhry, U.; Redman, E.M.; Raman, M.; Gilleard, J.S. Genetic evidence for the spread of a benzimidazole resistance mutation across southern India from a single origin in the parasitic nematode Haemonchus contortus. Int. J. Parasitol. 2015, 45, 721–728. [Google Scholar] [CrossRef]
- Chaudhry, U.; Miller, M.; Yazwinski, T.; Kaplan, R.; Gilleard, J. The presence of benzimidazole resistance mutations in Haemonchus placei from US cattle. Vet. Parasitol. 2014, 204, 411–415. [Google Scholar] [CrossRef]
- Baltrušis, P.; Halvarsson, P.; Höglund, J. Utilization of droplet digital PCR to survey resistance associated polymorphisms in the β tubulin gene of Haemonchus contortus in sheep flocks in Sweden. Vet. Parasitol. 2020, 288, 109278. [Google Scholar] [CrossRef]
- Sargison, N.D.; MacLeay, M.; Morrison, A.A.; Bartley, D.J.; Evans, M.; Chaudhry, U. Development of amplicon sequencing for the analysis of benzimidazole resistance allele frequencies in field populations of gastrointestinal nematodes. Int. J. Parasitol. Drugs drug Resist. 2019, 10, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Diawara, A.; Halpenny, C.M.; Churcher, T.S.; Mwandawiro, C.; Kihara, J.; Kaplan, R.M.; Streit, T.G.; Idaghdour, Y.; Scott, M.E.; Basáñez, M.-G.; et al. Association between response to albendazole treatment and β-tubulin genotype frequencies in soil-transmitted helminths. PLoS Negl. Trop. Dis. 2013, 7, e2247. [Google Scholar] [CrossRef] [PubMed]
- Roose, S.; Avramenko, R.W.; Pollo, S.M.J.; Wasmuth, J.D.; Ame, S.; Ayana, M.; Betson, M.; Cools, P.; Dana, D.; Jones, B.P.; et al. Characterization of the β-tubulin gene family in Ascaris lumbricoides and Ascaris suum and its implication for the molecular detection of benzimidazole resistance. PLoS Negl. Trop. Dis. 2021, 15, e0009777. [Google Scholar] [CrossRef] [PubMed]
- Saunders, G.I.; Wasmuth, J.D.; Beech, R.; Laing, R.; Hunt, M.; Naghra, H.; Cotton, J.A.; Berriman, M.; Britton, C.; Gilleard, J.S. Characterization and comparative analysis of the complete Haemonchus contortus β-tubulin gene family and implications for benzimidazole resistance in strongylid nematodes. Int. J. Parasitol. 2013, 43, 465–475. [Google Scholar] [CrossRef]
- Driscoll, M.; Dean, E.; Reilly, E.; Bergholz, E.; Chalfie, M. Genetic and molecular analysis of a Caenorhabditis elegans beta-tubulin that conveys benzimidazole sensitivity. J. Cell Biol. 1989, 109, 2993–3003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dilks, C.M.; Hahnel, S.R.; Sheng, Q.; Long, L.; McGrath, P.T.; Andersen, E.C. Quantitative benzimidazole resistance and fitness effects of parasitic nematode beta-tubulin alleles. Int. J. Parasitol. Drugs Drug Resist. 2020, 14, 28–36. [Google Scholar] [CrossRef]
- Dilks, C.M.; Koury, E.J.; Buchanan, C.M.; Andersen, E.C. Newly identified parasitic nematode beta-tubulin alleles confer resistance to benzimidazoles. Int. J. Parasitol. Drugs Drug Resist. 2021, 17, 168–175. [Google Scholar] [CrossRef]
- Hahnel, S.R.; Zdraljevic, S.; Rodriguez, B.C.; Zhao, Y.; McGrath, P.T.; Andersen, E.C. Extreme allelic heterogeneity at a Caenorhabditis elegans beta-tubulin locus explains natural resistance to benzimidazoles. PLoS Pathog. 2018, 14, e1007226. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Lü, L.; Nadler, S.A.; Gibson, D.I.; Zhang, L.-P.; Chen, H.-X.; Zhao, W.-T.; Guo, Y.-N. Molecular Phylogeny and Dating Reveal a Terrestrial Origin in the Early Carboniferous for Ascaridoid Nematodes. Syst. Biol. 2018, 67, 888–900. [Google Scholar] [CrossRef] [Green Version]
- Wilkes, E.J.A.; Cowling, A.; Woodgate, R.G.; Hughes, K.J. Inhomogeneity of the density of Parascaris spp. eggs in faeces of individual foals and the use of hypothesis testing for treatment decision making. Vet. Parasitol. 2016, 229, 131–138. [Google Scholar] [CrossRef]
- Wang, J.; Mitreva, M.; Berriman, M.; Thorne, A.; Magrini, V.; Koutsovoulos, G.; Kumar, S.; Blaxter, M.L.; Davis, R.E. Silencing of germline-expressed genes by DNA elimination in somatic cells. Dev. Cell 2012, 23, 1072–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Gao, S.; Mostovoy, Y.; Kang, Y.; Zagoskin, M.; Sun, Y.; Zhang, B.; White, L.K.; Easton, A.; Nutman, T.B.; et al. Comparative genome analysis of programmed DNA elimination in nematodes. Genome Res. 2017, 27, 2001–2014. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Howe, K.L.; Bolt, B.J.; Shafie, M.; Kersey, P.; Berriman, M. WormBase ParaSite—A comprehensive resource for helminth genomics. Mol. Biochem. Parasitol. 2017, 215, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Xia, X. DAMBE7: New and Improved Tools for Data Analysis in Molecular Biology and Evolution. Mol. Biol. Evol. 2018, 35, 1550–1552. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Lemey, P. Assessing substitution saturation with DAMBE. In The Phylogenetic Handbook: A Practical Approach to Phylogenetic Analysis and Hypothesis Testing; Lemey, P., Salemi, M., Vandamme, A.-M., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 615–630. [Google Scholar]
- Xia, X.; Xie, Z.; Salemi, M.; Chen, L.; Wang, Y. An index of substitution saturation and its application. Mol. Phylogenet. Evol. 2003, 26, 1–7. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisimova, M.; Gil, M.; Dufayard, J.-F.; Dessimoz, C.; Gascuel, O. Survey of branch support methods demonstrates accuracy, power, and robustness of fast likelihood-based approximation schemes. Syst. Biol. 2011, 60, 685–699. [Google Scholar] [CrossRef] [PubMed]
- Beech, R.N.; Wolstenholme, A.J.; Neveu, C.; Dent, J.A. Nematode parasite genes: What’s in a name? Trends Parasitol. 2010, 26, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Vercruysse, J.; Albonico, M.; Behnke, J.M.; Kotze, A.C.; Prichard, R.K.; McCarthy, J.S.; Montresor, A.; Levecke, B. Is anthelmintic resistance a concern for the control of human soil-transmitted helminths? Int. J. Parasitol. Drugs Drug Resist. 2011, 1, 14–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organisation Guideline: Preventive Chemotherapy to Control Soil-Transmitted Helminth Infections in At-Risk Population Groups. Available online: https://apps.who.int/iris/handle/10665/258983 (accessed on 20 March 2022).
- Ma, Z.; Michailides, T.J. Advances in understanding molecular mechanisms of fungicide resistance and molecular detection of resistant genotypes in phytopathogenic fungi. Crop Prot. 2005, 24, 853–863. [Google Scholar] [CrossRef]
- Stasiuk, S.J.; MacNevin, G.; Workentine, M.L.; Gray, D.; Redman, E.; Bartley, D.; Morrison, A.; Sharma, N.; Colwell, D.; Ro, D.K.; et al. Similarities and differences in the biotransformation and transcriptomic responses of Caenorhabditis elegans and Haemonchus contortus to five different benzimidazole drugs. Int. J. Parasitol. Drugs Drug Resist. 2019, 11, 13–29. [Google Scholar] [CrossRef]
- Stuchlíková, L.R.; Matoušková, P.; Vokřál, I.; Lamka, J.; Szotáková, B.; Sečkařová, A.; Dimunová, D.; Nguyen, L.T.; Várady, M.; Skálová, L. Metabolism of albendazole, ricobendazole and flubendazole in Haemonchus contortus adults: Sex differences, resistance-related differences and the identification of new metabolites. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 50–58. [Google Scholar] [CrossRef]
- Kellerová, P.; Raisová Stuchlíková, L.; Matoušková, P.; Štěrbová, K.; Lamka, J.; Navrátilová, M.; Vokřál, I.; Szotáková, B.; Skálová, L. Sub-lethal doses of albendazole induce drug metabolizing enzymes and increase albendazole deactivation in Haemonchus contortus adults. Vet. Res. 2020, 51, 94. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Nomenclature Roose et al. [43] | Nomenclature Martin et al. [16] | Previous Names a | Accession no. b Parascaris univalens | Accession no. b Ascaris suum | Accession no. b Ascaris lumbricoidea |
|---|---|---|---|---|---|
| bt-A | tbb-5 | β-tubulin tbb-1 tbb-1.2 β-tubulin isotype 1 | PgR007_g022_t01 | AgB02_g235_t03 | ALUE_0000927201 |
| bt-B | tbb-8b | tbb-2 | PgR003_g161_t01 | AgR022_g106_t03 | ALUE_0000986501 |
| bt-B’ | tbb-8a | ALUE_0001827701 | |||
| bt-C | tbb-6 | tbb-1 | PgE153_g002_t01 | AgE31_g003_t01 | ALUE_0000494801 |
| bt-D | tbb-7 | PgB10_g062_t01 | AgR043_g091_t01 | ALUE_0001031701 | |
| bt-E | tbb-4 | tbb-4 | PgB004_g136_t01 | AgB01_g252_t02 | ALUE_0000949301 |
| bt-F | tbb-3 | PgB04_g135_t01 | AgB01_g251_t03 | ALUE_0000949201 | |
| bt-G | tbb-9 | PgR045_g070_t01 | CM024273: 10203269-10211838 | ALUE_0001294101 |
| β-Tubulin Paralog | PgB04_g135/bt-E | PgB04_g136/bt-F | |||||
|---|---|---|---|---|---|---|---|
| Sample ID | Res. status a | Codon 167 | Codon 198 | Codon 200 | Codon 167 | Codon 198 | Codon 200 |
| S1 | S | F | E | F | F | E | F |
| S2 | S | F | E | F | F | E | F |
| S3 | S | F | E | F | F | E | F |
| S4 | S | F | E | F | F | E | F |
| S5 | S | F | E | F | F | E | F |
| S6 | S | F | E | F | F | E | F |
| S7 | S | F | E | F | F | E | F |
| S8 | S | F | E | F | F | E | F |
| S9 | S | F | E | F | F | E | F |
| S10 | S | F | E | F | F | E | F |
| S11 | S | F | E | F | F | E | F |
| S12 | S | F | E | F | F | E | F |
| S13 | S | F | E | F | n.a. | n.a. | n.a. |
| R1 | S | F | E | F | n.a. | n.a. | n.a. |
| R2 | R | F | E | F | F | E | F |
| R3 | R | F | E | F | F | E | F |
| R4 | R | F | E | F | F | E | F |
| R5 | R | F | E | F | F | E | F |
| R6 | R | F | E | F | F | E | F |
| R7 | R | F | E | F | F | E | F |
| R8 | R | F | E | F | F | E | F |
| R9 | R | F | E | F | n.a. | n.a. | n.a. |
| R11 | R | F | E | F | F | E | F |
| β-Tubulin Paralog | PgR007_g022/bt-A | PgR003_g161/bt-B | PgE153_g002/bt-C | PgB10_g062/bt-D | PgR045_g070/bt-G | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample ID | Res. statusa | Codon 167 | Codon 198 | Codon 200 | Codon 167 | Codon 198 | Codon 200 | Codon 167 | Codon 198 | Codon 200 | Codon 167 | Codon 198 | Codon 200 | Codon 167 | Codon 198 | Codon 200 |
| S1 | S | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| S2 | S | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| S3 | S | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| S4 | S | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| S5 | S | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| S6 | S | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| S7 | S | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| S8 | S | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| S9 | S | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| S10 | S | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| S11 | S | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| S12 | S | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| S13 | S | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| R1 | S | F | E | F | n.a. | n.a. | n.a. | F | E | F | F | E | Y | F | E | F |
| R2 | R | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| R3 | R | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| R4 | R | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| R5 | R | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| R6 | R | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| R7 | R | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| R8 | R | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| R9 | R | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
| R11 | R | F | E | F | F | E | F | F | E | F | F | E | Y | F | E | F |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Özben, M.; von Samson-Himmelstjerna, G.; Freiin von Streit, M.K.B.; Wilkes, E.J.A.; Hughes, K.J.; Krücken, J. Absence of Polymorphisms in Codons 167, 198 and 200 of All Seven β-Tubulin Isotypes of Benzimidazole Susceptible and Resistant Parascaris spp. Specimens from Australia. Pathogens 2022, 11, 490. https://doi.org/10.3390/pathogens11050490
Özben M, von Samson-Himmelstjerna G, Freiin von Streit MKB, Wilkes EJA, Hughes KJ, Krücken J. Absence of Polymorphisms in Codons 167, 198 and 200 of All Seven β-Tubulin Isotypes of Benzimidazole Susceptible and Resistant Parascaris spp. Specimens from Australia. Pathogens. 2022; 11(5):490. https://doi.org/10.3390/pathogens11050490
Chicago/Turabian StyleÖzben, Murat, Georg von Samson-Himmelstjerna, Malene K. B. Freiin von Streit, Edwina J. A. Wilkes, Kristopher J. Hughes, and Jürgen Krücken. 2022. "Absence of Polymorphisms in Codons 167, 198 and 200 of All Seven β-Tubulin Isotypes of Benzimidazole Susceptible and Resistant Parascaris spp. Specimens from Australia" Pathogens 11, no. 5: 490. https://doi.org/10.3390/pathogens11050490