
Pathogenicity of Different Betanodavirus RGNNV/SJNNV Reassortant Strains in European Sea Bass

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Clinical Observation and Survivorship Curves
2.2. Molecular Viral Detection in Experimentally Infected Fish
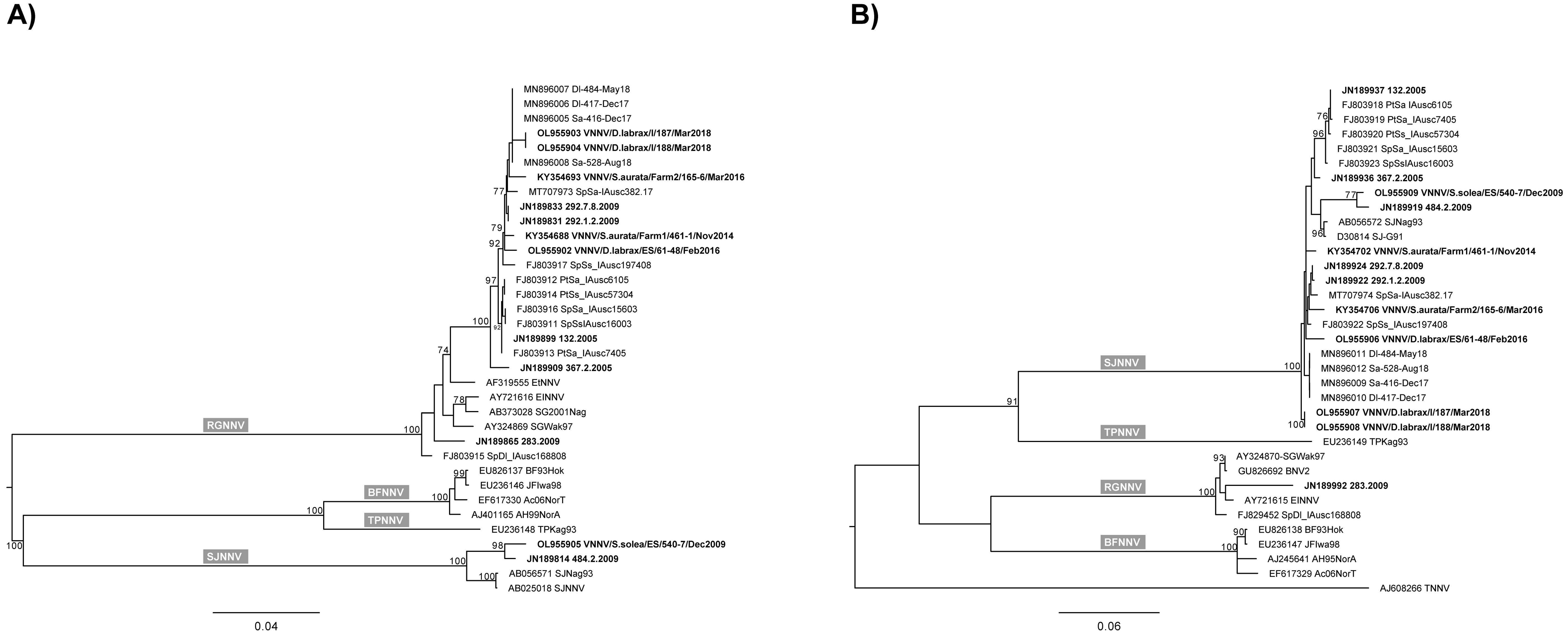
2.3. Genetic Characterization and Phylogenetic Analysis
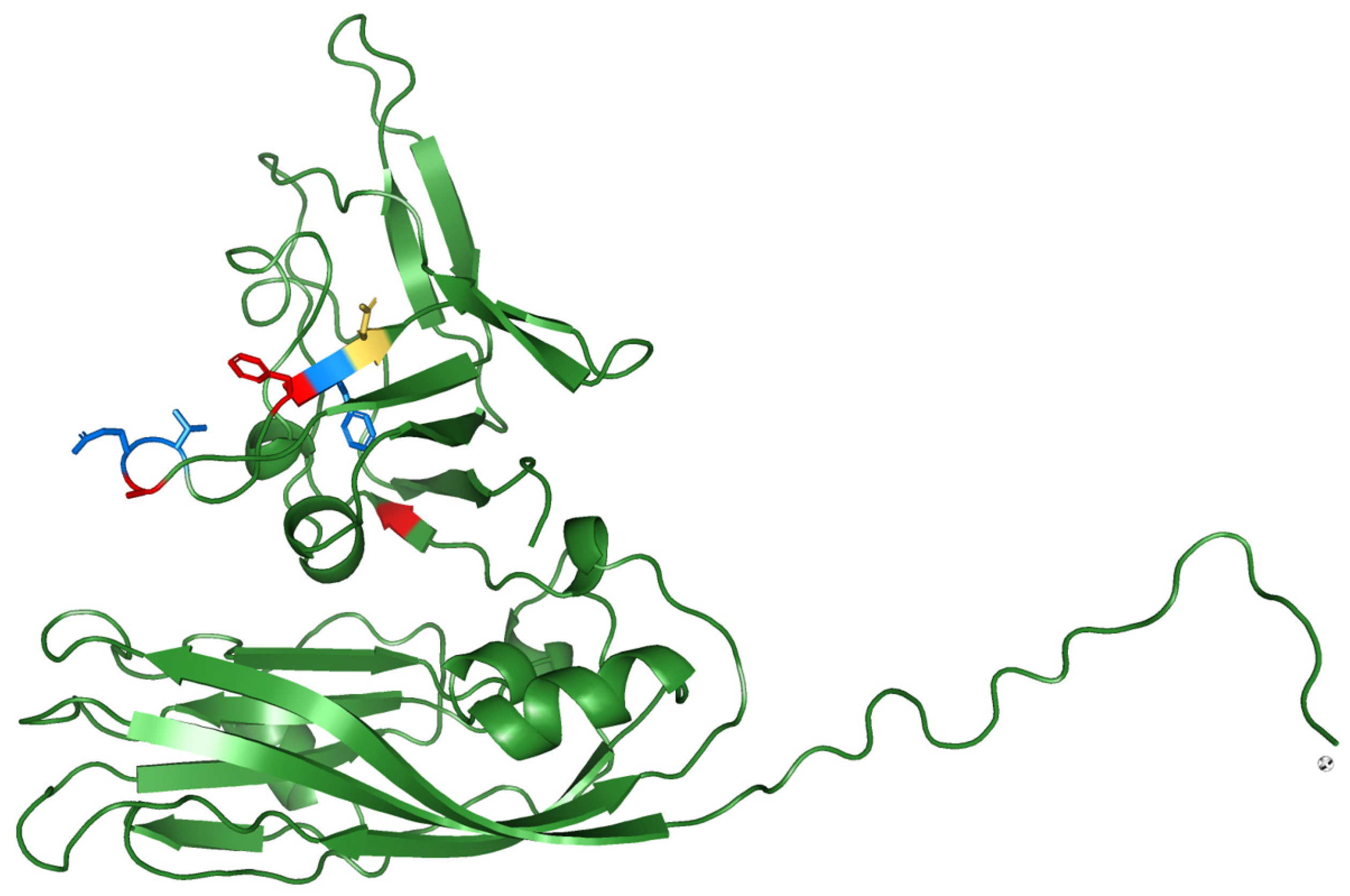
2.4. Protein-Structure Prediction
3. Discussion
4. Materials and Methods
4.1. Fish
4.2. Virus Isolation and Propagation
4.3. Experimental Challenges
4.3.1. Challenge by Intramuscular Injection
4.3.2. Challenge by Bath Immersion
4.4. Molecular Analysis for Viral RNA Detection in Experimentally Infected European Sea Bass
4.4.1. Qualitative Real-Time RT-PCR (rRT-PCR)
4.4.2. Quantification of RNA1 Genome Copy Numbers in Experimentally Infected Fish
4.5. Whole Genome Sequencing
4.6. Phylogenetic Analysis
4.7. Protein-Structure Prediction
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Toffolo, V.; Negrisolo, E.; Maltese, C.; Bovo, G.; Belvedere, P.; Colombo, L.; Dalla Valle, L. Phylogeny of betanodaviruses and molecular evolution of their RNA polymerase and coat proteins. Mol. Phylogenet. Evol. 2007, 43, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Crane, M.; Hyatt, A. Viruses of fish: An overview of significant pathogens. Viruses 2011, 3, 2025–2046. [Google Scholar] [CrossRef] [PubMed]
- Terlizzi, A.; Tedesco, P.; Patarnello, P. Spread of Pathogens from Marine Cage Aquaculture—A Potential Threat for Wild Fish Assemblages Under Protection Regimes. In Health and Enviroment in Aquaculture; Carvalho, E., Ed.; InTech: Rijeka, Croatia, 2012; Volume 16, pp. 403–414. ISBN 978-953-51-0497-1. [Google Scholar]
- Doan, Q.K.; Vandeputte, M.; Chatain, B.; Morin, T.; Allal, F. Viral encephalopathy and retinopathy in aquaculture: A review. J. Fish Dis. 2017, 40, 717–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muniesa, A.; Basurco, B.; Aguilera, C.; Furones, D.; Reverté, C.; Sanjuan-Vilaplana, A.; Jansen, M.D.; Brun, E.; Tavornpanich, S. Mapping the knowledge of the main diseases affecting sea bass and sea bream in Mediterranean. Transbound Emerg. Dis. 2020, 67, 1089–1100. [Google Scholar] [CrossRef] [PubMed]
- Vossen, M.T.M.; Westerhout, E.M.; Söderberg-Nauclér, C.; Wiertz, E.J.H.J. Viral immune evasion: A masterpiece of evolution. Immunogenetics 2002, 54, 527–542. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.I.; Nakai, T.; Muroga, K.; Arimoto, M.; Mushiake, K.; Furusawa, I. Properties of a new virus belonging to nodaviridae found in larval striped jack (Pseudocaranx dentex) with nervous necrosis. Virology 1992, 187, 368–371. [Google Scholar] [CrossRef]
- Iwamoto, T.; Mise, K.; Takeda, A.; Okinaka, Y.; Mori, K.I.; Arimoto, M.; Okuno, T.; Nakai, T. Characterization of striped jack nervous necrosis virus subgenomic RNA3 and biological activities of its encoded protein B2. J. Gen. Virol. 2005, 86, 2807–2816. [Google Scholar] [CrossRef]
- Fenner, B.J.; Thiagarajan, R.; Chua, H.K.; Kwang, J. Betanodavirus B2 is an RNA interference antagonist that facilitates intracellular viral RNA accumulation. J. Virol. 2006, 80, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Nishizawa, T.; Furuhashi, M.; Nagai, T.; Nakai, T.; Muroga, K. Genomic classification of fish nodaviruses by molecular phylogenetic analysis of the coat protein gene. Appl. Environ. Microbiol. 1997, 63, 1633–1636. [Google Scholar] [CrossRef] [Green Version]
- Olveira, J.G.; Souto, S.; Dopazo, C.P.; Thié, R.; Barja, J.L.; Bandín, I. Comparative analysis of both genomic segments of betanodaviruses isolated from epizootic outbreaks in farmed fish species provides evidence for genetic reassortment. J. Gen. Virol. 2009, 90, 2940–2951. [Google Scholar] [CrossRef]
- Panzarin, V.; Fusaro, A.; Monne, I.; Cappellozza, E.; Patarnello, P.; Bovo, G.; Capua, I.; Holmes, E.C.; Cattoli, G. Molecular epidemiology and evolutionary dynamics of betanodavirus in southern Europe. Infect. Genet. Evol. 2012, 12, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Bandín, I.; Souto, S. Betanodavirus and VER disease: A 30-year research review. Pathogens 2020, 9, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NaveenKumar, S.; Hassan, M.A.; Mahmoud, M.A.; Al-Ansari, A.; Al-Shwared, W.K. Betanodavirus infection in reared marine fishes along the Arabian Gulf. Aquac. Int. 2017, 25, 1543–1554. [Google Scholar] [CrossRef]
- Toffan, A.; Pascoli, F.; Pretto, T.; Panzarin, V.; Abbadi, M.; Buratin, A.; Quartesan, R.; Gijón, D.; Padrós, F. Viral nervous necrosis in gilthead sea bream (Sparus aurata) caused by reassortant betanodavirus RGNNV/SJNNV: An emerging threat for Mediterranean aquaculture. Sci. Rep. 2017, 7, 46755. [Google Scholar] [CrossRef] [PubMed]
- Volpe, E.; Gustinelli, A.; Caffara, M.; Errani, F.; Quaglio, F.; Fioravanti, M.L.; Ciulli, S. Viral nervous necrosis outbreaks caused by the RGNNV/SJNNV reassortant betanodavirus in gilthead sea bream (Sparus aurata) and European sea bass (Dicentrarchus labrax). Aquaculture 2020, 523, 735155. [Google Scholar] [CrossRef]
- Savoca, S.; Abbadi, M.; Toffan, A.; Salogni, C.; Iaria, C.; Capparucci, F.; Quartesan, R.; Alborali, G.L.; Guarnera, S.; Cangemi, G.; et al. Betanodavirus infection associated with larval enteropathy as a cause of mortality in cultured gilthead sea bream (Sparus aurata, Linnaeus, 1758). Aquaculture 2021, 541, 736844. [Google Scholar] [CrossRef]
- Federation of European Aquaculture Producers—FEAP. Available online: https://feap.info/index.php/data/ (accessed on 11 February 2022).
- Shetty, M.; Maiti, B.; Shivakumar Santhosh, K.; Venugopal, M.N.; Karunasagar, I. Betanodavirus of marine and freshwater fish: Distribution, genomic organization, diagnosis and control measures. Indian J. Virol. 2012, 23, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Munday, B.L.; Kwang, J.; Moody, N. Betanodavirus infections of teleost fish: A review. J. Fish Dis. 2002, 25, 127–142. [Google Scholar] [CrossRef]
- Chérif, N.; Thiéry, R.; Castric, J.; Biacchesi, S.; Brémont, M.; Thabti, F.; Limem, L.; Hammami, S. Viral encephalopathy and retinopathy of Dicentrarchus labrax and Sparus aurata farmed in Tunisia. Vet. Res. Commun. 2009, 33, 345–353. [Google Scholar] [CrossRef]
- Vendramin, N.; Toffan, A.; Mancin, M.; Cappellozza, E.; Panzarin, V.; Bovo, G.; Cattoli, G.; Capua, I.; Terregino, C. Comparative pathogenicity study of ten different betanodavirus strains in experimentally infected European sea bass, Dicentrarchus labrax (L.). J. Fish Dis. 2014, 37, 371–383. [Google Scholar] [CrossRef]
- Souto, S.; Lopez-Jimena, B.; Alonso, M.C.; García-Rosado, E.; Bandín, I. Experimental susceptibility of European sea bass and Senegalese sole to different betanodavirus isolates. Vet. Microbiol. 2015, 177, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Toffan, A.; Panzarin, V.; Toson, M.; Cecchettin, K.; Pascoli, F. Water temperature affects pathogenicity of different betanodavirus genotypes in experimentally challenged Dicentrarchus labrax. Dis. Aquat. Organ. 2016, 119, 231–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souto, S.; Mérour, E.; Biacchesi, S.; Brémont, M.; Olveira, J.G.; Bandín, I. In vitro and in vivo characterization of molecular determinants of virulence in reassortant betanodavirus. J. Gen. Virol. 2015, 96, 1287–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, P.; Souto, S.; Leiva-Rebollo, R.; Borrego, J.J.; Bandín, I.; Alonso, M.C. Capsid amino acids at positions 247 and 270 are involved in the virulence of betanodaviruses to European sea bass. Sci. Rep. 2019, 9, 14068. [Google Scholar] [CrossRef]
- Castric, J.; Thiéry, R.; Jeffroy, J.; de Kinkelin, P.; Raymond, J.C. Sea bream Sparus aurata, an asymptomatic contagious fish host for nodavirus. Dis. Aquat. Organ. 2001, 47, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korsnes, K.; Karlsbakk, E.; Nylund, A.; Nerland, A.H. Horizontal transmission of nervous necrosis virus between turbot Scophthalmus maximus and Atlantic cod Gadus morhua using cohabitation challenge. Dis. Aquat. Organ. 2012, 99, 13–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, Y.; Okinaka, Y.; Mori, K.-I.; Sugaya, T.; Nishioka, T.; Oka, M.; Nakai, T. Variable region of betanodavirus RNA2 is sufficient to determine host specificity. Dis. Aquat. Organ. 2008, 79, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, T.; Okinaka, Y.; Mise, K.; Mori, K.-I.; Arimoto, M.; Okuno, T.; Nakai, T. Identification of host-specificity determinants in betanodaviruses by using reassortants between striped jack nervous necrosis virus and sevenband grouper nervous necrosis virus. J. Virol. 2004, 78, 1256–1262. [Google Scholar] [CrossRef] [Green Version]
- Chao, J.A.; June, H.L.; Chapados, B.R.; Debler, E.W.; Schneemann, A.; Williamson, J.R. Dual modes of RNA-silencing suppression by Flock House virus protein B2. Nat. Struct. Mol. Biol. 2005, 12, 952–957. [Google Scholar] [CrossRef]
- Lingel, A.; Simon, B.; Izaurralde, E.; Sattler, M. The structure of the flock house virus B2 protein, a viral suppressor of RNA interference, shows a novel mode of double-stranded RNA recognition. EMBO Rep. 2005, 6, 1149–1155. [Google Scholar] [CrossRef]
- Souto, S.; Olveira, J.G.; Dopazo, C.P.; Borrego, J.J.; Bandín, I. Modification of betanodavirus virulence by substitutions in the 3′ terminal region of RNA2. J. Gen. Virol. 2018, 99, 1210–1220. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, Y.; Wang, M.; Cheng, A.; Yang, Q.; Wu, Y.; Jia, R.; Liu, M.; Zhu, D.; Chen, S.; et al. Structures and Functions of the 3′ Untranslated Regions of Positive-Sense Single-Stranded RNA Viruses Infecting Humans and Animals. Front. Cell. Infect. Microbiol. 2020, 10, 453. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, T.; Nakai, T.; Mori, K.; Arimoto, M.; Furusawa, I. Cloning of the fish cell line SSN-1 for piscine nodaviruses. Dis. Aquat. Organ. 2000, 43, 81–89. [Google Scholar] [CrossRef] [PubMed]
- OIE—Viral encephalopathy and retinopathy. In Manual of Diagnostic Tests for Aquatic Animals; World Organisation for Animal Health (OIE): Paris, France, 2019; Chapter 2.3.12; pp. 1–20.
- Pascoli, F.; Guazzo, A.; Buratin, A.; Toson, M.; Buonocore, F.; Scapigliati, G.; Toffan, A. Lack of in vivo cross-protection of two different betanodavirus species RGNNV and SJNNV in European sea bass Dicentrachus labrax. Fish Shellfish. Immunol. 2019, 85, 85–89. [Google Scholar] [CrossRef]
- Frerichs, G.N.; Rodger, H.D.; Peric, Z. Cell culture isolation of piscine neuropathy nodavirus from juvenile sea bass, Dicentrarchus labrax. J. Gen. Virol. 1996, 77, 2067–2071. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Baud, M.; Cabon, J.; Salomoni, A.; Toffan, A.; Panzarin, V.; Bigarré, L. First generic one step real-time Taqman RT-PCR targeting the RNA1 of betanodaviruses. J. Virol. Methods 2015, 211, 1–7. [Google Scholar] [CrossRef]
- Toffan, A.; Biasini, L.; Pretto, T.; Abbadi, M.; Buratin, A.; Franch, R.; Dalla Rovere, G.; Panzarin, V.M.; Marsella, A.; Bargelloni, L.; et al. Age dependency of RGNNV/SJNNV viral encephalo-retinopathy in Gilthead Sea Bream (Sparus aurata). Aquaculture 2021, 539, 736605. [Google Scholar] [CrossRef]
- Panzarin, V.; Cappellozza, E.; Mancin, M.; Milani, A.; Toffan, A.; Terregino, C.; Cattoli, G. In vitro study of the replication capacity of the RGNNV and the SJNNV betanodavirus genotypes and their natural reassortants in response to temperature. Vet. Res. 2014, 45, 56. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0. molecular biology and evolution. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Belle, G.; Fisher, L.; Heagerty, P.J.; Lumley, T. Biostatistics: A Methodology for the Health Sciences, 2nd ed.; John Wiley & Sons: New York, NY, USA, 2004; ISBN 978-0-471-03185-7. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Abbreviation | Year | Host | Origin | Genotype | RNA1 | RNA2 | Reference |
|---|---|---|---|---|---|---|---|---|
| GenBank Accession No. | ||||||||
| 283.2009 | 283 | 2009 | D. labrax | Italy | RG | JN189865.2 | JN189992.2 | [12] |
| 132.2005 | 132 | 2005 | D. labrax | Italy | RG/SJ | JN189899.2 | JN189937.2 | [12] |
| 292.7.8.2009 | 292-7.8 | 2009 | D. labrax | Greece | RG/SJ | JN189833.2 | JN189924.2 | [12] |
| 292.1.2.2009 | 292-1.2 | 2009 | S. aurata | Greece | RG/SJ | JN189831.2 | JN189922.2 | [12] |
| VNNV/D.labrax/I/187/Mar2018 | 187 | 2018 | D. labrax | Italy | RG/SJ | OL955903 | OL955907 | This study |
| 367.2.2005 | 367-2 | 2005 | D. labrax | Italy | RG/SJ | JN189909.2 | JN189936.2 | [12] |
| VNNV/D.labrax/ES/61-48/Feb2016 | 61-48 | 2016 | D. labrax | Spain | RG/SJ | OL955902 | OL955906 | This study |
| VNNV/S.aurata/I/188/Mar2018 | 188 | 2018 | S. aurata | Italy | RG/SJ | OL955904 | OL955908 | This study |
| VNNV/S.aurata/Farm1/461-1/Nov2014 | 461-1 | 2014 | S. aurata | n.d. | RG/SJ | KY354688.2 | KY354702.2 | [15] |
| VNNV/S.aurata/Farm2/165-6/Mar2016 | 165-6 | 2016 | S. aurata | n.d. | RG/SJ | KY354693.2 | KY354706.2 | [15] |
| VNNV/S.solea/ES/540-7/Dec2009 | 540-7 | 2009 | S. solea | Spain | SJ | OL955905 | OL955909 | This study |
| 484.2.2009 | 484-2 | 2009 | S. senegalensis | Spain | SJ | JN189814.2 | JN189919.2 | [12] |
| Route of Viral Infection | Challenge Group | TCID50/mL 1 | % Mortality (95% CI) | LCN ± SD 2 |
|---|---|---|---|---|
| Intramuscular injection | RG283 | 105.80 | 34.55 (22.24–48.58) | 5.26 ± 0.16 |
| RG/SJ132 | 106.80 | 26.00 (14.63–40.35) | 4.81 ± 0.32 | |
| RG/SJ292-7.8 | 105.80 | 15.69 (7.02–28.59) | n.e. | |
| RG/SJ292-1.2 | 105.80 | 12.73 (5.27–24.48) | 4.84 ± 0.48 | |
| RG/SJ187 | 106.05 | 10.91 (4.11–22.25) | 4.90 ± 0.27 | |
| RG/SJ367-2 | 105.80 | 7.14 (1.98–17.29) | 5.01 ± 0.47 | |
| RG/SJ61-48 | 107.05 | 6.38 (1.34–17.54) | 4.82 ± 0.21 | |
| RG/SJ188 | 106.80 | 5.66 (1.18–15.66) | n.e. | |
| RG/SJ461-1 | 107.55 | 4.08 (0.50–13.98) | 4.34 ± 0.31 | |
| RG/SJ165-6 | 106.55 | 2.04 (0.05–10.85) | 4.74 ± 0.27 | |
| SJ540-7 | 106.80 | 1.82 (0.05–9.72) | n.a. | |
| SJ484-2 | 107.30 | 0.00 (0.00–6.49) | n.a. | |
| Bath immersion | RG283 | 105.05 | 18.52 (9.25–31.43) | 4.67 ± 0.39 |
| RG/SJ132 | 105.05 | 7.41 (2.06–17.89) | 4.79 ± 0.37 | |
| RG/SJ292-7.8 | 105.05 | 3.70 (0.45–12.75) | 4.27 ± 0.58 | |
| RG/SJ461-1 | 105.05 | 7.41 (2.06–17.89) | 4.43 ± 1.08 # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biasini, L.; Berto, P.; Abbadi, M.; Buratin, A.; Toson, M.; Marsella, A.; Toffan, A.; Pascoli, F. Pathogenicity of Different Betanodavirus RGNNV/SJNNV Reassortant Strains in European Sea Bass. Pathogens 2022, 11, 458. https://doi.org/10.3390/pathogens11040458
Biasini L, Berto P, Abbadi M, Buratin A, Toson M, Marsella A, Toffan A, Pascoli F. Pathogenicity of Different Betanodavirus RGNNV/SJNNV Reassortant Strains in European Sea Bass. Pathogens. 2022; 11(4):458. https://doi.org/10.3390/pathogens11040458
Chicago/Turabian StyleBiasini, Lorena, Paola Berto, Miriam Abbadi, Alessandra Buratin, Marica Toson, Andrea Marsella, Anna Toffan, and Francesco Pascoli. 2022. "Pathogenicity of Different Betanodavirus RGNNV/SJNNV Reassortant Strains in European Sea Bass" Pathogens 11, no. 4: 458. https://doi.org/10.3390/pathogens11040458