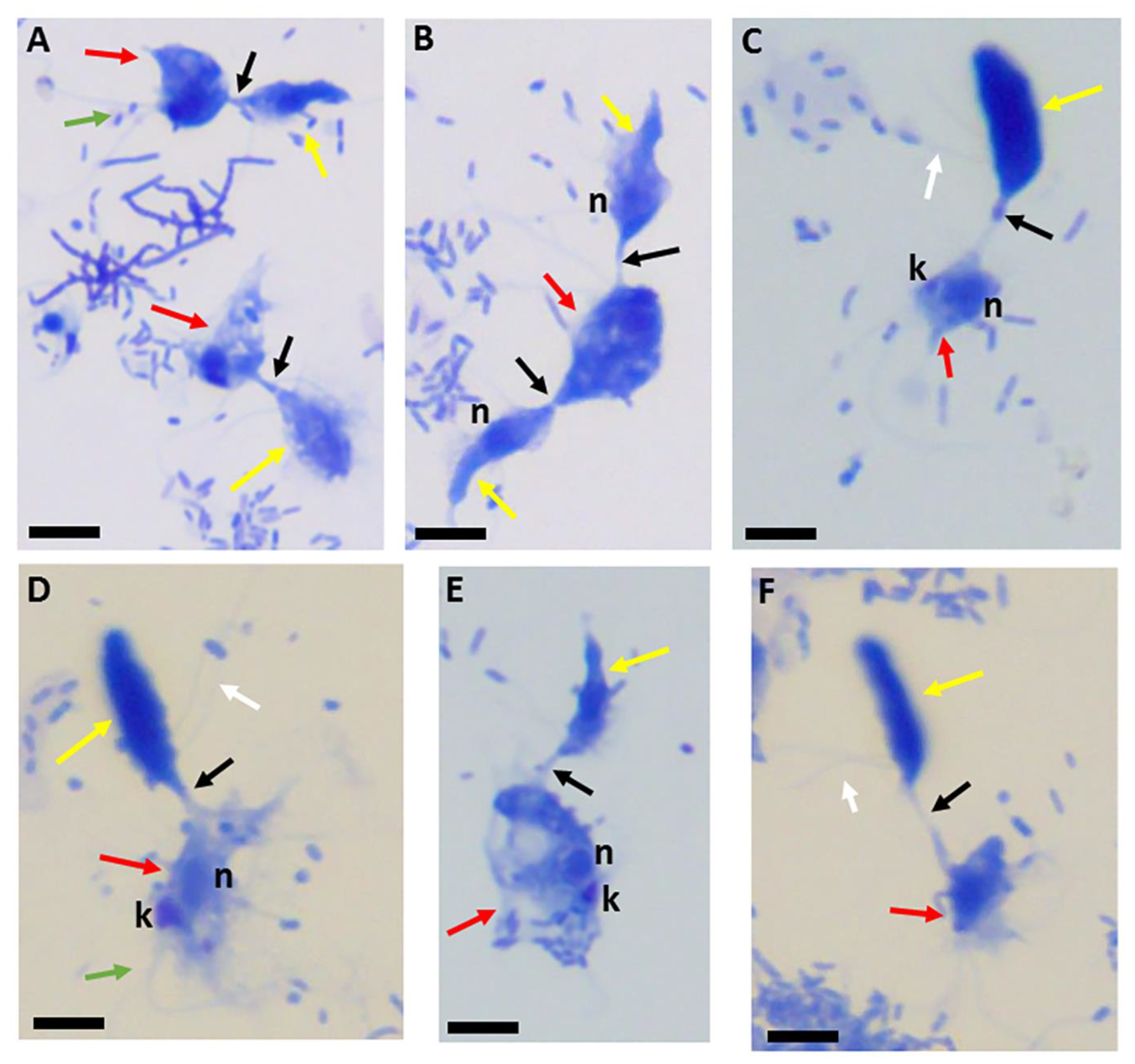
Figure 1.
Giemsa-stained formalin-fixed Colpodella sp. (ATCC 50594) (yellow arrow) and Parabodo caudatus (red arrow) in myzocytosis showing different lengths of the tubular tether formed between both protists (black arrow). The flagella of Colpodella sp. (ATCC 50594) are indicated by the white arrows and flagella for P. caudatus by the green arrow. Attachment can occur with a single trophozoite of Colpodella sp. (ATCC 50594) attached to P. caudatus (A,C–F) or P. caudatus can be attacked by two Colpodella sp. (ATCC 50594) trophozoites at the same time as seen in panel (B). Scale bars, 10 µm.
Figure 1.
Giemsa-stained formalin-fixed Colpodella sp. (ATCC 50594) (yellow arrow) and Parabodo caudatus (red arrow) in myzocytosis showing different lengths of the tubular tether formed between both protists (black arrow). The flagella of Colpodella sp. (ATCC 50594) are indicated by the white arrows and flagella for P. caudatus by the green arrow. Attachment can occur with a single trophozoite of Colpodella sp. (ATCC 50594) attached to P. caudatus (A,C–F) or P. caudatus can be attacked by two Colpodella sp. (ATCC 50594) trophozoites at the same time as seen in panel (B). Scale bars, 10 µm.
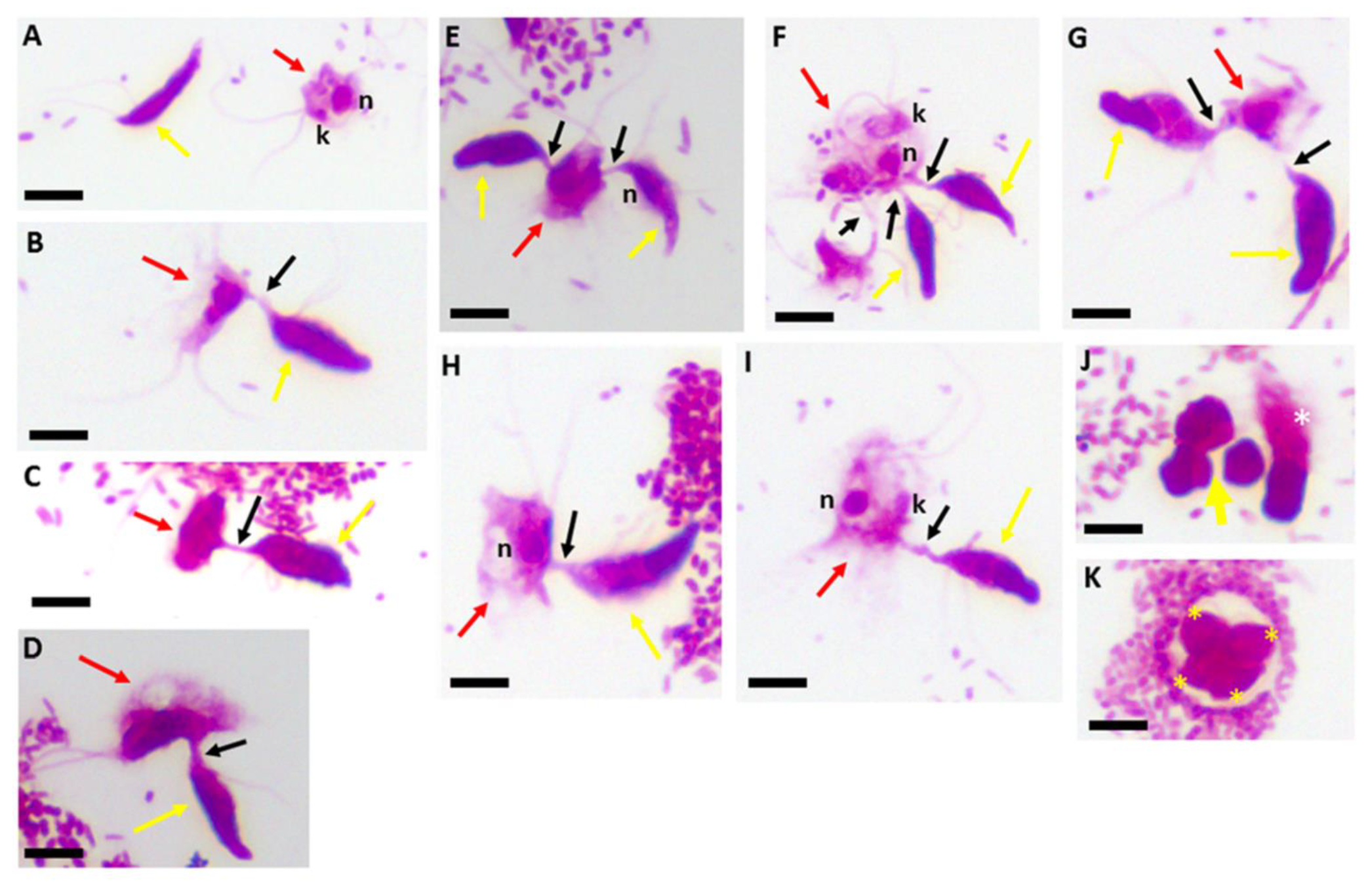
Figure 2.
Kinyoun’s carbol fuchsin-stained formalin-fixed Colpodella sp. (ATCC 50594) (yellow arrow) and Parabodo caudatus (red arrow) shown individually (A), in myzocytosis showing different lengths of the tubular tether formed between the two protists and the flagella of both protists. Single attachments are shown in panels (B–D,H,I). Parabodo caudatus attacked by two trophozoites of Colpodella sp. (ATCC 50594) show that predators can attach to prey in close proximity or on opposite sides of the prey (E–G). Demilune cysts of Colpodella sp. (ATCC 50594) are shown in panel (J) (yellow arrow) and a pre-cyst showing the frayed anterior end of the trophozoite (white asterisk) is shown. A mature cyst of Colpodella sp. (ATCC 50594) containing four trophozoites (yellow asterisks) and a clear zone surrounding the cyst, free of bacteria is shown in panel (K). Scale bars, 10 µm.
Figure 2.
Kinyoun’s carbol fuchsin-stained formalin-fixed Colpodella sp. (ATCC 50594) (yellow arrow) and Parabodo caudatus (red arrow) shown individually (A), in myzocytosis showing different lengths of the tubular tether formed between the two protists and the flagella of both protists. Single attachments are shown in panels (B–D,H,I). Parabodo caudatus attacked by two trophozoites of Colpodella sp. (ATCC 50594) show that predators can attach to prey in close proximity or on opposite sides of the prey (E–G). Demilune cysts of Colpodella sp. (ATCC 50594) are shown in panel (J) (yellow arrow) and a pre-cyst showing the frayed anterior end of the trophozoite (white asterisk) is shown. A mature cyst of Colpodella sp. (ATCC 50594) containing four trophozoites (yellow asterisks) and a clear zone surrounding the cyst, free of bacteria is shown in panel (K). Scale bars, 10 µm.
Figure 3.
Sam-Yellowe’s trichrome-stained formalin-fixed Colpodella sp. (ATCC 50594) (yellow arrow) and Parabodo caudatus (red arrow) in myzocytosis showing different lengths of the tubular tether and differentiation of cyst stages. Single predator attacks are shown in panels (A,C–E). A single prey attacked by two predators is shown in panel (B). The nuclei (n) of both protists are shown in panels (A,C). The kinetoplast (k) of Parabodo caudatus is shown in panel (C). An enlarged darkly stained food vacuole in Colpodella sp. (ATCC 50594) is seen in panel (D). Demilune and single nucleus (yellow asterisk) cysts are shown in panel (F). Parabodo caudatus cysts (red arrow) can be distinguished from Colpodella sp. (ATCC 50594) cysts. The single nucleus cyst of Colpodella sp. (ATCC 50594) indicated by the yellow asterisk can be distinguished from the immature cysts of Colpodella sp. (ATCC 50594) showing partly light and dark stained sections in the cyst (yellow arrow). Scale bars, 10 µm.
Figure 3.
Sam-Yellowe’s trichrome-stained formalin-fixed Colpodella sp. (ATCC 50594) (yellow arrow) and Parabodo caudatus (red arrow) in myzocytosis showing different lengths of the tubular tether and differentiation of cyst stages. Single predator attacks are shown in panels (A,C–E). A single prey attacked by two predators is shown in panel (B). The nuclei (n) of both protists are shown in panels (A,C). The kinetoplast (k) of Parabodo caudatus is shown in panel (C). An enlarged darkly stained food vacuole in Colpodella sp. (ATCC 50594) is seen in panel (D). Demilune and single nucleus (yellow asterisk) cysts are shown in panel (F). Parabodo caudatus cysts (red arrow) can be distinguished from Colpodella sp. (ATCC 50594) cysts. The single nucleus cyst of Colpodella sp. (ATCC 50594) indicated by the yellow asterisk can be distinguished from the immature cysts of Colpodella sp. (ATCC 50594) showing partly light and dark stained sections in the cyst (yellow arrow). Scale bars, 10 µm.
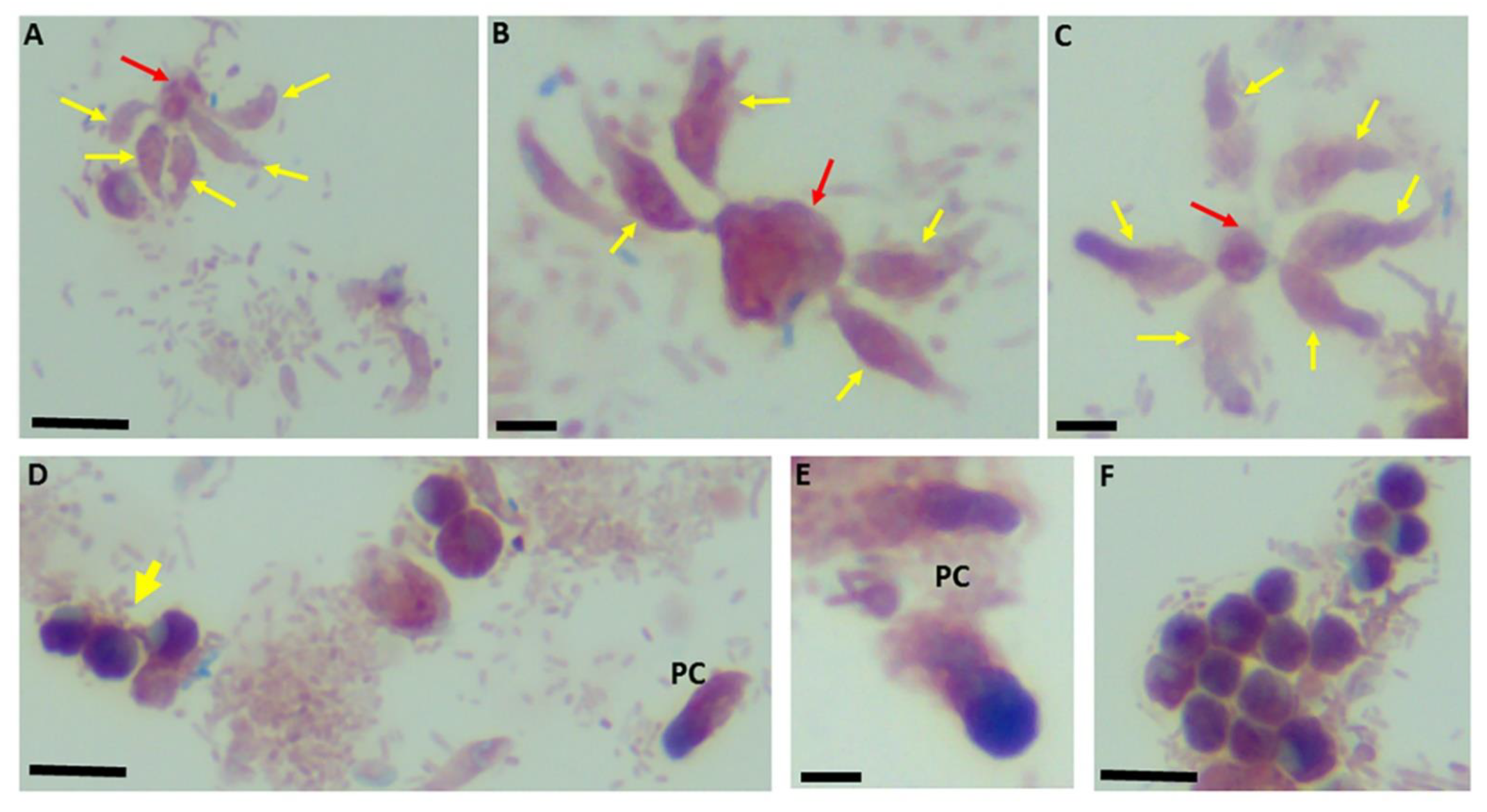
Figure 4.
Sam-Yellowe’s trichrome staining of formalin-fixed Colpodella sp. (ATCC 50594) trophozoites (yellow arrows) attached to P. caudatus (red arrow). Multiple predators are shown attached to a single P. caudatus prey (A–C). Demilune cysts of Colpodella sp. (ATCC 50594) are shown in panels (D,F). Pre-cysts (PC) of Colpodella sp. (ATCC 50594) are shown in panels (D,E). Scale bars, 10 µm in panels (B,C,E). Scale bars, 15 µm in panels (A,D,F).
Figure 4.
Sam-Yellowe’s trichrome staining of formalin-fixed Colpodella sp. (ATCC 50594) trophozoites (yellow arrows) attached to P. caudatus (red arrow). Multiple predators are shown attached to a single P. caudatus prey (A–C). Demilune cysts of Colpodella sp. (ATCC 50594) are shown in panels (D,F). Pre-cysts (PC) of Colpodella sp. (ATCC 50594) are shown in panels (D,E). Scale bars, 10 µm in panels (B,C,E). Scale bars, 15 µm in panels (A,D,F).
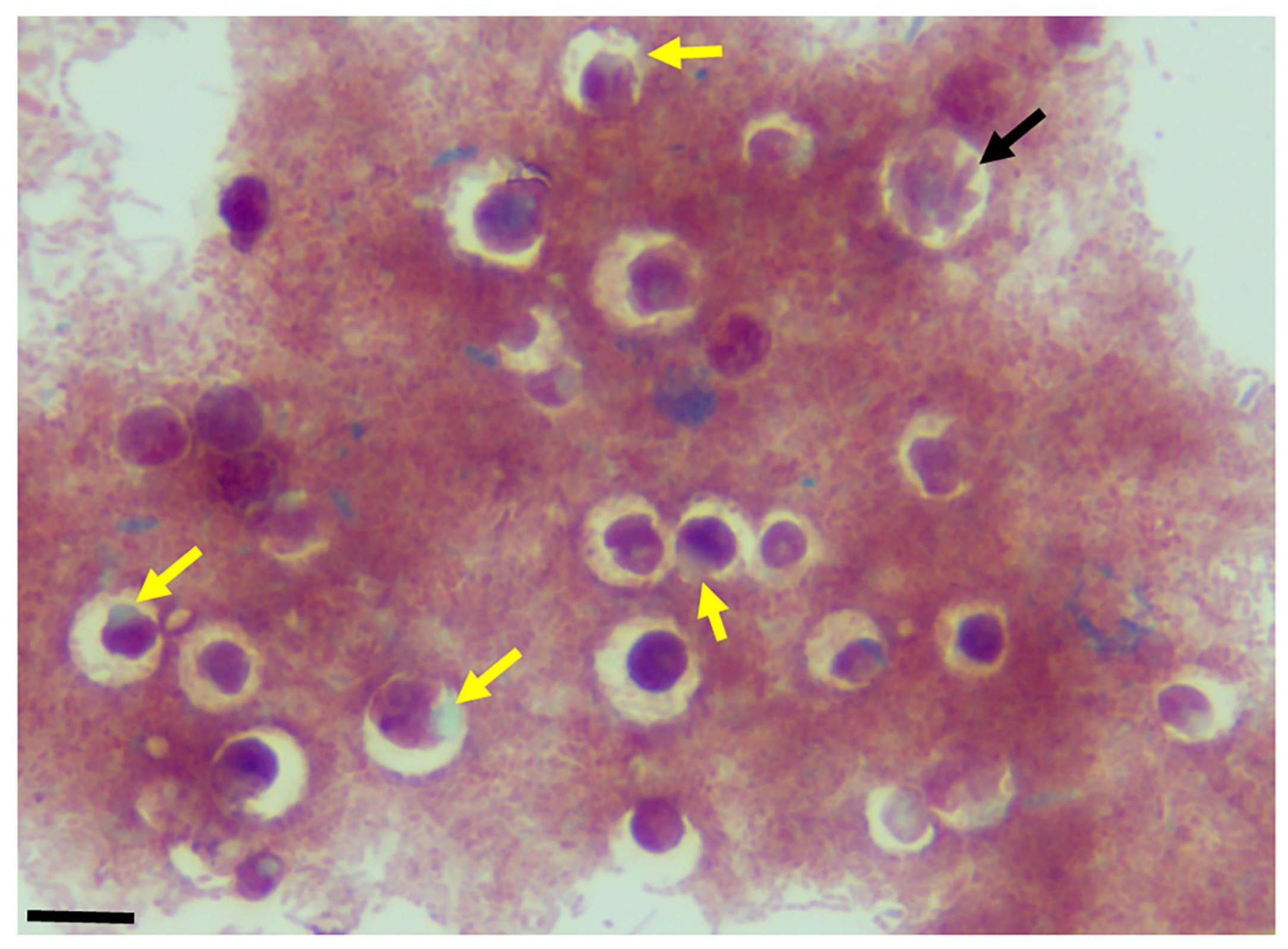
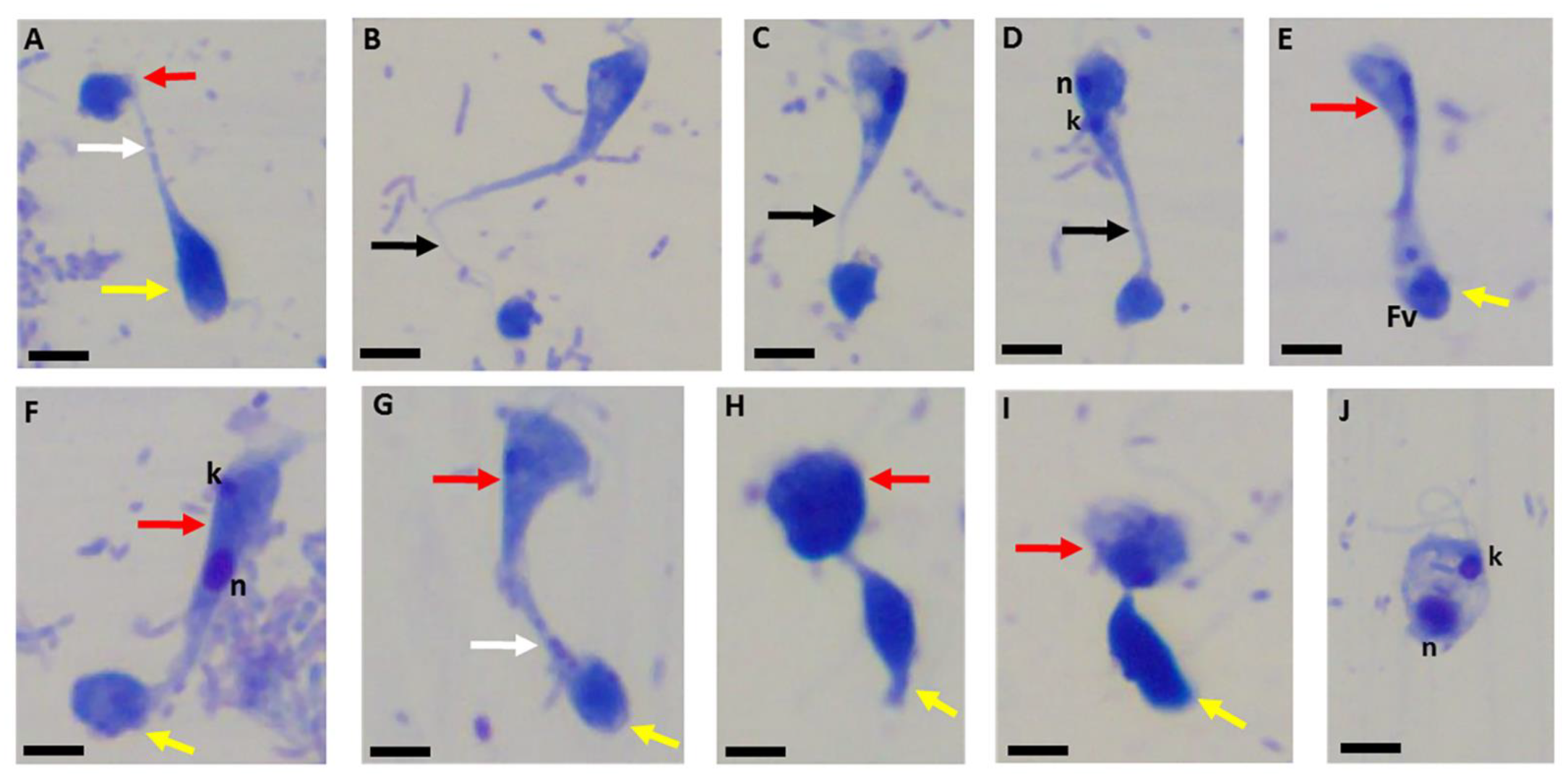
Figure 5.
Sam-Yellowe’s trichrome-stained formalin-fixed cysts of Colpodella sp. (ATCC 50594) (yellow arrow) showing clear bacteria-free zones surrounding cysts. A mature cyst of Colpodella sp. (ATCC 50594) containing multiple juvenile trophozoites is shown (black arrow). Scale bars, 15 µm.
Figure 5.
Sam-Yellowe’s trichrome-stained formalin-fixed cysts of Colpodella sp. (ATCC 50594) (yellow arrow) showing clear bacteria-free zones surrounding cysts. A mature cyst of Colpodella sp. (ATCC 50594) containing multiple juvenile trophozoites is shown (black arrow). Scale bars, 15 µm.
Figure 6.
Transmission electron microscopy showing attachment site of Colpodella sp. (ATCC 50594) trophozoite (yellow arrow) to P. caudatus (red arrow). The attachment site is posterior to the apical tip of the rostrum indicated by the orange arrow (A). (B) shows the attachment site in an enlargement of panel (A) showing a two-point attachment (black arrows in boxed area). The white arrows identify the membrane of Colpodella sp. (ATCC 50594 at the attachment site. Bacteria (B) in the cytoplasm of P. caudatus are shown. Mitochondria (m) were also detected. Scale bars, (A) 1 µm and (B) 0.5 µm.
Figure 6.
Transmission electron microscopy showing attachment site of Colpodella sp. (ATCC 50594) trophozoite (yellow arrow) to P. caudatus (red arrow). The attachment site is posterior to the apical tip of the rostrum indicated by the orange arrow (A). (B) shows the attachment site in an enlargement of panel (A) showing a two-point attachment (black arrows in boxed area). The white arrows identify the membrane of Colpodella sp. (ATCC 50594 at the attachment site. Bacteria (B) in the cytoplasm of P. caudatus are shown. Mitochondria (m) were also detected. Scale bars, (A) 1 µm and (B) 0.5 µm.
Figure 7.
Transmission electron microscopy showing attachment of Colpodella sp. (ATCC 50594) trophozoites (yellow arrow) to P. cuadatus (red arrow). Panel (A) is enlarged in panels (B–D) to show the detail at the attachment site. The plasma membrane and cytoplasm of the prey are aspirated into the cytoplasm of the predator. White arrows show the prey’s plasma membrane. The attachment site is posterior to the apical tip (orange arrow) of the rostrum. Microtubules organized at the attachment zone (black arrows) are shown. Panels (C,D) show details of the attachment site and the tubular tether consisting of membranes of Colpodella sp. (ATCC 50594) and P. caudatus. The membranes of Colpodella sp. (ATCC 50594) surround the membrane and cytoplasm of P. caudatus. Thickening of the pellicle is shown with the three black arrows on either side of the tether. aF, anterior flagellum; B, bacteria; m, mitochondria. Scale bars, (A) 2 µm, (B) 0.5 µm, (C) 500 nm and (D) 200 nm.
Figure 7.
Transmission electron microscopy showing attachment of Colpodella sp. (ATCC 50594) trophozoites (yellow arrow) to P. cuadatus (red arrow). Panel (A) is enlarged in panels (B–D) to show the detail at the attachment site. The plasma membrane and cytoplasm of the prey are aspirated into the cytoplasm of the predator. White arrows show the prey’s plasma membrane. The attachment site is posterior to the apical tip (orange arrow) of the rostrum. Microtubules organized at the attachment zone (black arrows) are shown. Panels (C,D) show details of the attachment site and the tubular tether consisting of membranes of Colpodella sp. (ATCC 50594) and P. caudatus. The membranes of Colpodella sp. (ATCC 50594) surround the membrane and cytoplasm of P. caudatus. Thickening of the pellicle is shown with the three black arrows on either side of the tether. aF, anterior flagellum; B, bacteria; m, mitochondria. Scale bars, (A) 2 µm, (B) 0.5 µm, (C) 500 nm and (D) 200 nm.
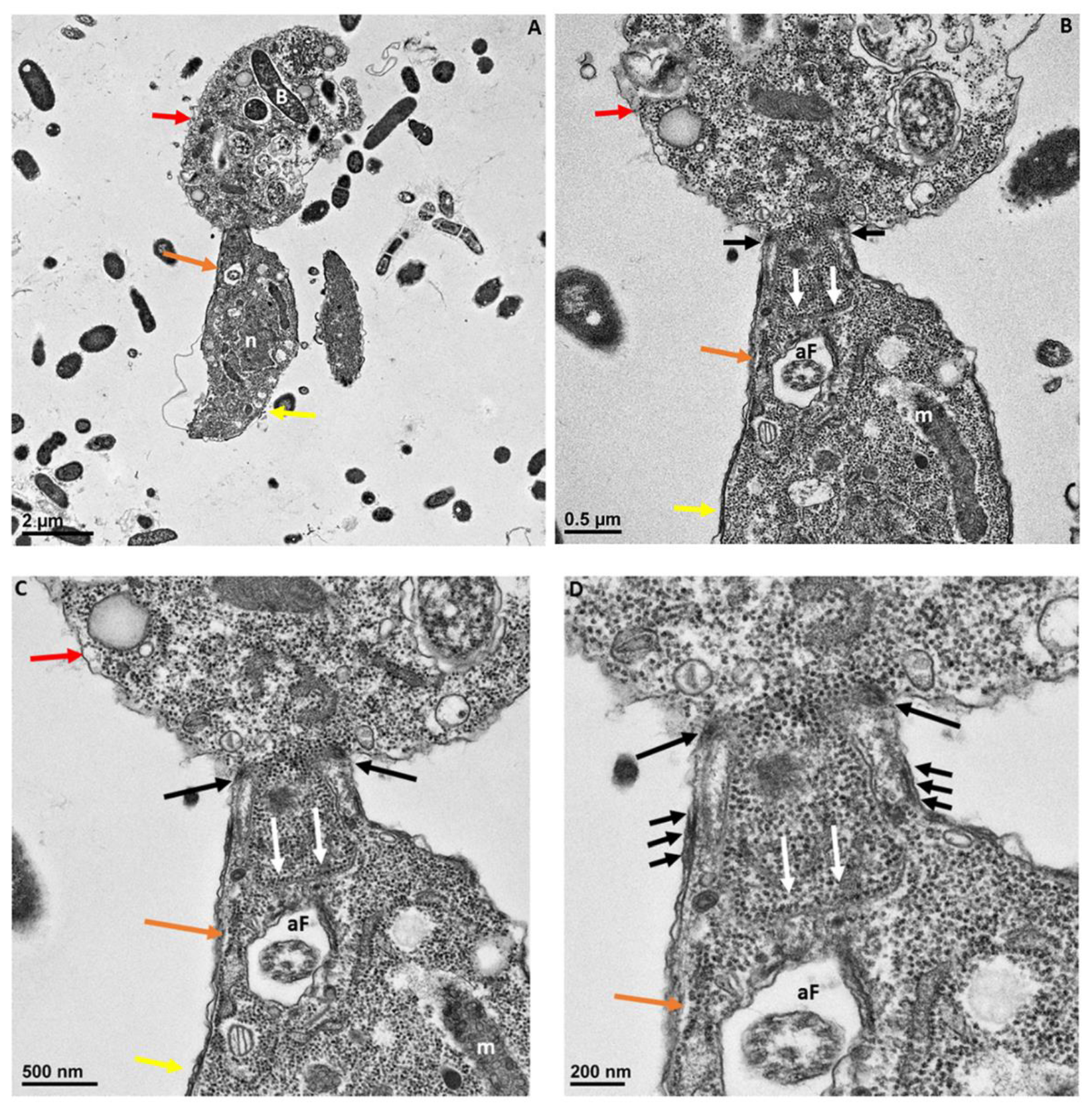
Figure 8.
Transmission electron microscopy showing attachment of Colpodella sp. (ATCC 50594) (yellow arrow) to P. caudatus (red arrow). Panel (A) is enlarged to show details of attachment and tubular tether in panels (B–D). Orange arrow shows apical tip of rostrum. The attachment site is indicated by black arrows and shows the membrane of Colpodella sp. (ATCC 50594) trophozoite surrounding the plasma membrane and cytoplasm of aspirated P. caudatus (panel (B)). The progression of aspirated contents, organelles, and plasma membrane of P. caudatus is indicated by the white arrows. Thickened pellicle of Colpodella sp. (ATCC 50594) and microtubular organization is shown by the triple arrows. An area posterior to the apical tip of the rostrum forms an “aperture” or duct that surrounds and pulls in the plasma membrane and cytoplasmic contents of the prey (black arrows). aF, anterior flagellum; B, bacteria; m, mitochondria; R, rhoptries; V, vacuole. Scale bars, (A) 1 µm, (B) 0.5 µm, (C) 200 nm, and (D) 100 nm.
Figure 8.
Transmission electron microscopy showing attachment of Colpodella sp. (ATCC 50594) (yellow arrow) to P. caudatus (red arrow). Panel (A) is enlarged to show details of attachment and tubular tether in panels (B–D). Orange arrow shows apical tip of rostrum. The attachment site is indicated by black arrows and shows the membrane of Colpodella sp. (ATCC 50594) trophozoite surrounding the plasma membrane and cytoplasm of aspirated P. caudatus (panel (B)). The progression of aspirated contents, organelles, and plasma membrane of P. caudatus is indicated by the white arrows. Thickened pellicle of Colpodella sp. (ATCC 50594) and microtubular organization is shown by the triple arrows. An area posterior to the apical tip of the rostrum forms an “aperture” or duct that surrounds and pulls in the plasma membrane and cytoplasmic contents of the prey (black arrows). aF, anterior flagellum; B, bacteria; m, mitochondria; R, rhoptries; V, vacuole. Scale bars, (A) 1 µm, (B) 0.5 µm, (C) 200 nm, and (D) 100 nm.
![Pathogens 11 00455 g008]()
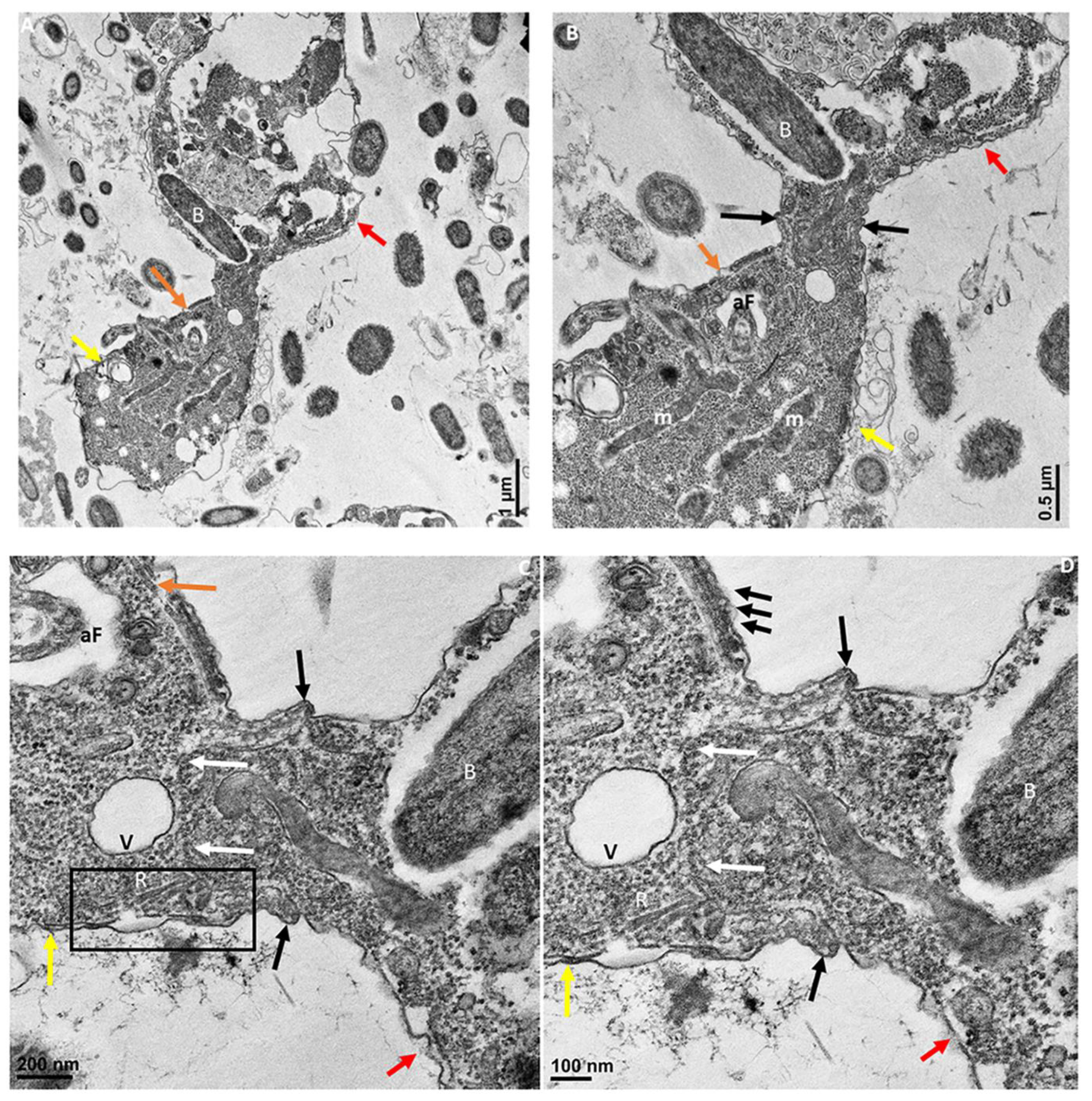
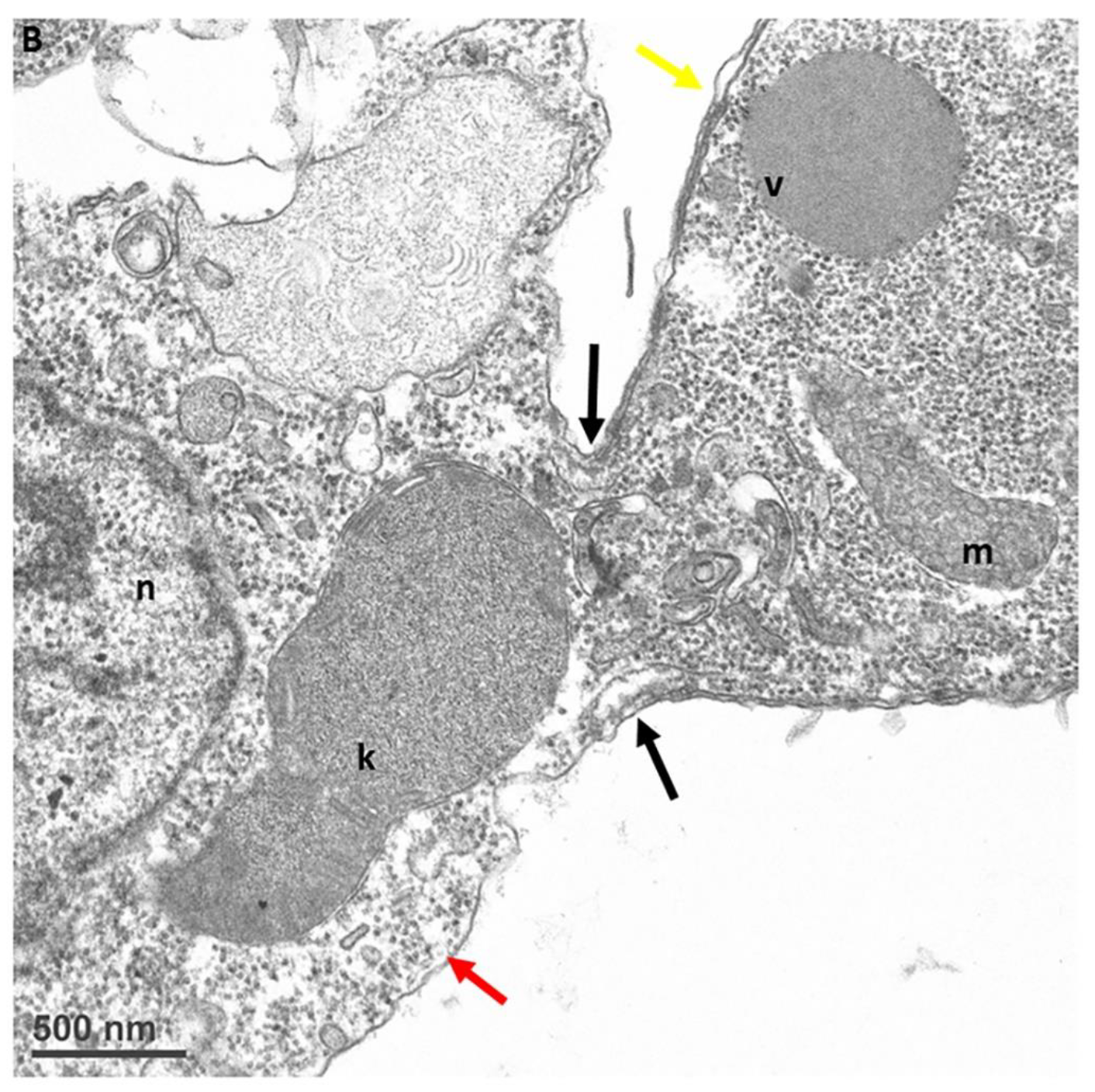
Figure 9.
Transmission electron microscopy showing attachment of Colpodella sp. (ATCC 50594) (yellow arrow) to P. caudatus (red arrow). Panel (A) shows attachment of a Colpodella sp. (ATCC 50594) trophozoite to P. caudatus. Contents from the prey flow into the cytoplasm of the predator. Panel (B) is enlarged from panel (A) to show details of the attachment and tubular tether. The attachment site is indicated by black arrows and shows that the rims of the membrane of Colpodella sp. (ATCC 50594) trophozoite surround the opening created by dissolution of the plasma membrane of the prey. k, kinetoplast; m, mitochondria; n, nucleus; v, vesicle. Scale bars, (A) 1 µm, (B) 500 nm.
Figure 9.
Transmission electron microscopy showing attachment of Colpodella sp. (ATCC 50594) (yellow arrow) to P. caudatus (red arrow). Panel (A) shows attachment of a Colpodella sp. (ATCC 50594) trophozoite to P. caudatus. Contents from the prey flow into the cytoplasm of the predator. Panel (B) is enlarged from panel (A) to show details of the attachment and tubular tether. The attachment site is indicated by black arrows and shows that the rims of the membrane of Colpodella sp. (ATCC 50594) trophozoite surround the opening created by dissolution of the plasma membrane of the prey. k, kinetoplast; m, mitochondria; n, nucleus; v, vesicle. Scale bars, (A) 1 µm, (B) 500 nm.
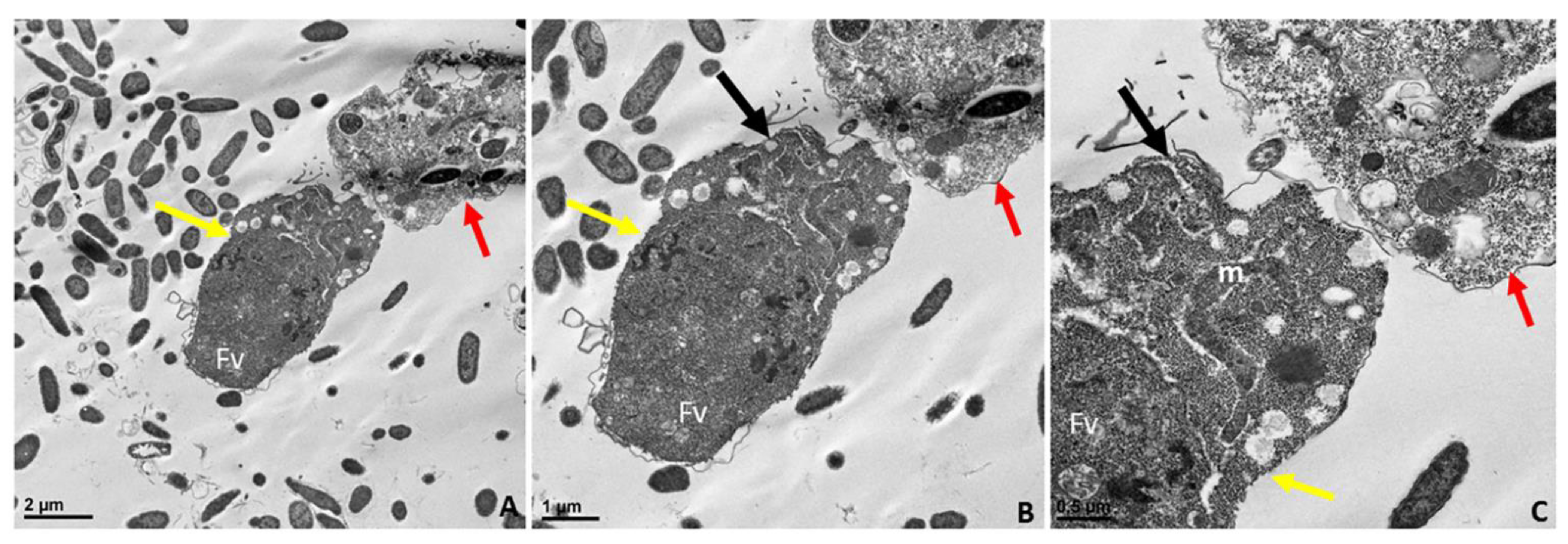
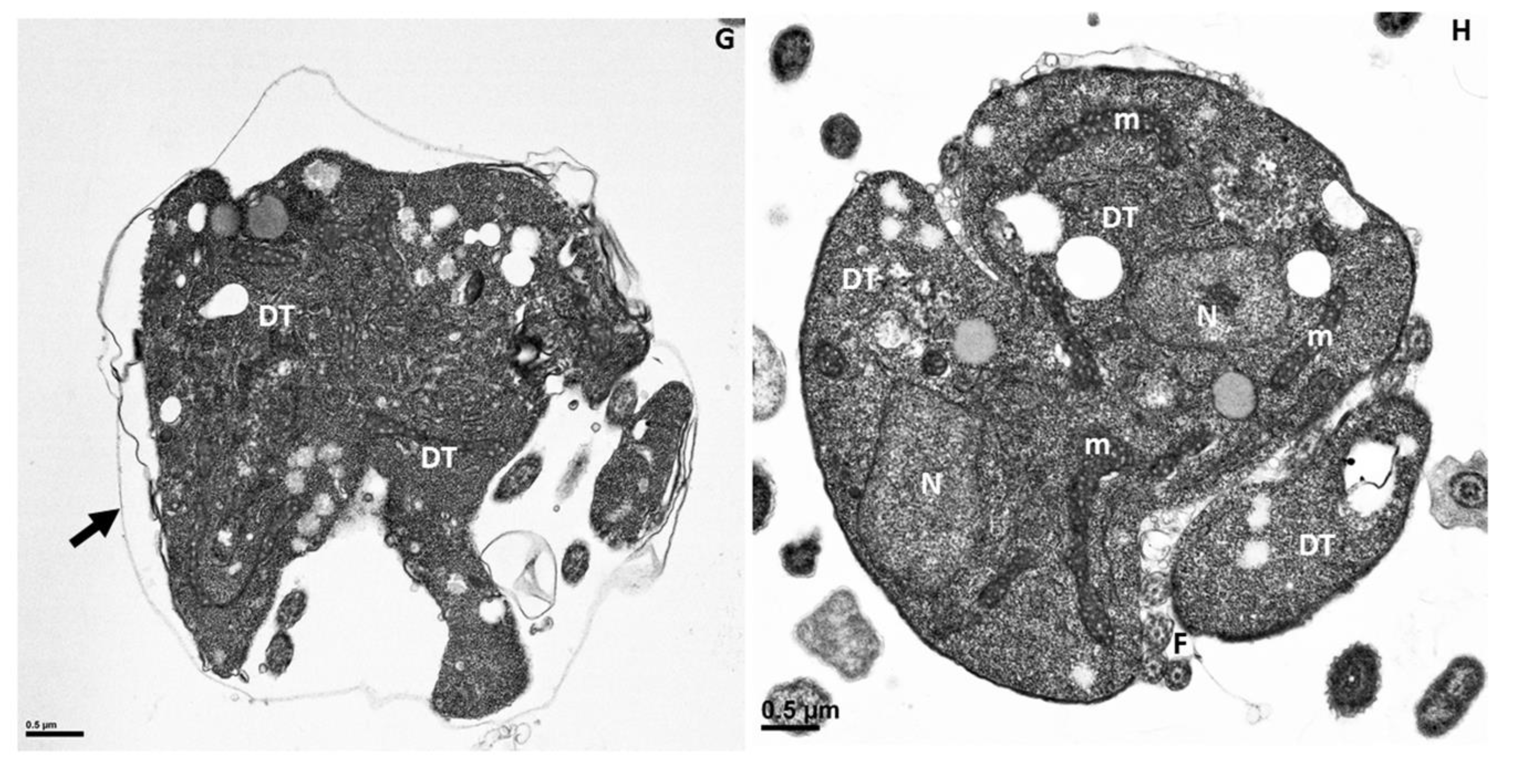
Figure 10.
Transmission electron microscopy of the pre-cyst stage of Colpodella sp. (ATCC 50594) (A). The pre-cyst stage is shown enlarged in panels (B,C) and shows the disintegration of the apical end and loss of the anterior end of the cell after feeding. The food vacuole (Fv) becomes enlarged during myzocytosis before cyst formation. Scale bars, (A) 2 µm, (B) 1 µm, (C) 0.5 µm.
Figure 10.
Transmission electron microscopy of the pre-cyst stage of Colpodella sp. (ATCC 50594) (A). The pre-cyst stage is shown enlarged in panels (B,C) and shows the disintegration of the apical end and loss of the anterior end of the cell after feeding. The food vacuole (Fv) becomes enlarged during myzocytosis before cyst formation. Scale bars, (A) 2 µm, (B) 1 µm, (C) 0.5 µm.
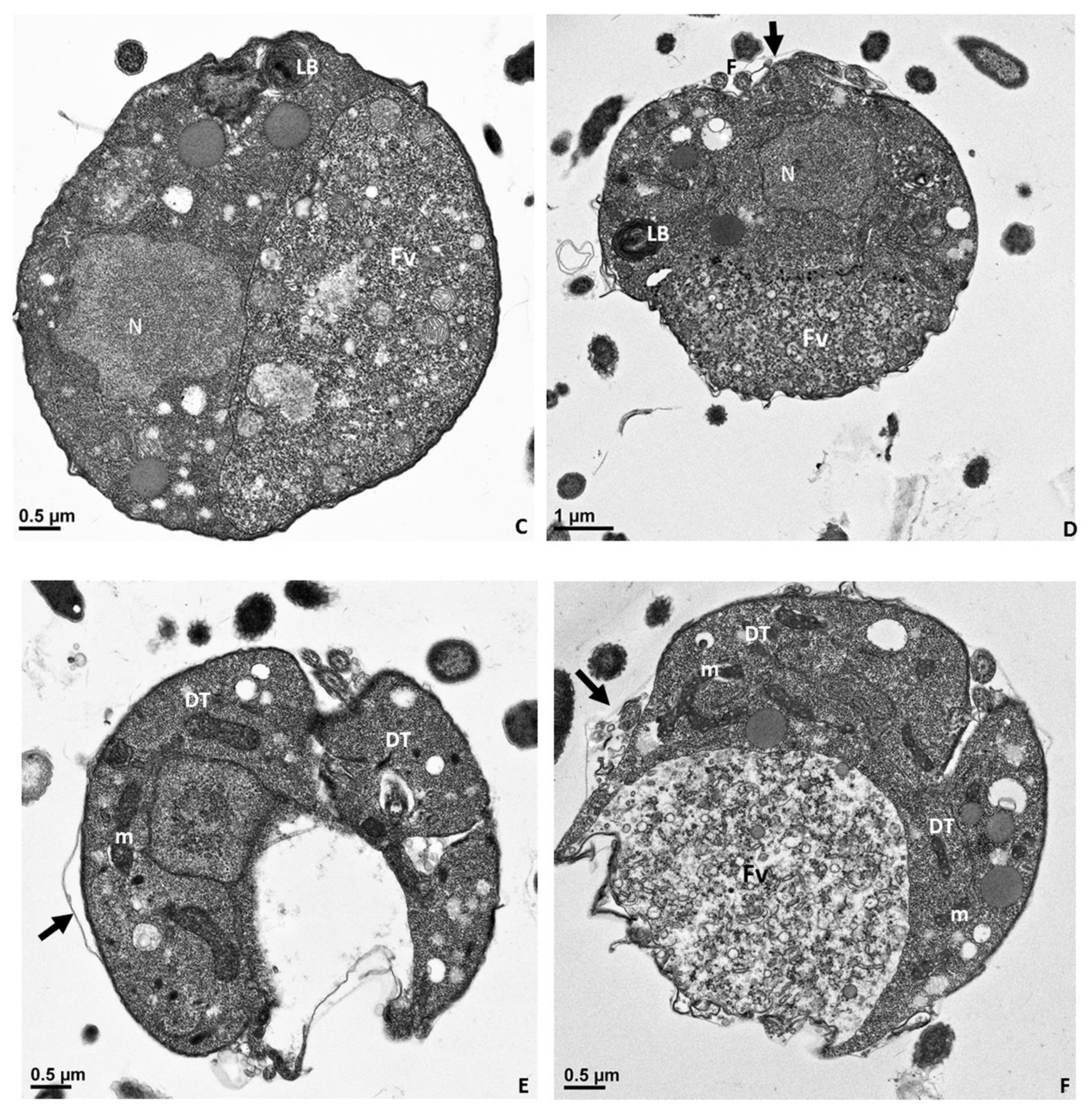
Figure 11.
Transmission electron microscopy of cyst stages of Colpodella sp. (ATCC 50594). Panel (A) shows a young demilune cyst stage followed by single nucleus cyst stages in panels (B–D). Division of the cyst shows the developing trophozoites in panels (E–G) with a food vacuole still present as seen in panel (F). Three developing trophozoites (DT) are shown in panel (H). Flagella (F) were observed in the developing cyst. A thin cyst wall surrounds the cyst (black arrow). DT, developing cyst; F, flagella; Fv, food vacuole; LB, Lamella bodies; m, mitochondria; N, nucleus. Scale bars, (A–C), and (E–H) 0.5 µm, D, 1 µm.
Figure 11.
Transmission electron microscopy of cyst stages of Colpodella sp. (ATCC 50594). Panel (A) shows a young demilune cyst stage followed by single nucleus cyst stages in panels (B–D). Division of the cyst shows the developing trophozoites in panels (E–G) with a food vacuole still present as seen in panel (F). Three developing trophozoites (DT) are shown in panel (H). Flagella (F) were observed in the developing cyst. A thin cyst wall surrounds the cyst (black arrow). DT, developing cyst; F, flagella; Fv, food vacuole; LB, Lamella bodies; m, mitochondria; N, nucleus. Scale bars, (A–C), and (E–H) 0.5 µm, D, 1 µm.
Figure 12.
Actin-green staining of Colpodella sp. (ATCC 50594) (yellow arrow) and P. caudatus (red arrow) trophozoites. Actin is distributed in the cytoskeleton of both protists. Scale bar, 5 µm.
Figure 12.
Actin-green staining of Colpodella sp. (ATCC 50594) (yellow arrow) and P. caudatus (red arrow) trophozoites. Actin is distributed in the cytoskeleton of both protists. Scale bar, 5 µm.
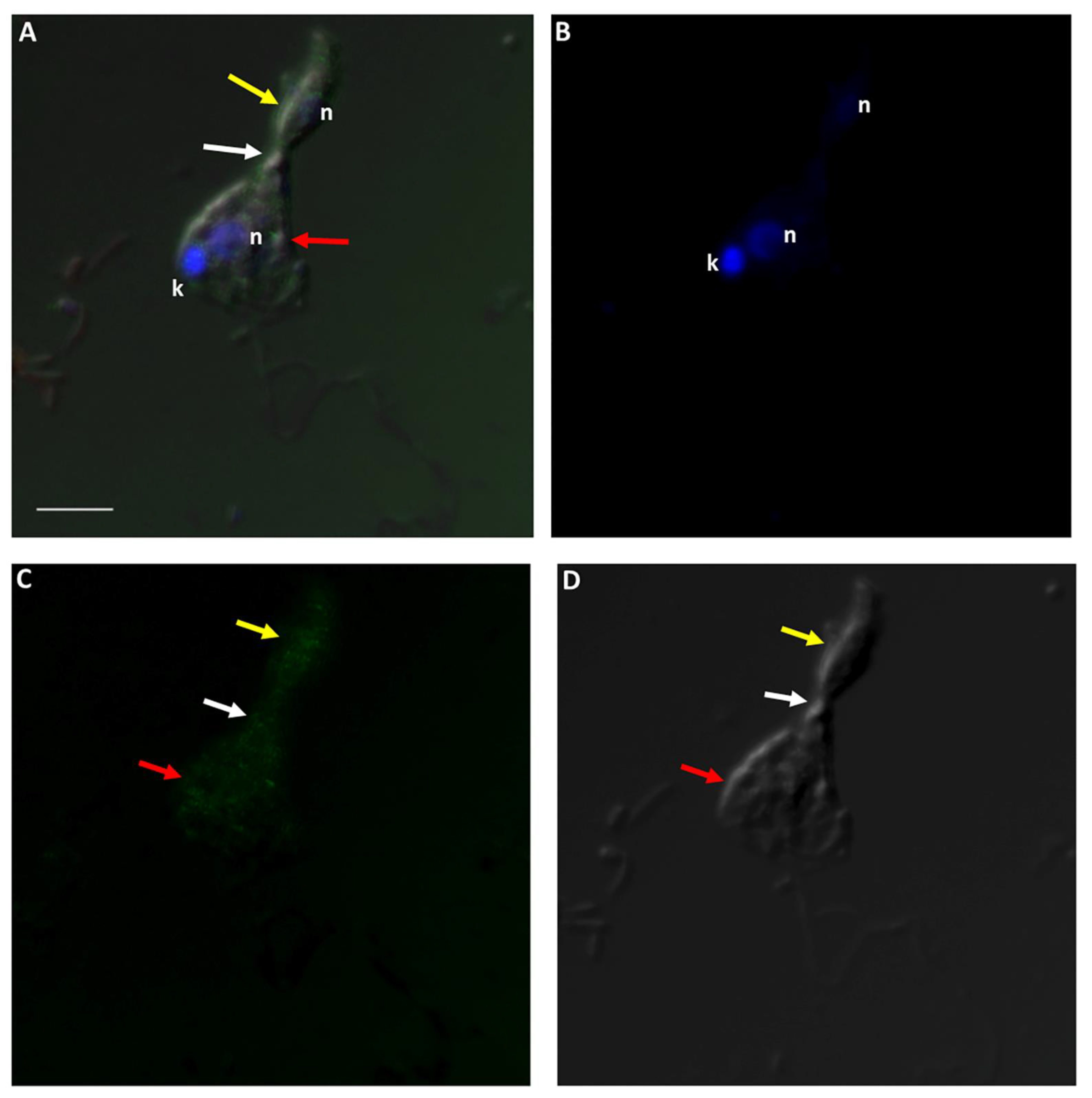
Figure 13.
Actin-green staining of Colpodella sp. (ATCC 50594) (yellow arrow) and P. caudatus (red arrow) trophozoites for confocal microscopy and differential interference contrast (DIC) microscopy. Pre-cyst stage of Colpodella sp. (ATCC 50594) was identified with an enlarged food vacuole. n, nucleus; k, kinetoplast. (A). Overlay of actin-green, DIC, DAPI, and RhopH3 staining. (B). DAPI staining, (C). Actin-green stain, (D). DIC. Scale bar, 10 µm.
Figure 13.
Actin-green staining of Colpodella sp. (ATCC 50594) (yellow arrow) and P. caudatus (red arrow) trophozoites for confocal microscopy and differential interference contrast (DIC) microscopy. Pre-cyst stage of Colpodella sp. (ATCC 50594) was identified with an enlarged food vacuole. n, nucleus; k, kinetoplast. (A). Overlay of actin-green, DIC, DAPI, and RhopH3 staining. (B). DAPI staining, (C). Actin-green stain, (D). DIC. Scale bar, 10 µm.
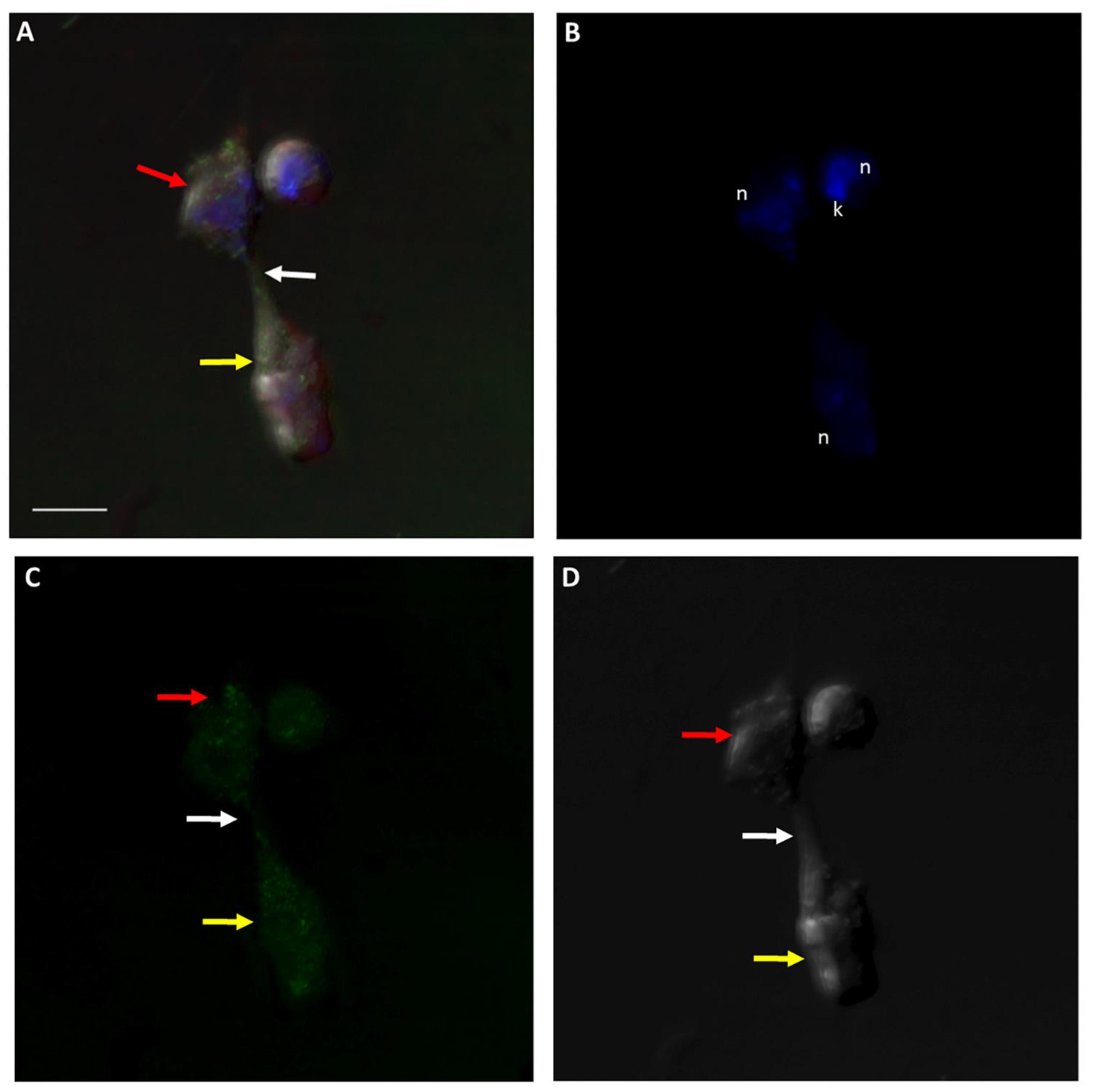
Figure 14.
Actin-green staining of Colpodella sp. (ATCC 50594) (yellow arrow) and P. caudatus (red arrow) trophozoites in myzocytosis for confocal microscopy and differential interference contrast (DIC) microscopy. Actin green staining is distributed in the cytoskeleton of both protists and also in the area of the tubular tether (white arrow). n, nucleus; k, kinetoplast. (A). Overlay of actin green, DIC, DAPI, and RhopH3 staining, (B). DAPI staining, (C). Actin green staining, (D). DIC. Scale bar, 5 µm.
Figure 14.
Actin-green staining of Colpodella sp. (ATCC 50594) (yellow arrow) and P. caudatus (red arrow) trophozoites in myzocytosis for confocal microscopy and differential interference contrast (DIC) microscopy. Actin green staining is distributed in the cytoskeleton of both protists and also in the area of the tubular tether (white arrow). n, nucleus; k, kinetoplast. (A). Overlay of actin green, DIC, DAPI, and RhopH3 staining, (B). DAPI staining, (C). Actin green staining, (D). DIC. Scale bar, 5 µm.
Figure 15.
Actin-green staining of Colpodella sp. (ATCC 50594) (yellow arrow) and P. caudatus (red arrow) trophozoites in myzocytosis for confocal microscopy and differential interference contrast (DIC) microscopy. Actin green staining is distributed in the cytoskeleton of both predator and prey, in the area of the tubular tether (white arrow), and in the cyst of P. caudatus. n, nucleus; k, kinetoplast. (A). Overlay of actin green, DIC, DAPI, and RhopH3 staining. (B). DAPI staining, (C). Actin-green staining, (D). DIC. Scale bar, 5 µm.
Figure 15.
Actin-green staining of Colpodella sp. (ATCC 50594) (yellow arrow) and P. caudatus (red arrow) trophozoites in myzocytosis for confocal microscopy and differential interference contrast (DIC) microscopy. Actin green staining is distributed in the cytoskeleton of both predator and prey, in the area of the tubular tether (white arrow), and in the cyst of P. caudatus. n, nucleus; k, kinetoplast. (A). Overlay of actin green, DIC, DAPI, and RhopH3 staining. (B). DAPI staining, (C). Actin-green staining, (D). DIC. Scale bar, 5 µm.
Figure 16.
Giemsa staining of CytoD-treated cells from diprotist culture. Morphological distortions were observed in both trophozoites of Colpodella sp. (ATCC 50594) (yellow arrows) and P. caudatus (red arrows). Distorted tethers stretched between predator (yellow arrow) and prey (red arrow) are shown in panels (A–G). In some distorted tethers, round grainy material was observed in the tubular tethers (white arrows, panels (A,G)). Black arrows identify the tubular tethers. Trophozoites in myzocytosis were also observed with “normal” morphology as shown in panels (H,I). Kinetoplast (k) and nucleus (n) of P. caudatus appear to be stretched apart (panel (J)). Scale bars, 10 µm.
Figure 16.
Giemsa staining of CytoD-treated cells from diprotist culture. Morphological distortions were observed in both trophozoites of Colpodella sp. (ATCC 50594) (yellow arrows) and P. caudatus (red arrows). Distorted tethers stretched between predator (yellow arrow) and prey (red arrow) are shown in panels (A–G). In some distorted tethers, round grainy material was observed in the tubular tethers (white arrows, panels (A,G)). Black arrows identify the tubular tethers. Trophozoites in myzocytosis were also observed with “normal” morphology as shown in panels (H,I). Kinetoplast (k) and nucleus (n) of P. caudatus appear to be stretched apart (panel (J)). Scale bars, 10 µm.
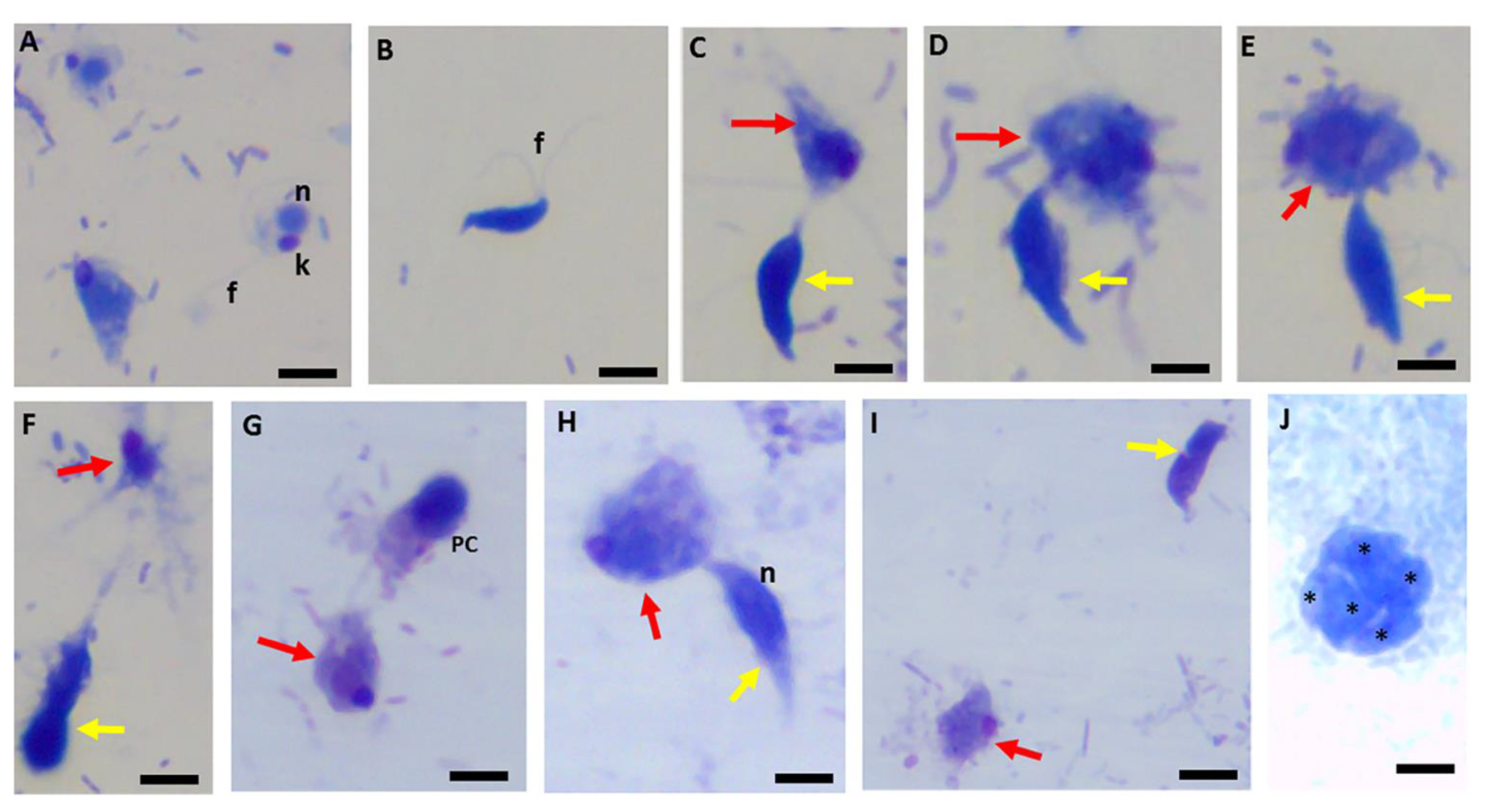
Figure 17.
Untreated controls (panels (A–F)) show normal morphologies of P. caudatus trophozoites (panel (A)) and Colpodella sp. (ATCC 50594) trophozoite (panel (B)). Yellow arrows indicate Colpodella sp. (ATCC 50594), and red arrows indicate Parabodo caudatus. DMSO-treated controls in panel (H) show a trophozoite in myzocytosis. Panel (G) shows a precyst (PC) of Colpodella sp. (ATCC 50594) and trophozoite of P. caudatus (red arrows). Panel (H) shows a trophozoite of Colpodella sp. (ATCC 50594) (yellow arrow) in myzocytosis with P. caudatus (red arrow). Panel (I) shows trophozoites of Colpodella sp. (ATCC 50594) (yellow arrow) and P. caudatus (red arrow). Panel (J) shows a mature cyst of Colpodella sp. (ATCC 50594) with five juveniles (black asterisks). n, nucleus; k, kinetoplast; f, flagella. Scale bars, 10 µm.
Figure 17.
Untreated controls (panels (A–F)) show normal morphologies of P. caudatus trophozoites (panel (A)) and Colpodella sp. (ATCC 50594) trophozoite (panel (B)). Yellow arrows indicate Colpodella sp. (ATCC 50594), and red arrows indicate Parabodo caudatus. DMSO-treated controls in panel (H) show a trophozoite in myzocytosis. Panel (G) shows a precyst (PC) of Colpodella sp. (ATCC 50594) and trophozoite of P. caudatus (red arrows). Panel (H) shows a trophozoite of Colpodella sp. (ATCC 50594) (yellow arrow) in myzocytosis with P. caudatus (red arrow). Panel (I) shows trophozoites of Colpodella sp. (ATCC 50594) (yellow arrow) and P. caudatus (red arrow). Panel (J) shows a mature cyst of Colpodella sp. (ATCC 50594) with five juveniles (black asterisks). n, nucleus; k, kinetoplast; f, flagella. Scale bars, 10 µm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



















