
Prevalence and Antimicrobial Resistance of Staphylococcus aureus and Coagulase-Negative Staphylococcus/Mammaliicoccus from Retail Ground Meat: Identification of Broad Genetic Diversity in Fosfomycin Resistance Gene fosB

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Isolation of Staphylococcus/Mammaliicoccus Isolates
2.2. AMR and Genetic Characterization of S. aureus
2.3. AMR and Antimicrobial Resistance Genes in CoNS/Mammaliicoccus
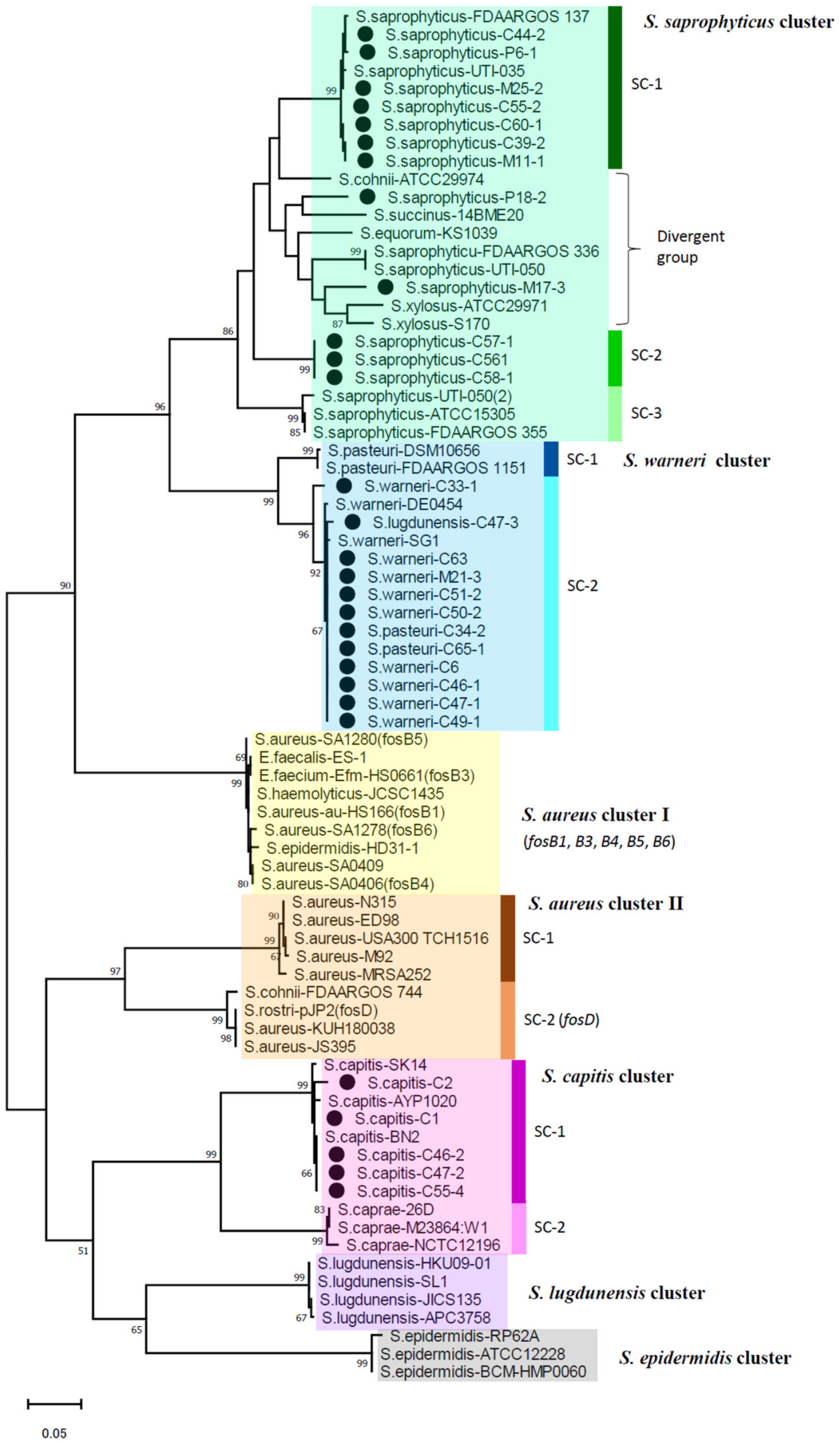
2.4. Phylogenetic Analysis of fosB
2.5. Prevalence of 6-TG Synthesis Genes among CoNS/Mammaliicoccus
3. Discussion
4. Materials and Methods
4.1. Isolation and Identification of Staphylococcus/Mammaliicoccus Species
4.2. Antimicrobial Susceptibility Test
4.3. Detection of Antimicrobial Resistance Genes, Genetic Analysis of fosB
4.4. Genotyping and Detection of Virulence Factors
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Götz, F.; Bannerman, T.; Schleifer, K.H. The Genera Staphylococcus and Macrococcus. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 5–75. [Google Scholar]
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-negative staphylococci. Clin. Microbiol. Rev. 2014, 27, 870–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.Y.C.; Schaumburg, F.; Ellington, M.J.; Corander, J.; Pichon, B.; Leendertz, F.; Bentley, S.D.; Parkhill, J.; Holt, D.C.; Peters, G.; et al. Novel staphylococcal species that form part of a Staphylococcus aureus-related complex: The non-pigmented Staphylococcus argenteus sp. nov. and the non-human primate-associated Staphylococcus schweitzeri sp. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Chew, K.L.; Octavia, S.; Lai, D.; Lin, R.T.P.; Teo, J.W.P. Staphylococcus singaporensis sp. nov., a new member of the Staphylococcus aureus complex, isolated from human clinical specimens. Int. J. Syst. Evol. Microbiol. 2021, 71, 005067. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Schaumburg, F.; Kearns, A.; Larsen, A.R.; Lindsay, J.A.; Skov, R.L.; Westh, H. Implications of identifying the recently defined members of the Staphylococcus aureus complex S. argenteus and S. schweitzeri: A position paper of members of the ESCMID Study Group for Staphylococci and Staphylococcal Diseases (ESGS). Clin. Microbiol. Infect. 2019, 25, 1064–1070. [Google Scholar] [CrossRef]
- Heilmann, C.; Ziebuhr, W.; Becker, K. Are coagulase-negative staphylococci virulent? Clin. Microbiol. Infect. 2019, 25, 1071–1080. [Google Scholar] [CrossRef]
- Madhaiyan, M.; Wirth, J.S.; Saravanan, V.S. Phylogenomic analyses of the Staphylococcaceae family suggest the reclassification of five species within the genus Staphylococcus as heterotypic synonyms, the promotion of five subspecies to novel species, the taxonomic reassignment of five Staphylococcus species to Mammaliicoccus gen. nov., and the formal assignment of Nosocomiicoccus to the family Staphylococcaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 5926–5936. [Google Scholar]
- Argudín, M.A.; Deplano, A.; Meghraoui, A.; Dodémont, M.; Heinrichs, A.; Denis, O.; Nonhoff, C.; Roisin, S. Bacteria from Animals as a Pool of Antimicrobial Resistance Genes. Antibiotics 2017, 6, 12. [Google Scholar] [CrossRef]
- Tsubakishita, S.; Kuwahara-Arai, K.; Sasaki, T.; Hiramatsu, K. Origin and molecular evolution of the determinant of methicillin resistance in staphylococci. Antimicrob. Agents Chemother. 2020, 54, 4352–4359. [Google Scholar] [CrossRef] [Green Version]
- Huber, H.; Ziegler, D.; Pflüger, V.; Vogel, G.; Zweifel, C.; Stephan, R. Prevalence and characteristics of methicillin-resistant coagulase-negative staphylococci from livestock, chicken carcasses, bulk tank milk, minced meat, and contact persons. BMC Vet. Res. 2011, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Osman, K.M.; Amer, A.M.; Badr, J.M.; Saad, A.S. Prevalence and antimicrobial resistance profile of Staphylococcus species in chicken and beef raw meat in Egypt. Foodborne Pathog. Dis. 2015, 12, 406–413. [Google Scholar] [CrossRef]
- Can, H.Y.; Elmalı, M.; Karagöz, A. Molecular Typing and Antimicrobial Susceptibility of Staphylococcus aureus Strains Isolated from Raw Milk, Cheese, Minced Meat, and Chicken Meat Samples. Korean J. Food Sci. Anim. Resour 2017, 37, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Fox, A.; Pichon, B.; Wilkinson, H.; Doumith, M.; Hill, R.L.; McLauchlin, J.; Kearns, A.M. Detection and molecular characterization of Livestock-Associated MRSA in raw meat on retail sale in North West England. Lett. Appl. Microbiol. 2017, 64, 239–245. [Google Scholar] [CrossRef]
- Ge, B.; Mukherjee, S.; Hsu, C.H.; Davis, J.A.; Tran, T.T.T.; Yang, Q.; Abbott, J.W.; Ayers, S.L.; Young, S.R.; Crarey, E.T.; et al. MRSA and multidrug-resistant Staphylococcus aureus in U.S. retail meats, 2010–2011. Food Microbiol. 2017, 62, 289–297. [Google Scholar] [CrossRef]
- Tang, Y.; Larsen, J.; Kjeldgaard, J.; Andersen, P.S.; Skov, R.; Ingmer, H. Methicillin-resistant and -susceptible Staphylococcus aureus from retail meat in Denmark. Int. J. Food Microbiol. 2017, 249, 72–76. [Google Scholar] [CrossRef]
- Wu, S.; Huang, J.; Wu, Q.; Zhang, J.; Zhang, F.; Yang, X.; Wu, H.; Zeng, H.; Chen, M.; Ding, Y.; et al. Staphylococcus aureus Isolated From Retail Meat and Meat Products in China: Incidence, Antibiotic Resistance and Genetic Diversity. Front. Microbiol. 2018, 9, 2767. [Google Scholar] [CrossRef]
- Islam, M.A.; Parveen, S.; Rahman, M.; Huq, M.; Nabi, A.; Khan, Z.U.M.; Ahmed, N.; Wagenaar, J.A. Occurrence and Characterization of Methicillin Resistant Staphylococcus aureus in Processed Raw Foods and Ready-to-Eat Foods in an Urban Setting of a Developing Country. Front. Microbiol. 2019, 10, 503. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, H.S.; Kim, S.; Kim, M.; Kwak, H.S. Prevalence and Characteristics of Antimicrobial-Resistant Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus from Retail Meat in Korea. Food Sci. Anim. Resour 2020, 40, 758–771. [Google Scholar] [CrossRef]
- Tanomsridachchai, W.; Changkaew, K.; Changkwanyeun, R.; Prapasawat, W.; Intarapuk, A.; Fukushima, Y.; Yamasamit, N.; Flav Kapalamula, T.; Nakajima, C.; Suthienkul, C.; et al. Antimicrobial Resistance and Molecular Characterization of Methicillin-Resistant Staphylococcus aureus Isolated from Slaughtered Pigs and Pork in the Central Region of Thailand. Antibiotics 2021, 10, 206. [Google Scholar] [CrossRef]
- Tegegne, H.A.; Koláčková, I.; Florianová, M.; Gelbíčová, T.; Madec, J.Y.; Haenni, M.; Karpíšková, R. Detection and molecular characterisation of methicillin-resistant Staphylococcus aureus isolated from raw meat in the retail market. J. Glob. Antimicrob. Resist. 2021, 26, 233–238. [Google Scholar] [CrossRef]
- Thwala, T.; Madoroba, E.; Basson, A.; Butaye, P. Prevalence and Characteristics of Staphylococcus aureus Associated with Meat and Meat Products in African Countries: A Review. Antibiotics 2021, 10, 1108. [Google Scholar] [CrossRef]
- Şanlıbaba, P. Prevalence, antibiotic resistance, and enterotoxin production of Staphylococcus aureus isolated from retail raw beef, sheep, and lamb meat in Turkey. Int. J. Food Microbiol. 2022, 361, 109461. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.I.; Kim, S.D.; Park, J.H.; Yang, S.J. Species Distribution, Antimicrobial Resistance, and Enterotoxigenicity of Non-aureus Staphylococci in Retail Chicken Meat. Antibiotics 2020, 9, 809. [Google Scholar] [CrossRef] [PubMed]
- Kitai, S.; Shimizu, A.; Kawano, J.; Sato, E.; Nakano, C.; Kitagawa, H.; Fujio, K.; Matsumura, K.; Yasuda, R.; Inamoto, T. Prevalence and characterization of Staphylococcus aureus and enterotoxigenic Staphylococcus aureus in retail raw chicken meat throughout Japan. J. Vet. Med. Sci. 2005, 67, 269–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiroi, M.; Kawamori, F.; Harada, T.; Sano, Y.; Miwa, N.; Sugiyama, K.; Hara-Kudo, Y.; Masuda, T. Antibiotic resistance in bacterial pathogens from retail raw meats and food-producing animals in Japan. J. Food Prot. 2012, 75, 1774–1782. [Google Scholar] [CrossRef]
- Sato, T.; Usui, M.; Konishi, N.; Kai, A.; Matsui, H.; Hanaki, H.; Tamura, Y. Closely related methicillin-resistant Staphylococcus aureus isolates from retail meat, cows with mastitis, and humans in Japan. PLoS ONE 2017, 12, e0187319. [Google Scholar] [CrossRef]
- Candel, F.J.; Matesanz David, M.; Barberán, J. New perspectives for reassessing fosfomycin: Applicability in current clinical practice. Rev. Esp. Quimioter. 2019, 32 (Suppl. S1), 1–7. [Google Scholar]
- Castañeda-García, A.; Blázquez, J.; Rodríguez-Rojas, A. Molecular Mechanisms and Clinical Impact of Acquired and Intrinsic Fosfomycin Resistance. Antibiotics 2013, 2, 217–236. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Liu, Y.; Chen, C.; Guo, Y.; Ma, Y.; Yang, Y.; Hu, F.; Xu, X.; Wang, M. Characterization of Fosfomycin Resistance Gene, fosB, in Methicillin-Resistant Staphylococcus aureus Isolates. PLoS ONE 2016, 11, e0154829. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.C.; Chen, P.Y.; Wang, J.T.; Chang, S.C. Prevalence of fosfomycin resistance and gene mutations in clinical isolates of methicillin-resistant Staphylococcus aureus. Antimicrob. Resist. Infect. Control 2020, 9, 135. [Google Scholar] [CrossRef]
- Campanile, F.; Wootton, M.; Davies, L.; Aprile, A.; Mirabile, A.; Pomponio, S.; Demetrio, F.; Bongiorno, D.; Walsh, T.R.; Stefani, S.; et al. Gold standard susceptibility testing of fosfomycin in Staphylococcus aureus and Enterobacterales using a new agar dilution panel®. J. Glob. Antimicrob. Resist. 2020, 23, 334–337. [Google Scholar] [CrossRef]
- Song, Z.; Wang, X.; Zhou, X.; Jiang, S.; Li, Y.; Ahmad, O.; Qi, L.; Li, P.; Li, J. Taxonomic Distribution of FosB in Human-Microbiota and Activity Comparison of Fosfomycin Resistance. Front. Microbiol. 2019, 10, 200. [Google Scholar] [CrossRef]
- Chin, D.; Goncheva, M.I.; Flannagan, R.S.; Deecker, S.R.; Guariglia-Oropeza, V.; Ensminger, A.W.; Heinrichs, D.E. Coagulase-negative staphylococci release a purine analog that inhibits Staphylococcus aureus virulence. Nat. Commun. 2021, 12, 1887. [Google Scholar] [CrossRef]
- Torki Baghbaderani, Z.; Shakerian, A.; Rahimi, E. Phenotypic and Genotypic Assessment of Antibiotic Resistance of Staphylococcus aureus Bacteria Isolated from Retail Meat. Infect. Drug Resist. 2020, 13, 1339–1349. [Google Scholar] [CrossRef]
- Buyukcangaz, E.; Velasco, V.; Sherwood, J.S.; Stepan, R.M.; Koslofsky, R.J.; Logue, C.M. Molecular typing of Staphylococcus aureus and methicillin-resistant S. aureus (MRSA) isolated from animals and retail meat in North Dakota, United States. Foodborne Pathog. Dis. 2013, 10, 608–617. [Google Scholar] [CrossRef]
- Jackson, C.R.; Davis, J.A.; Barrett, J.B. Prevalence and characterization of methicillin-resistant Staphylococcus aureus isolates from retail meat and humans in Georgia. J. Clin. Microbiol. 2013, 51, 1199–1207. [Google Scholar] [CrossRef] [Green Version]
- Thapaliya, D.; Forshey, B.M.; Kadariya, J.; Quick, M.K.; Farina, S.; O’ Brien, A.; Nair, R.; Nworie, A.; Hanson, B.; Kates, A.; et al. Prevalence and molecular characterization of Staphylococcus aureus in commercially available meat over a one-year period in Iowa, USA. Food Microbiol. 2017, 65, 122–129. [Google Scholar] [CrossRef] [Green Version]
- Anjum, M.F.; Marco-Jimenez, F.; Duncan, D.; Marín, C.; Smith, R.P.; Evans, S.J. Livestock-Associated Methicillin-Resistant Staphylococcus aureus From Animals and Animal Products in the UK. Front. Microbiol. 2019, 10, 2136. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Tang, T.; Stegger, M.; Dalsgaard, A.; Liu, T.; Leisner, J.J. Characterization of antimicrobial-resistant Staphylococcus aureus from retail foods in Beijing, China. Food Microbiol. 2021, 93, 103603. [Google Scholar] [CrossRef]
- Verkade, E.; Kluytmans, J. Livestock-associated Staphylococcus aureus CC398: Animal reservoirs and human infections. Infect. Genet. Evol. 2014, 21, 523–530. [Google Scholar] [CrossRef]
- Hata, E.; Katsuda, K.; Kobayashi, H.; Uchida, I.; Tanaka, K.; Eguchi, M. Genetic variation among Staphylococcus aureus strains from bovine milk and their relevance to methicillin-resistant isolates from humans. J. Clin. Microbiol. 2010, 48, 2130–2139. [Google Scholar] [CrossRef] [Green Version]
- Dastmalchi Saei, H.; Panahi, M. Genotyping and antimicrobial resistance of Staphylococcus aureus isolates from dairy ruminants: Differences in the distribution of clonal types between cattle and small ruminants. Arch. Microbiol. 2020, 202, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Achek, R.; El-Adawy, H.; Hotzel, H.; Hendam, A.; Tomaso, H.; Ehricht, R.; Neubauer, H.; Nabi, I.; Hamdi, T.M.; Monecke, S. Molecular Characterization of Staphylococcus aureus Isolated from Human and Food Samples in Northern Algeria. Pathogens 2021, 10, 1276. [Google Scholar] [CrossRef] [PubMed]
- Rainard, P.; Corrales, J.C.; Barrio, M.B.; Cochard, T.; Poutrel, B. Leucotoxic activities of Staphylococcus aureus strains isolated from cows, ewes, and goats with mastitis: Importance of LukM/LukF’-PV leucotoxin. Clin. Diagn. Lab. Immunol. 2003, 10, 272–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chajęcka-Wierzchowska, W.; Zadernowska, A.; Nalepa, B.; Sierpińska, M.; Łaniewska-Trokenheim, L. Coagulase-negative staphylococci (CoNS) isolated from ready-to-eat food of animal origin—Phenotypic and genotypic antibiotic resistance. Food Microbiol. 2015, 46, 222–226. [Google Scholar] [CrossRef]
- Guran, H.S.; Kahya, S. Species Diversity and Pheno- and Genotypic Antibiotic Resistance Patterns of Staphylococci Isolated from Retail Ground Meats. J. Food Sci. 2015, 80, M1291–M1298. [Google Scholar] [CrossRef]
- Jans, C.; Merz, A.; Johler, S.; Younan, M.; Tanner, S.A.; Kaindi, D.W.M.; Wangoh, J.; Bonfoh, B.; Meile, L.; Tasara, T. East and West African milk products are reservoirs for human and livestock-associated Staphylococcus aureus. Food Microbiol. 2017, 65, 64–73. [Google Scholar] [CrossRef]
- Xu, W.; Chen, T.; Wang, H.; Zeng, W.; Wu, Q.; Yu, K.; Xu, Y.; Zhang, X.; Zhou, T. Molecular Mechanisms and Epidemiology of Fosfomycin Resistance in Staphylococcus aureus Isolated From Patients at a Teaching Hospital in China. Front. Microbiol. 2020, 11, 1290. [Google Scholar] [CrossRef]
- DiCicco, M.; Weese, S.; Neethirajan, S.; Rousseau, J.; Singh, A. Fosfomycin susceptibility of canine methicillin-resistant Staphylococcus pseudintermedius isolates. Res. Vet. Sci. 2014, 96, 251–253. [Google Scholar] [CrossRef]
- Argudín, M.A.; Vanderhaeghen, W.; Butaye, P. Diversity of antimicrobial resistance and virulence genes in methicillin-resistant non-Staphylococcus aureus staphylococci from veal calves. Res. Vet. Sci. 2015, 99, 10–16. [Google Scholar] [CrossRef]
- Higashide, M.; Kuroda, M.; Omura, C.T.; Kumano, M.; Ohkawa, S.; Ichimura, S.; Ohta, T. Methicillin-resistant Staphylococcus saprophyticus isolates carrying staphylococcal cassette chromosome mec have emerged in urogenital tract infections. Antimicrob. Agents Chemother. 2008, 52, 2061–2068. [Google Scholar] [CrossRef] [Green Version]
- Nakaminami, H.; Noguchi, N.; Nishijima, S.; Kurokawa, I.; Sasatsu, M. Characterization of the pTZ2162 encoding multidrug efflux gene qacB from Staphylococcus aureus. Plasmid 2008, 60, 108–117. [Google Scholar] [CrossRef]
- He, T.; Wang, Y.; Schwarz, S.; Zhao, Q.; Shen, J.; Wu, C. Genetic environment of the multi-resistance gene cfr in methicillin-resistant coagulase-negative staphylococci from chickens, ducks, and pigs in China. Int. J. Med. Microbiol. 2014, 304, 257–261. [Google Scholar] [CrossRef]
- Liu, B.H.; Lei, C.W.; Zhang, A.Y.; Pan, Y.; Kong, L.H.; Xiang, R.; Wang, Y.X.; Yang, Y.X.; Wang, H.N. Colocation of the Multiresistance Gene cfr and the Fosfomycin Resistance Gene fosD on a Novel Plasmid in Staphylococcus arlettae from a Chicken Farm. Antimicrob. Agents Chemother. 2017, 61, e01388-17. [Google Scholar] [CrossRef] [Green Version]
- De Freire Bastos, M.D.C.; Miceli de Farias, F.; Carlin Fagundes, P.; Varella Coelho, M.L. Staphylococcins: An update on antimicrobial peptides produced by staphylococci and their diverse potential applications. Appl. Microbiol. Biotechnol. 2020, 104, 10339–10368. [Google Scholar] [CrossRef]
- Hirose, M.; Aung, M.S.; Fukuda, A.; Yahata, S.; Fujita, Y.; Saitoh, M.; Hirose, Y.; Urushibara, N.; Kobayashi, N. Antimicrobial Resistance and Molecular Epidemiological Characteristics of Methicillin-Resistant and Susceptible Staphylococcal Isolates from Oral Cavity of Dental Patients and Staff in Northern Japan. Antibiotics 2021, 10, 1316. [Google Scholar] [CrossRef]
- Zhang, K.; McClure, J.A.; Elsayed, S.; Louie, T.; Conly, J.M. Novel multiplex PCR assay for simultaneous identification of community-associated methicillin-resistant Staphylococcus aureus strains USA300 and USA400 and detection of mecA and Panton-Valentine leukocidin genes, with discrimination of Staphylococcus aureus from coagulase-negative staphylococci. J. Clin. Microbiol. 2008, 46, 1118–1122. [Google Scholar]
- Zhang, D.F.; Xu, X.; Song, Q.; Bai, Y.; Zhang, Y.; Song, M.; Shi, C.; Shi, X. Identification of Staphylococcus argenteus in Eastern China based on a nonribosomal peptide synthetase (NRPS) gene. Future Microbiol. 2016, 11, 1113–1121. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; CLSI: Wayne, PA, USA, 2018; p. M100. Available online: http://iacld.ir/DL/public/CLSI-2018-M100-S28.pdf (accessed on 1 May 2021).
- The European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 11.0. 2021. Available online: http://www.eucast.org (accessed on 1 May 2021).
- Watanabe, A.; Yanagihara, K.; Matsumoto, T.; Kohno, S.; Aoki, N.; Oguri, T.; Sato, J.; Muratani, T.; Yagisawa, M.; Ogasawara, K.; et al. National surveillance of bacterial respiratory pathogens conducted by the Surveillance Committee of Japanese Society of Chemotherapy, Japanese Association for Infectious Diseases. J. Infect. Chemother. 2012, 18, 609–620. [Google Scholar] [CrossRef] [Green Version]
- Aung, M.S.; Urushibara, N.; Kawaguchiya, M.; Sumi, A.; Shinagawa, M.; Takahashi, S.; Kobayashi, N. Clonal Diversity and Genetic Characteristics of Methicillin-Resistant Staphylococcus aureus Isolates from a Tertiary Care Hospital in Japan. Microb. Drug Resist. 2019, 25, 1164–1175. [Google Scholar] [CrossRef]
- Roy, S.; Aung, M.S.; Paul, S.K.; Ahmed, S.; Haque, N.; Khan, E.R.; Barman, T.K.; Islam, A.; Abedin, S.; Sultana, C.; et al. Drug Resistance Determinants in Clinical Isolates of Enterococcus faecalis in Bangladesh: Identification of Oxazolidinone Resistance Gene optrA in ST59 and ST902 Lineages. Microorganisms 2020, 8, 1240. [Google Scholar] [CrossRef]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shopsin, B.; Gomez, M.; Montgomery, S.O.; Smith, D.H.; Waddington, M.; Dodge, D.E.; Bost, D.A.; Riehman, M.; Naidich, S.; Kreiswirth, B.N. Evaluation of protein A gene polymorphic region DNA sequencing for typing of Staphylococcus aureus strains. J. Clin. Microbiol. 1999, 37, 3556–3563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aung, M.S.; Urushibara, N.; Kawaguchiya, M.; Ito, M.; Habadera, S.; Kobayashi, N. Prevalence and Genetic Diversity of Staphylococcal Enterotoxin (-Like) Genes sey, selw, selx, selz, sel26 and sel27 in Community-Acquired Methicillin-Resistant Staphylococcus aureus. Toxins 2020, 12, 347. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Type of Ground Meat (No. of Samples) | No. of S. aureus Isolates (% in Each Type of Ground Meat) | CoNS *1/Mammaliicoccus Species | |
|---|---|---|---|
| No. of Isolates | Bacterial Species (No. of Isolates) | ||
| Chicken (93) | 3 (3.2%) | 61 | S. agnetis (1), S. capitis (8), S. carnosus (2), S. chromogenes (3), S. cohnii (5), S. condimenti (3), S. lugdunensis (1), S. pasteuri (8), S. saprophyticus (14), S. warneri (8), S. xylosus (1), M. sciuri (4), M. stepanovicii (3) |
| Pork (22) | 2 (9.1%) | 16 | S. agnetis (1), S. hyicus (1), S. pasteuri (1), S. rostri (1), S. saprophyticus (4), S. warneri (1), M. sciuri (6), M. vitulinus (1) |
| Beef and pork (31) | 5 (16.1%) *2 | 35 | S. chromogenes (3), S. haemolyticus (3), S. kloosii (1), S. pasteuri (2), S. rostri (1), S. saprophyticus (5), S. sciuri (10), S. simulans (1), S. warneri (5), M. lentus (1), M. vitulinus (3) |
| Total (146) | 10 (6.8%) | 112 | S. saprophyticus (23), M. sciuri (20), S. warneri (14), S. pasteuri (11), S. capitis (8), S. chromogenes (6), S. cohnii (5), other 13 species (25) |
| Isolate ID *1 | Specimen Type | coa Type | ST (CC) *2 | spa Type | Resistance Profile *3 | Resistance Gene *4 | Virulence Factor (Gene) Profile *5 |
|---|---|---|---|---|---|---|---|
| C26 | Chicken | IIa | ST5 (CC5) | t3478 | All susceptible | ND | lukDE, hla, hlb, hld, hlg, seg, sei, sem, sen, seo, selx, selw, eno, fib, sdrD |
| C35 | Chicken | IIa | ST5 (CC5) | t3478 | All susceptible | ND | lukDE, hla, hlb, hld, hlg, seg, sei, sem, sen, seo, selx, selw, eno, fib, sdrD |
| M3 | Pork and beef (mixed) | VIc | ST352 (CC97) | t5695 | All susceptible | ND | lukDE, lukM, hla, hlb, hld, hlg, selx, selw, fib, sdrD |
| M4-2 | Pork and beef (mixed) | VIIb | ST398 | t571 | AMP, LVX | blaZ | hla, hlb, hld, hlg, eno |
| P12-1 | Pork | VIIb | ST398 | t1419 | ERY, CLI-i | erm(C) | hla, hlb, hld, hlg, eno |
| M22-1 | Pork and beef (mixed) | VIIb | ST398 | t1419 | ERY, CLI-i | erm(C) | hla, hlb, hld, hlg, eno |
| C25-1 | Chicken | Xa | ST15 | t084 | AMP | blaZ | lukDE, hla, hlb, hld, hlg, seb, selx, selw, eno, fib, sdrD, scn |
| M20-1 | Pork and beef (mixed) | Xa | ST15 | t5819 | AMP | blaZ | lukDE, hla, hlb, hld, hlg, seb, selx, selw, eno, fib, sdrD, scn |
| M2 | Pork and beef (mixed) | XIc | ST9 (CC1) | t337 | AMP | blaZ | hla, hlb, hld, seg, sei, sem, sen, seo, selx, selw, sey, sel26, sel27, eno, fib, sdrD |
| P7-1 | Pork | XIc | ST9 (CC1) | t337 | AMP | blaZ | hla, hlb, hld, seg, sei, sem, sen, seo, selx, selw, sey, sel26, sel27, eno, fib, sdrD |
| Staphylococcus (Mammaliicoccus) Species | No. of Isolates | No. of Isolates Showing Resistance to Antimicrobials *1 (%) | No. of Isolates Having Resistance Genes *2 (%) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AMP | GEN | ERY | CLI | CLI-i | LVX | SXT | TET | blaZ | erm(C) | lnu(B) | aac(6′)-Ie-aph(2″)-Ia | ant(4′)-Ia | tet(K) | tet(M) | fosB | ||
| S. aureus | 10 | 5 (50) *3 | 0 (0) | 2 (20) | 0 (0) | 2 (20) | 1 (10) | 0 (0) | 0 (0) | 5 (50) *3 | 2 (20) | 0 | 0 | 0 | 0 | 0 | 0 |
| S. agnetis | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| S. capitis | 8 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 3 | 1 | 0 | 0 | 0 | 0 | 3 | 0 | 5 |
| S. carnosus | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S. chromogenes | 6 | 2 | 0 | 3 *4 | 4 *4 | 0 | 0 | 1 | 4 *4 | 2 | 3 *4 | 2 | 0 | 0 | 4 *4 | 1 | 0 |
| S. cohnii | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S. condimenti | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S. haemolyticus | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S. hyicus | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| S. kloosii | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S. lugdunensis | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 |
| S. pasteuri | 11 | 1 | 0 | 0 | 2 | 0 | 0 | 0 | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 2 | 2 |
| S. rostri | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 |
| S. saprophyticus | 23 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 2 | 1 | 13 *4 |
| S. simulans | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S. warneri | 14 | 13 *4 | 8 *4 | 1 | 1 | 1 | 0 | 1 | 3 | 10 *4 | 1 | 0 | 8 *4 | 8 *4 | 3 | 0 | 9 *4 |
| S. xylosus | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| M. lentus | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| M. sciuri | 20 | 2 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| M. stepanovicii | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| M. vitulinus | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| Total no. of CoNS/ Mammaliicoccus | 112 | 22 (19.6) | 9 (8.0) | 4 (3.6) | 9 (8.0) | 1 (0.9) | 0 (0) | 4 (3.6) | 21 (18.8) | 19 (17.0) | 4 (3.6) | 2 (1.8) | 9 (8.0) | 9 (8.0) | 16 (14.3) | 8 (7.1) | 30 (26.8) |
| Species | Isolate ID | Presence of tgs Genes | Other Staphylococcal Species Isolated from the Same Specimen |
|---|---|---|---|
| S. capitis | C1 | tgsB, C, D | - |
| C2 | tgsB, C, D | - | |
| C3 | tgsB, C, D | - | |
| C4 | - | - | |
| C5 | - | - | |
| C46-2 | - | S. warneri | |
| C47-2 | - | S. warneri, S. lugdunensis | |
| S. chromogenes | M22-2 | - | MSSA (ST398), S. scuiri |
| P12-4 | - | MSSA (ST398), S. pasteuri, S. warneri, S. haemolyticus | |
| C13-1 | tgsC, D | S. cohnii, M. stepanovicii | |
| C27-1 | tgsC, D | S. carnosus | |
| C36 | - | - | |
| C52-1 | - | - | |
| M1 | - | - |
| S. aureus Isolate ID | coa Type | ST (CC) | CoNS/Mammaliicoccus Isolated from the Same Specimen |
|---|---|---|---|
| C26 | IIa | ST5 (CC5) | - |
| C35 | IIa | ST5 (CC5) | - |
| M3 | VIc | ST352 (CC97) | - |
| M4-2 | VIIb | ST398 | M. sciuri |
| P12-1 | VIIb | ST398 | S. chromogenes, S. haemolyticus, S. pasteuri, S. warneri |
| M22-1 | VIIb | ST398 | S. chromogenes, M. sciuri |
| C25-1 | Xa | ST15 | S. saprophyticus |
| M20-1 | Xa | ST15 | S. pasteuri, S. warneri, M. vitulinus |
| M2 | XIc | ST9 (CC1) | - |
| P7-1 | XIc | ST9 (CC1) | S. rostri |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osada, M.; Aung, M.S.; Urushibara, N.; Kawaguchiya, M.; Ohashi, N.; Hirose, M.; Kobayashi, N. Prevalence and Antimicrobial Resistance of Staphylococcus aureus and Coagulase-Negative Staphylococcus/Mammaliicoccus from Retail Ground Meat: Identification of Broad Genetic Diversity in Fosfomycin Resistance Gene fosB. Pathogens 2022, 11, 469. https://doi.org/10.3390/pathogens11040469
Osada M, Aung MS, Urushibara N, Kawaguchiya M, Ohashi N, Hirose M, Kobayashi N. Prevalence and Antimicrobial Resistance of Staphylococcus aureus and Coagulase-Negative Staphylococcus/Mammaliicoccus from Retail Ground Meat: Identification of Broad Genetic Diversity in Fosfomycin Resistance Gene fosB. Pathogens. 2022; 11(4):469. https://doi.org/10.3390/pathogens11040469
Chicago/Turabian StyleOsada, Masako, Meiji Soe Aung, Noriko Urushibara, Mitsuyo Kawaguchiya, Nobuhide Ohashi, Mina Hirose, and Nobumichi Kobayashi. 2022. "Prevalence and Antimicrobial Resistance of Staphylococcus aureus and Coagulase-Negative Staphylococcus/Mammaliicoccus from Retail Ground Meat: Identification of Broad Genetic Diversity in Fosfomycin Resistance Gene fosB" Pathogens 11, no. 4: 469. https://doi.org/10.3390/pathogens11040469