
Prevalence of Tick-Borne Pathogens in Questing Ixodes ricinus and Dermacentor reticulatus Ticks Collected from Recreational Areas in Northeastern Poland with Analysis of Environmental Factors

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Ticks
2.2. DNA Isolation
2.3. PCR Amplification
2.4. Evolutionary Relationships of Taxa
- 118 nucleotide sequences for Borrelia isolated from I. ricinus in Białystok (1264 bp),
- 94 nucleotide sequences for Borrelia isolated from I. ricinus in Augustów (1243 bp),
- 17 nucleotide sequences for Babesia isolated from I. ricinus (289 bp),
- 19 nucleotide sequences for Babesia isolated from D. reticulatus (275 bp).
2.5. Statistical Analysis of Previous and Present Research
3. Results
3.1. Sequencing Analysis
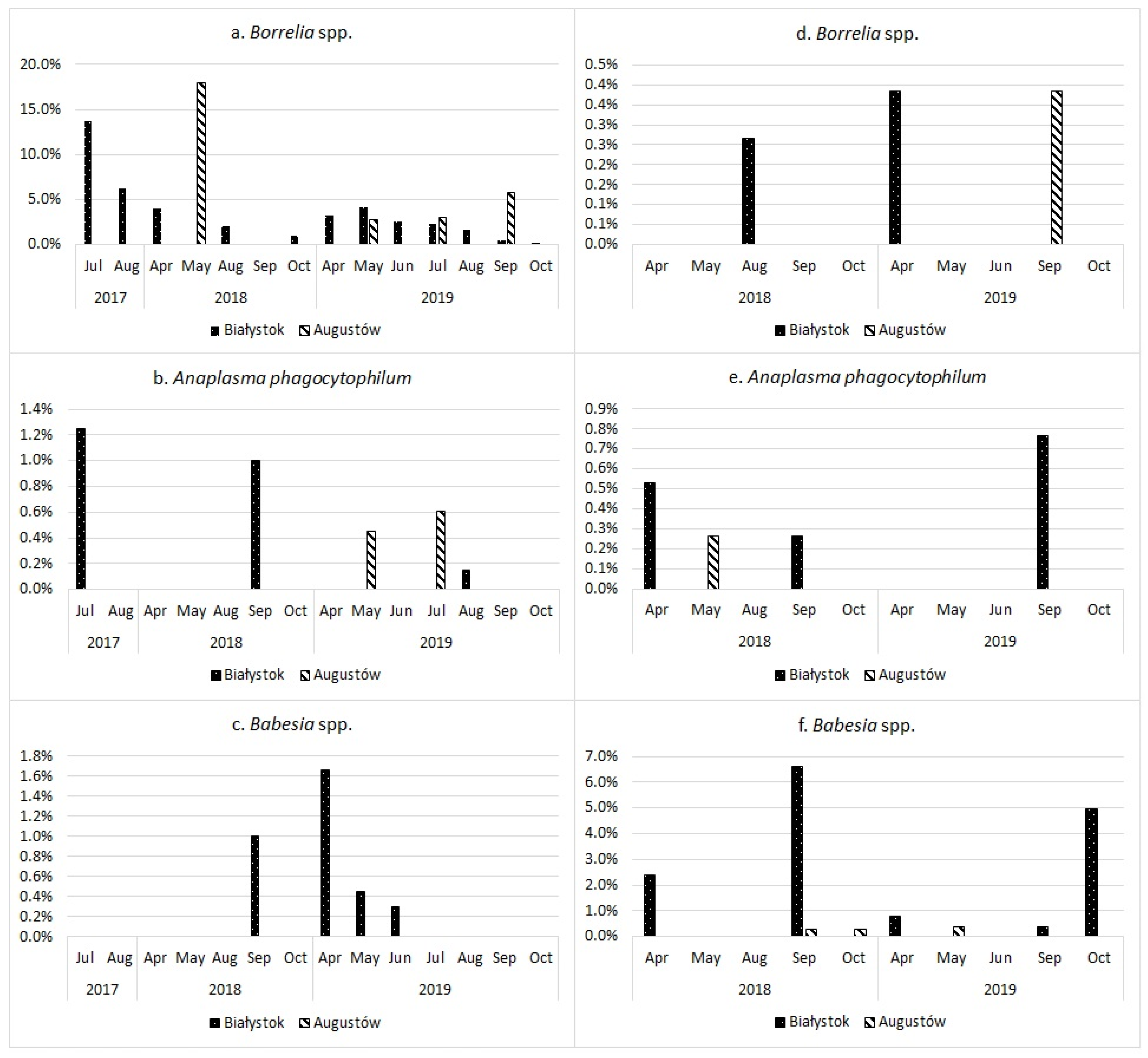
3.2. Co-Infections
3.3. Phylogenetic Analysis
3.4. Statistical Analysis
3.4.1. Mann-Whitney Test
Overall Infection Rate
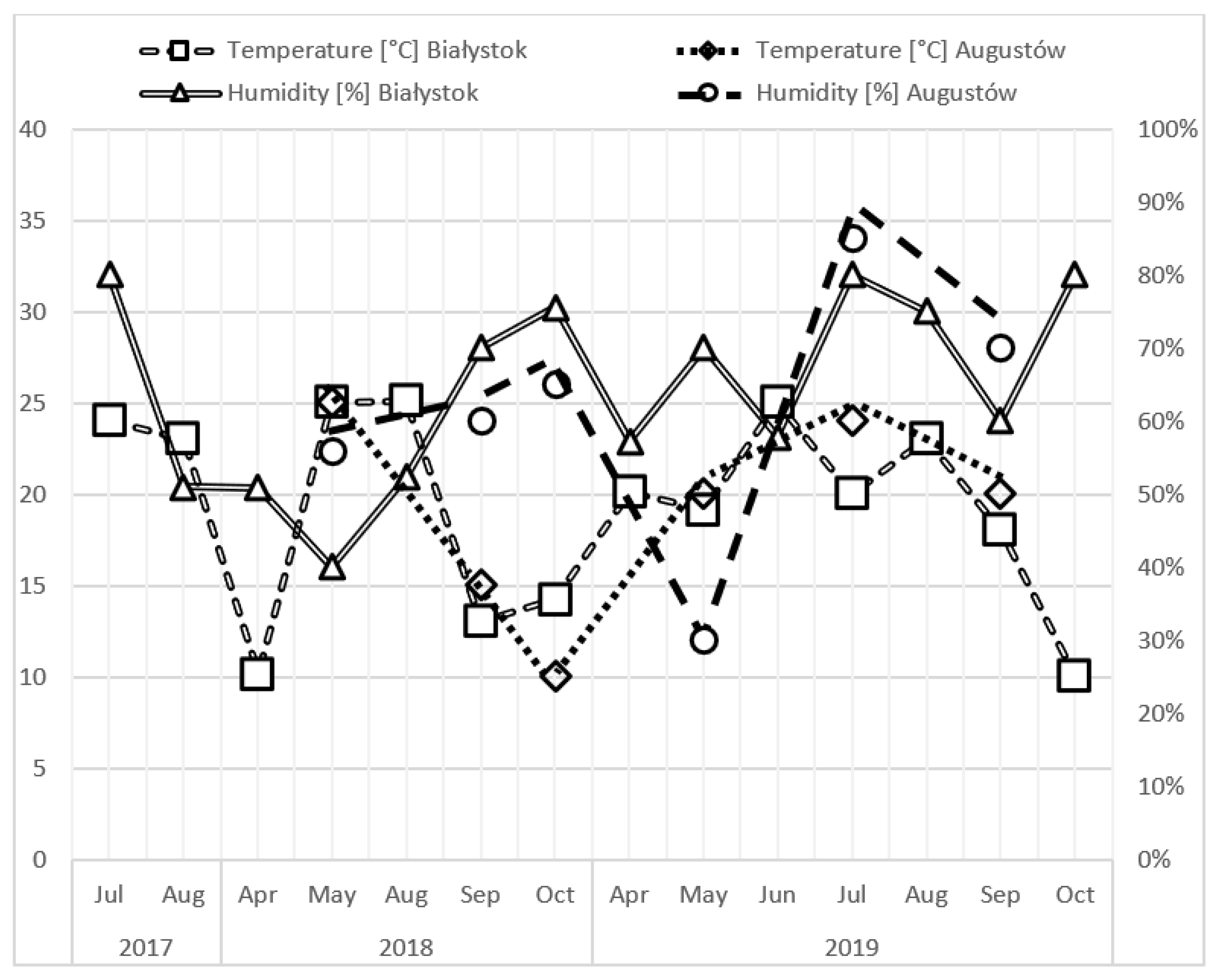
Air Temperature
Relative Air Humidity
Sampling Season
3.4.2. Multivariate Logistic Regression Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubalek, Z.; Foldvari, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Spitalska, E.; et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: New Hazards and Relevance for Public Health. Front. Public Health 2014, 2, 251. [Google Scholar] [CrossRef] [PubMed]
- Akimov, I.A.; Nebogatkin, I.V. Ixodid ticks (Acari, Ixodidae) in urban landscapes. A Review. Vestn. Zool. 2016, 50, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Heylen, D.; Lasters, R.; Adriaensen, F.; Fonville, M.; Sprong, H.; Matthysen, E. Ticks and tick-borne diseases in the city: Role of Landscape Connectivity and Green Space Characteristics in a Metropolitan Area. Sci. Total Environ. 2019, 670, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Venclíková, K.; Mendel, J.; Betášová, L.; Blažejová, H.; Jedličková, P.; Straková, P.; Hubálek, Z.; Rudolf, I. Neglected tick-borne pathogens in the Czech Republic, 2011–2014. Ticks Tick. Borne. Dis. 2016, 7, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Didyk, Y.M.; Blaňárová, L.; Pogrebnyak, S.; Akimov, I.; Peťko, B.; Víchová, B. Emergence of tick-borne pathogens (Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum, Rickettsia raoultii and Babesia microti) in the Kyiv urban parks, Ukraine. Ticks Tick. Borne. Dis. 2017, 8, 219–225. [Google Scholar] [CrossRef]
- Kybicová, K.; Baštová, K.; Malý, M. Detection of Borrelia burgdorferi sensu lato and Anaplasma phagocytophilum in questing ticks Ixodes ricinus from the Czech Republic. Ticks Tick. Borne. Dis. 2017, 8, 483–487. [Google Scholar] [CrossRef]
- Lommano, E.; Bertaiola, L.; Dupasquier, C.; Gern, L. Infections and coinfections of questing Ixodes ricinus ticks by emerging zoonotic pathogens in Western Switzerland. Appl. Environ. Microbiol. 2012, 78, 4606–4612. [Google Scholar] [CrossRef] [Green Version]
- Oechslin, C.P.; Heutschi, D.; Lenz, N.; Tischhauser, W.; Péter, O.; Rais, O.; Beuret, C.M.; Leib, S.L.; Bankoul, S.; Ackermann-Gäumann, R. Prevalence of tick-borne pathogens in questing Ixodes ricinus ticks in urban and suburban areas of Switzerland. Parasites Vectors 2017, 10, 558. [Google Scholar] [CrossRef] [Green Version]
- Kowalec, M.; Szewczyk, T.; Welc-Falȩciak, R.; Siński, E.; Karbowiak, G.; Bajer, A. Ticks and the city—Are there any differences between city parks and natural forests in terms of tick abundance and prevalence of spirochaetes? Parasites Vectors 2017, 10, 573. [Google Scholar] [CrossRef]
- Blazejak, K.; Janecek, E.; Strube, C. A 10-Year Surveillance of Rickettsiales (Rickettsia spp. and Anaplasma phagocytophilum) in the city of Hanover, Germany, reveals Rickettsia spp. as emerging pathogens in ticks. Parasites Vectors 2017, 10, 588. [Google Scholar] [CrossRef] [Green Version]
- Welc-Falȩciak, R.; Kowalec, M.; Karbowiak, G.; Bajer, A.; Behnke, J.M.; Siński, E. Rickettsiaceae and Anaplasmataceae infections in Ixodes ricinus ticks from urban and natural forested areas of Poland. Parasites Vectors 2014, 7, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada-Peña, A.; Ayllón, N.; de la Fuente, J. Impact of climate trends on tick-borne pathogen transmission. Front. Physiol. 2012, 3, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogden, N.H.; Lindsay, L.R. Effects of climate and climate change on vectors and vector-borne diseases: Ticks are Different. Trends Parasitol. 2016, 32, 646–656. [Google Scholar] [CrossRef]
- National Institute of Public Health. Infectious Diseases and Poisonings in Poland in 2019. Available online: http://wwwold.pzh.gov.pl/oldpage/epimeld/2019/Ch_2019.pdf (accessed on 13 October 2021).
- Grochowska, A.; Dunaj, J.; Pancewicz, S.; Czupryna, P.; Majewski, P.; Wondim, M.; Tryniszewska, E.; Moniuszko-Malinowska, A. Detection of Borrelia burgdorferi s.l., Anaplasma phagocytophilum and Babesia spp. in Dermacentor reticulatus ticks found within the city of Białystok, Poland-first data. Exp. Appl. Acarol. 2021, 85, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Chmura, M. Fauna Kleszczy (Ixodida) Europy Środkowej; Wydawnictwo Naukowe Uniwersytetu Pedagogicznego: Kraków, Poland, 2013; ISBN 978-83-7271-829-7. [Google Scholar]
- Dunaj, J.; Moniuszko-Malinowska, A.; Święcicka, I.; Andersson, M.; Czupryna, P.; Rutkowski, K.; Zambrowski, G.; Zajkowska, J.; Grygorczuk, S.; Kondrusik, M.; et al. Tick-borne infections and co-infections in patients with non-specific symptoms in Poland. Adv. Med. Sci. 2018, 63, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Moniuszko-Malinowska, A.; Święcicka, I.; Dunaj, J.; Zajkowska, J.; Czupryna, P.; Zambrowski, G.; Chmielewska–Badora, J.; Żukiewicz-Sobczak, W.; Swierzbinska, R.; Rutkowski, K.; et al. Infection with Babesia microti in humans with non-specific symptoms in north east Poland. Infect. Dis. 2016, 48, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Katargina, O.; Geller, J.; Vasilenko, V.; Kuznetsova, T.; Järvekülg, L.; Vene, S.; Lundkvist, Ã.; Golovljova, I. Detection and characterization of Babesia species in Ixodes ticks in Estonia. Vector-Borne Zoonotic Dis. 2011, 11, 923–928. [Google Scholar] [CrossRef]
- Pichon, B.; Kahl, O.; Hammer, B.; Gray, J.S. Pathogens and host DNA in Ixodes ricinus nymphal ticks from a German Forest. Vector Borne Zoonotic Dis. 2006, 6, 382–387. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Reye, A.L.; Hübschen, J.M.; Sausy, A.; Muller, C.P. Prevalence and Seasonality of Tick-Borne Pathogens in Questing Ixodes ricinus ticks from Luxembourg. Appl. Environ. Microbiol. 2010, 76, 2923–2931. [Google Scholar] [CrossRef] [Green Version]
- Kiewra, D.; Stańczak, J.; Richter, M. Ixodes ricinus ticks (Acari, Ixodidae) as a vector of Borrelia burgdorferi sensu lato and Borrelia miyamotoi in Lower Silesia, Poland—Preliminary study. Ticks Tick. Borne. Dis. 2014, 5, 892–897. [Google Scholar] [CrossRef] [PubMed]
- May, K.; Jordan, D.; Fingerle, V.; Strube, C. Borrelia burgdorferi sensu lato and co-infections with Anaplasma phagocytophilum and Rickettsia spp. in Ixodes ricinus in Hamburg, Germany. Med. Vet. Entomol. 2015, 29, 425–429. [Google Scholar] [CrossRef]
- Pangrácová, L.; Derdáková, M.; Pekárik, L.; Hviščová, I.; Víchová, B.; Stanko, M.; Hlavatá, H.; Peťko, B. Ixodes ricinus abundance and its infection with the tick-borne pathogens in urban and suburban areas of eastern Slovakia. Parasites Vectors 2013, 6, 238. [Google Scholar] [CrossRef] [Green Version]
- Krause, P.J.; Narasimhan, S.; Wormser, G.P.; Rollend, L.; Fikrig, E.; Lepore, T.; Barbour, A.; Fish, D. Human Borrelia Miyamotoi Infection in the United States. N. Engl. J. Med. 2013, 368, 291–293. [Google Scholar] [CrossRef] [Green Version]
- Platonov, A.E.; Karan, L.S.; Kolyasnikova, N.M.; Makhneva, N.A.; Toporkova, M.G.; Maleev, V.V.; Fish, D.; Krause, P.J. Humans infected with relapsing fever spirochete Borrelia miyamotoi, Russia. Emerg. Infect. Dis. 2011, 17, 1816–1823. [Google Scholar] [CrossRef]
- Kubiak, K.; Szczotko, M.; Dmitryjuk, M. Borrelia miyamotoi—an emerging human tick-borne pathogen in Europe. Microorganisms 2021, 9, 154. [Google Scholar] [CrossRef]
- Sato, K.; Takano, A.; Konnai, S.; Nakao, M.; Ito, T.; Koyama, K.; Kaneko, M.; Ohnishi, M.; Kawabata, H. Human Infections with Borrelia miyamotoi, Japan. Emerg. Infect. Dis. 2014, 20, 1391–1393. [Google Scholar] [CrossRef]
- Strnad, M.; Honig, V.; Ruzek, D.; Grubhoffer, L.; Rego, R.O.M. Europe-wide meta-analysis of Borrelia burgdorferi sensu lato prevalence in questing Ixodes ricinus ticks. Appl. Environ. Microbiol. 2017, 83, e00609-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mierzejewska, E.J.; Pawełczyk, A.; Radkowski, M.; Welc-Falęciak, R.; Bajer, A. Pathogens vectored by the tick, Dermacentor reticulatus, in endemic regions and zones of expansion in Poland. Parasites Vectors 2015, 8, 490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzięgiel, B.; Kubrak, T.; Adaszek, Ł.; Dębiak, P.; Wyłupek, D.; Bogucka-kocka, A.; Lechowski, J.; Winiarczyk, S. Prevalence of Babesia canis, Borrelia burgdorferi sensu lato, and Anaplasma phagocytophilum in hard ticks collected from meadows of Lubelskie Voivodship (Eastern Poland). Bull. Vet. Inst. Pulawy 2014, 58, 29–33. [Google Scholar] [CrossRef] [Green Version]
- Zając, V.; Wójcik-Fatla, A.; Sawczyn, A.; Cisak, E.; Sroka, J.; Kloc, A.; Zając, Z.; Buczek, A.; Dutkiewicz, J.; Bartosik, K. Prevalence of infections and co-infections with 6 pathogens in Dermacentor reticulatus ticks collected in eastern Poland. Ann. Agric. Environ. Med. 2017, 24, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Król, N.; Kiewra, D.; Szymanowski, M.; Lonc, E. The role of domestic dogs and cats in the zoonotic cycles of ticks and pathogens. preliminary studies in the Wroclaw Agglomeration (SW Poland). Vet. Parasitol. 2015, 214, 208–212. [Google Scholar] [CrossRef]
- Reye, A.L.; Stegniy, V.; Mishaeva, N.P.; Velhin, S.; Hübschen, J.M.; Ignatyev, G.; Muller, C.P. Prevalence of tick-borne pathogens in Ixodes ricinus and Dermacentor reticulatus ticks from different geographical locations in Belarus. PLoS ONE 2013, 8, e54476. [Google Scholar] [CrossRef] [Green Version]
- Richter, D.; Kohn, C.; Matuschka, F.-R. Absence of Borrelia spp., Candidatus Neoehrlichia mikurensis, and Anaplasma phagocytophilum in questing adult Dermacentor reticulatus ticks. Parasitol. Res. 2013, 112, 107–111. [Google Scholar] [CrossRef]
- Tomanovic, S.; Chochlakis, D.; Radulovic, Z.; Milutinovic, M.; Cakic, S.; Mihaljica, D.; Tselentis, Y.; Psaroulaki, A. Analysis of pathogen co-occurrence in host-seeking adult hard ticks from Serbia. Exp. Appl. Acarol. 2013, 59, 367–376. [Google Scholar] [CrossRef]
- Rudolf, I.; Hubalek, Z. Effect of the salivary gland and midgut extracts from Ixodes ricinus and Dermacentor reticulatus (Acari: Ixodidae) on the growth of Borrelia garinii in vitro. Folia Parasitol. 2003, 50, 159–160. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.S.; Dautel, H.; Estrada-Peña, A.; Kahl, O.; Lindgren, E. Effects of climate change on ticks and tick-borne diseases in Europe. Interdiscip. Perspect. Infect. Dis. 2009, 2009, 593232. [Google Scholar] [CrossRef]
- Steinbrink, A.; Brugger, K.; Margos, G.; Kraiczy, P.; Klimpel, S. The evolving story of Borrelia burgdorferi sensu lato transmission in Europe. Parasitol. Res. 2022, 121, 781–803. [Google Scholar] [CrossRef] [PubMed]
- Grochowska, A.; Milewski, R.; Pancewicz, S.; Dunaj, J.; Czupryna, P.; Milewska, A.J.; Róg-Makal, M.; Grygorczuk, S.; Moniuszko-Malinowska, A. Comparison of tick-borne pathogen prevalence in Ixodes ricinus ticks collected in urban areas of Europe. Sci. Rep. 2020, 10, 6975. [Google Scholar] [CrossRef] [PubMed]
- Keith, K.; Årestedt, K.; Tjernberg, I. The relationship between the laboratory diagnosis of Lyme neuroborreliosis and climate factors in Kalmar County Sweden—An overview between 2008 and 2019. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Bennet, L.; Halling, A.; Berglund, J. Increased incidence of Lyme borreliosis in southern Sweden following mild winters and during warm, humid summers. Eur. J. Clin. Microbiol. Infect. Dis. 2006, 25, 426–432. [Google Scholar] [CrossRef] [Green Version]
- Földvári, G.; Široký, P.; Szekeres, S.; Majoros, G.; Sprong, H. Dermacentor reticulatus: A Vector on the Rise. Parasites Vectors 2016, 9, 314. [Google Scholar] [CrossRef] [Green Version]
- Tokhov, Y.M.; Lutsuk, S.N.; Dyachenko, Y.V. Phenology of Ixodid ticks of the genus Dermacentor in the Central Ciscaucasia. Entomol. Rev. 2014, 94, 426–433. [Google Scholar] [CrossRef]
- Stańczak, J.; Cieniuch, S.; Lass, A.; Biernat, B.; Racewicz, M. Detection and quantification of Anaplasma phagocytophilum and Babesia spp. in Ixodes ricinus ticks from urban and rural environment, northern Poland, by Real-Time Polymerase Chain Reaction. Exp. Appl. Acarol. 2015, 66, 63–81. [Google Scholar] [CrossRef] [Green Version]
- Mehlhorn, H.; Mehlhorn, T.; Müller, M.; Vogt, M.; Rissland, J. Tick survey for prevalent pathogens in peri-urban recreation sites in Saarland and Rhineland-Palatinate (Germany). Parasitol. Res. 2016, 115, 1167–1172. [Google Scholar] [CrossRef]
- Svitálková, Z.; Haruštiaková, D.; Mahríková, L.; Berthová, L.; Slovák, M.; Kocianová, E.; Kazimírová, M. Anaplasma phagocytophilum prevalence in ticks and rodents in an urban and natural habitat in south-western Slovakia. Parasites Vectors 2015, 8, 276. [Google Scholar] [CrossRef] [Green Version]
- Derdáková, M.; Václav, R.; Pangrácova-Blaňárová, L.; Selyemová, D.; Koči, J.; Walder, G.; Špitalská, E. Candidatus Neoehrlichia mikurensis and its co-circulation with Anaplasma Phagocytophilum in Ixodes ricinus ticks across ecologically different habitats of Central Europe. Parasites Vectors 2014, 7, 160. [Google Scholar] [CrossRef] [Green Version]
- Rogovskyy, A.; Batool, M.; Gillis, D.C.; Holman, P.J.; Nebogatkin, I.V.; Rogovska, Y.V.; Rogovskyy, M.S. Diversity of Borrelia spirochetes and other zoonotic agents in ticks from Kyiv, Ukraine. Ticks Tick. Borne. Dis. 2018, 9, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Opalińska, P.; Wierzbicka, A.; Asman, M. The PCR and nested PCR detection of Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum and Babesia microti in Dermacentor reticulatus F. Collected in a new location in Poland (Trzciel, Western Poland). Acta Parasitol. 2016, 61, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Huhn, C.; Winter, C.; Wolfsperger, T.; Wuppenhorst, N.; Strasek Smrdel, K.; Skuballa, J.; Pfaffle, M.; Petney, T.; Silaghi, C.; Dyachenko, V.; et al. Analysis of the population structure of Anaplasma phagocytophilum using multilocus sequence typing. PLoS ONE 2014, 9, e93725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wójcik-Fatla, A.; Zając, V.; Sawczyn, A.; Cisak, E.; Dutkiewicz, J. Babesia spp. in questing ticks from eastern Poland: Prevalence and Species Diversity. Parasitol. Res. 2015, 114, 3111–3116. [Google Scholar] [CrossRef] [Green Version]
- Hamšíková, Z.; Kazimírová, M.; Haruštiaková, D.; Mahríková, L.; Slovák, M.; Berthová, L.; Kocianová, E.; Schnittger, L. Babesia spp. in ticks and wildlife in different habitat types of Slovakia. Parasites Vectors 2016, 9, 292. [Google Scholar] [CrossRef] [Green Version]
- Obiegala, A.; Pfeffer, M.; Pfister, K.; Karnath, C.; Silaghi, C. Molecular examinations of Babesia microti in rodents and rodent-attached ticks from urban and sylvatic habitats in Germany. Ticks Tick. Borne. Dis. 2015, 6, 445–449. [Google Scholar] [CrossRef]
- Karbowiak, G. The occurrence of the Dermacentor reticulatus tick--its expansion to new areas and possible causes. Ann. Parasitol. 2014, 60, 37–47. [Google Scholar]
- Dwużnik-Szarek, D.; Mierzejewska, E.J.; Rodo, A.; Goździk, K.; Behnke-Borowczyk, J.; Kiewra, D.; Kartawik, N.; Bajer, A. Monitoring the expansion of Dermacentor reticulatus and occurrence of canine babesiosis in Poland in 2016–2018. Parasit. Vectors 2021, 14, 267. [Google Scholar] [CrossRef]
- Karbowiak, G.; Vichová, B.; Slivinska, K.; Werszko, J.; Didyk, J.; Peťko, B.; Stanko, M.; Akimov, I. The infection of questing Dermacentor reticulatus ticks with Babesia canis and Anaplasma phagocytophilum in the Chernobyl Exclusion Zone. Vet. Parasitol. 2014, 204, 372–375. [Google Scholar] [CrossRef]
- Svehlová, A.; Berthová, L.; Sallay, B.; Boldiš, V.; Sparagano, O.A.E.; Spitalská, E. Sympatric occurrence of Ixodes ricinus, Dermacentor reticulatus and Haemaphysalis concinna ticks and Rickettsia and Babesia species in Slovakia. Ticks Tick. Borne. Dis. 2014, 5, 600–605. [Google Scholar] [CrossRef]
- Zając, V.; Wójcik-Fatla, A.; Dutkiewicz, J.; Szymańska, J. Bartonella henselae in eastern Poland: The Relationship between Tick Infection Rates and the Serological Response of Individuals Occupationally Exposed to Tick Bites. J. Vector Ecol. 2015, 40, 75–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wójcik-Fatla, A.; Bartosik, K.; Buczek, A.; Dutkiewicz, J. Babesia microti in adult Dermacentor reticulatus ticks from eastern Poland. Vector Borne Zoonotic Dis. 2012, 12, 841–843. [Google Scholar] [CrossRef] [PubMed]
- Dautel, H.; Dippel, C.; Oehme, R.; Hartelt, K.; Schettler, E. Evidence for an increased geographical distribution of Dermacentor reticulatus in Germany and detection of Rickettsia Sp. RpA4. Int. J. Med. Microbiol. 2006, 296, 149–156. [Google Scholar] [CrossRef]
- Chmielewski, T.; Andrzejewski, K.; Maczka, I.; Fiecek, B.; Radlinska, M.; Tylewska-Wierzbanowska, S. Ticks infected with bacteria pathogenic to humans in municipal parks in Warsaw. Przegl. Epidemiol. 2011, 65, 577–581. [Google Scholar] [PubMed]
- Tylewska-Wierzbanowska, S.; Kruszewska, D.; Chmielewski, T.; Zukowski, K.; Zabicka, J. Ticks as a reservoir of Borrelia burgdorferi and Coxiella burnetii on Polish terrain. Przegl. Epidemiol. 1996, 50, 245–251. [Google Scholar] [PubMed]
- Pilloux, L.; Baumgartner, A.; Jaton, K.; Lienhard, R.; Ackermann-Gäumann, R.; Beuret, C.; Greub, G. Prevalence of Anaplasma phagocytophilum and Coxiella burnetii in Ixodes ricinus ticks in Switzerland: An Underestimated Epidemiologic Risk. New Microbes New Infect. 2019, 27, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Schötta, A.M.; Wijnveld, M.; Stockinger, H.; Stanek, G. Approaches for reverse line blot-based detection of microbial pathogens in Ixodes ricinus ticks collected in Austria and impact of the chosen method. Appl. Environ. Microbiol. 2017, 83, e00489-17. [Google Scholar] [CrossRef] [Green Version]
- Wallménius, K.; Pettersson, J.H.O.; Jaenson, T.G.T.; Nilsson, K. Prevalence of Rickettsia spp., Anaplasma phagocytophilum, and Coxiella burnetii in adult Ixodes ricinus ticks from 29 study areas in central and southern Sweden. Ticks Tick. Borne. Dis. 2012, 3, 100–106. [Google Scholar] [CrossRef]
- Szymańska-Czerwińska, M.; Monika Galińska, E.; Niemczuk, K.; Zasepa, M. Prevalence of Coxiella burnetii infection in foresters and ticks in south-eastern Poland and comparison of diagnostic methods. Ann. Agric. Environ. Med. 2013, 20, 699–704. [Google Scholar]
- Smetanová, K.; Schwarzová, K.; Kocianová, E. Detection of Anaplasma phagocytophilum, Coxiella burnetii, Rickettsia spp., and Borrelia burgdorferi s. l. in ticks, and wild-living animals in western and middle Slovakia. Ann. N. Y. Acad. Sci. 2006, 1078, 312–315. [Google Scholar] [CrossRef]
- Minichová, L.; Hamšíková, Z.; Mahríková, L.; Slovák, M.; Kocianová, E.; Kazimírová, M.; Škultéty, Ľ.; Štefanidesová, K.; Špitalská, E. Molecular Evidence of Rickettsia spp. in Ixodid ticks and rodents in suburban, natural and rural habitats in Slovakia. Parasites Vectors 2017, 10, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Špitalská, E.; Sparagano, O.; Stanko, M.; Schwarzová, K.; Špitalský, Z.; Škultéty, Ľ.; Havlíková, S.F. Diversity of Coxiella-like and Francisella-like endosymbionts, and Rickettsia spp., Coxiella burnetii as pathogens in the tick populations of Slovakia, Central Europe. Ticks Tick. Borne. Dis. 2018, 9, 1207–1211. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, A.; Franke, J.; Schmoock, G.; Pauliks, K.; Krämer, A.; Straube, E. Diversity and coexistence of tick-borne pathogens in central Germany. J. Med. Entomol. 2011, 48, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Pluta, S.; Hartelt, K.; Oehme, R.; Mackenstedt, U.; Kimmig, P. Prevalence of Coxiella burnetii and Rickettsia spp. in ticks and rodents in southern Germany. Ticks Tick. Borne. Dis. 2010, 1, 145–147. [Google Scholar] [CrossRef]
- Mancini, F.; Di Luca, M.; Toma, L.; Vescio, F.; Bianchi, R.; Khoury, C.; Marini, L.; Rezza, G.; Ciervo, A. Prevalence of tick-borne pathogens in an urban park in Rome, Italy. Ann. Agric. Environ. Med. 2014, 21, 723–727. [Google Scholar] [CrossRef] [Green Version]
- Schorn, S.; Pfister, K.; Reulen, H.; Mahling, M.; Silaghi, C. Occurrence of Babesia spp., Rickettsia spp. and Bartonella spp. in Ixodes ricinus in Bavarian public parks, Germany. Parasites Vectors 2011, 4, 135. [Google Scholar] [CrossRef] [Green Version]
- Raileanu, C.; Moutailler, S.; Pavel, I.; Porea, D.; Mihalca, A.D.; Savuta, G.; Vayssier-Taussat, M. Borrelia diversity and co-infection with other tick borne pathogens in ticks. Front. Cell. Infect. Microbiol. 2017, 7, 36. [Google Scholar] [CrossRef] [Green Version]
- Podsiadly, E.; Karbowiak, G.; Tylewska-Wierzbanowska, S. Presence of Bartonella spp. in Ixodidae Ticks. Clin. Microbiol. Infect. 2009, 15, 120–121. [Google Scholar] [CrossRef] [Green Version]
- Sytykiewicz, H.; Karbowiak, G.; Werszko, J.; Czerniewicz, P.; Sprawka, I.; Mitrus, J. Molecular screening for Bartonella henselae and Borrelia burgdorferi sensu lato co-existence within Ixodes ricinus populations in central and eastern parts of Poland. Ann. Agric. Environ. Med. 2012, 19, 451–456. [Google Scholar]
- Špitalská, E.; Boldiš, V.; Derdáková, M.; Selyemová, D.; Rusňáková Tarageľová, V. Rickettsial infection in Ixodes ricinus ticks in urban and natural habitats of Slovakia. Ticks Tick. Borne. Dis. 2014, 5, 161–165. [Google Scholar] [CrossRef]
- Szekeres, S.; Docters van Leeuwen, A.; Rigó, K.; Jablonszky, M.; Majoros, G.; Sprong, H.; Földvári, G. Prevalence and diversity of human pathogenic Rickettsiae in urban versus rural habitats, Hungary. Exp. Appl. Acarol. 2016, 68, 223–226. [Google Scholar] [CrossRef] [PubMed]
- May, K.; Strube, C. Prevalence of Rickettsiales (Anaplasma phagocytophilum and Rickettsia spp.) in hard ticks (Ixodes ricinus) in the city of Hamburg, Germany. Parasitol. Res. 2014, 113, 2169–2175. [Google Scholar] [CrossRef] [PubMed]
- Stańczak, J. Detection of Spotted Fever Group (SFG) Rickettsiae in Dermacentor reticulatus (Acari: Ixodidae) in Poland. Int. J. Med. Microbiol. 2006, 296, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Stańczak, J.; Biernat, B.; Racewicz, M.; Zalewska, M.; Matyjasek, A. Prevalence of different Rickettsia spp. in Ixodes ricinus and Dermacentor reticulatus ticks (Acari: Ixodidae) in north-eastern Poland. Ticks Tick. Borne. Dis. 2018, 9, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Wójcik-Fatla, A.; Cisak, E.; Zajac, V.; Sroka, J.; Sawczyn, A.; Dutkiewicz, J. Study on tick-borne Rickettsiae in eastern Poland. I. Prevalence in Dermacentor reticulatus (Acari: Amblyommidae). Ann. Agric. Environ. Med. 2013, 20, 276–279. [Google Scholar]
- Nijhof, A.M.; Bodaan, C.; Postigo, M.; Nieuwenhuijs, H.; Opsteegh, M.; Franssen, L.; Jebbink, F.; Jongejan, F. Ticks and associated pathogens collected from domestic animals in the Netherlands. Vector-Borne Zoonotic Dis. 2007, 7, 585–595. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ixodes ricinus | Dermacentor reticulatus | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Collection Site | Sampling Year | Females | Males | Nymphs | Larvae | Total | Females | Males | Nymphs | Total |
| Białystok | 2017 | 33 | 41 | 6 | - | 80 | - | - | - | - |
| 2018 | 16 | 14 | 2 | - | 32 | - | - | - | - | |
| 2019 | 127 | 92 | 79 | 50 | 348 | 150 | 94 | 8 | 252 | |
| Total | 176 | 147 | 87 | 50 | 460 | 150 | 94 | 8 | 252 | |
| Augustów | ||||||||||
| “Królowa Woda” resort | 2018 | 18 | 15 | 35 | - | 68 | 5 | 4 | - | 9 |
| Public Beach | - | - | - | - | - | - | - | - | - | |
| “Królowa Woda” resort | 2019 | 3 | 14 | 14 | - | 31 | 1 | - | - | 1 |
| Public Beach | 42 | 31 | 183 | 27 | 283 | 6 | 2 | - | 8 | |
| Total | 63 | 60 | 232 | 27 | 382 | 12 | 6 | - | 18 | |
| Ixodes ricinus | Dermacentor reticulatus | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Collection Site | Sampling Year | Bor | Bab | Ap | Total | Bor | Bab | Ap | Total |
| Białystok | 2017 | 20% (16/80) | 0% (0/80) | 1.3% (1/80) | 21.3% (17/80) | - | - | - | - |
| 2018 | 21.9% (7/32) | 3.1% (1/32) | 3.1% (1/32) | 28.1% (9/32) | - | - | - | - | |
| 2019 | 27.3% (95/348) | 4.6% (16/348) | 0.3% (1/348) | 28.4% (99/348) | 0.4% (1/252) | 6.4% (16/252) | 0.8% (2/252) | 7.6% (19/252) | |
| Total | 25.7% (118/460) | 3.7% (17/460) | 0.7% (3/460) | 27.2% (125/460) | 0.4% (1/252) | 6.4% (16/252) | 0.8% (2/252) | 7.6% (19/252) | |
| Augustów | |||||||||
| “Królowa Woda” resort | 2018 | 26.5% (18/68) | 0% (0/68) | 0% (0/68) | 26.5% (18/68) | 0% (0/9) | 22.2% (2/9) | 11.1% (1/9) | 33.3% (3/9) |
| “Królowa Woda” resort | 2019 | 12.9% (4/31) | 0% (0/31) | 3.2% (1/31) | 16.1% (5/31) | 0% (0/1) | 0% (0/1) | 0% (0/1) | 0% (0/1) |
| Public Beach | 25.4% (72/283) | 0% (0/283) | 2.1% (6/283) | 27.0% (77/283) | 12.5% (1/8) | 12.5% (1/8) | 0% (0/8) | 25% (2/8) | |
| Total | 24.6% (94/382) | 0% (0/382) | 1.8% (7/382) | 26.2% (100/382) | 5.5% (1/18) | 16.7% (3/18) | 5.5% (1/18) | 27.7% (5/18) | |
| Category | Variable | p Value |
|---|---|---|
| Overall infection rate between Dermacentor reticulatus and Ixodes ricinus | Borrelia spp. | 0.001 |
| Babesia spp. | 0.001 | |
| Temperature in April-July | Borrelia spp. | 0.003 |
| Babesia spp. | 0.001 | |
| Temperature in August-October | Borrelia spp. | 0.001 |
| Babesia spp. | 0.001 | |
| Relative air humidity in April-July | Babesia spp. | 0.001 |
| Relative air humidity in August-October | Borrelia spp. | 0.001 |
| Sampling season (April-July, August-October) | Borrelia spp. in females | 0.001 |
| Borrelia spp. in males | 0.001 | |
| Borrelia spp. in nymphs | 0.046 |
| Parameter | Odds Ratio | p Value | 95% Confidence Interval | |
|---|---|---|---|---|
| Tick species | 0.01 | 0.001 | 0.003 | 0.033 |
| Sampling year | 1.35 | 0.024 | 1.04 | 1.76 |
| Developmental stage | 0.63 | 0.001 | 0.53 | 0.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grochowska, A.; Dunaj-Małyszko, J.; Pancewicz, S.; Czupryna, P.; Milewski, R.; Majewski, P.; Moniuszko-Malinowska, A. Prevalence of Tick-Borne Pathogens in Questing Ixodes ricinus and Dermacentor reticulatus Ticks Collected from Recreational Areas in Northeastern Poland with Analysis of Environmental Factors. Pathogens 2022, 11, 468. https://doi.org/10.3390/pathogens11040468
Grochowska A, Dunaj-Małyszko J, Pancewicz S, Czupryna P, Milewski R, Majewski P, Moniuszko-Malinowska A. Prevalence of Tick-Borne Pathogens in Questing Ixodes ricinus and Dermacentor reticulatus Ticks Collected from Recreational Areas in Northeastern Poland with Analysis of Environmental Factors. Pathogens. 2022; 11(4):468. https://doi.org/10.3390/pathogens11040468
Chicago/Turabian StyleGrochowska, Anna, Justyna Dunaj-Małyszko, Sławomir Pancewicz, Piotr Czupryna, Robert Milewski, Piotr Majewski, and Anna Moniuszko-Malinowska. 2022. "Prevalence of Tick-Borne Pathogens in Questing Ixodes ricinus and Dermacentor reticulatus Ticks Collected from Recreational Areas in Northeastern Poland with Analysis of Environmental Factors" Pathogens 11, no. 4: 468. https://doi.org/10.3390/pathogens11040468