
More Insights about the Efficacy of Copper Ion Treatment on Mycobacterium avium subsp. paratuberculosis (MAP): A Clue for the Observed Tolerance


 ,
,
Abstract
:1. Introduction
2. Results
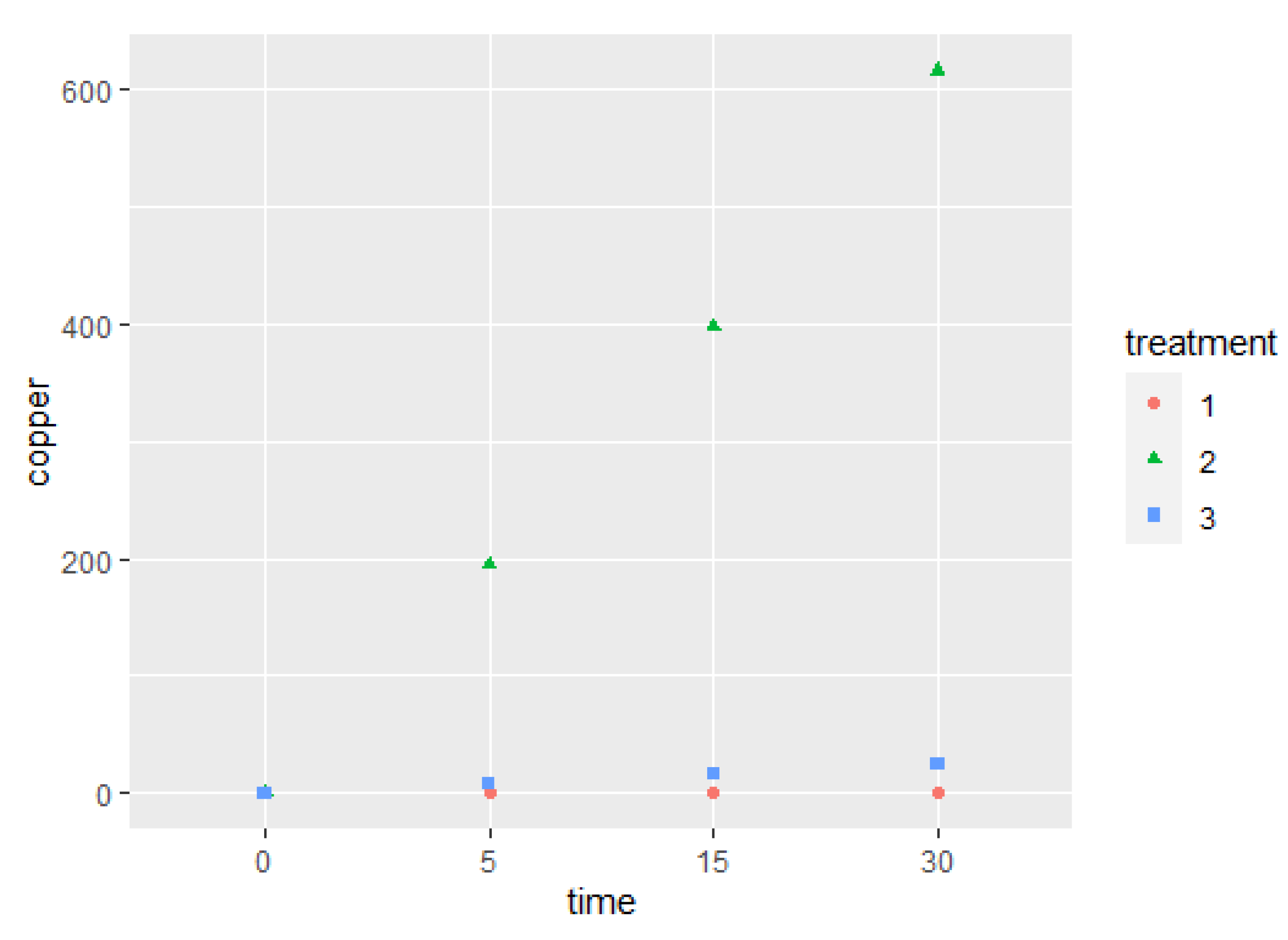
2.1. Evaluation of Copper Concentration
2.2. The Efficacy of Copper Ion Treatment: Estimation of Live Bacterial Load
2.3. The Efficacy of Copper Ion Treatment: Estimation of Bacterial Load by the qPCR Approach
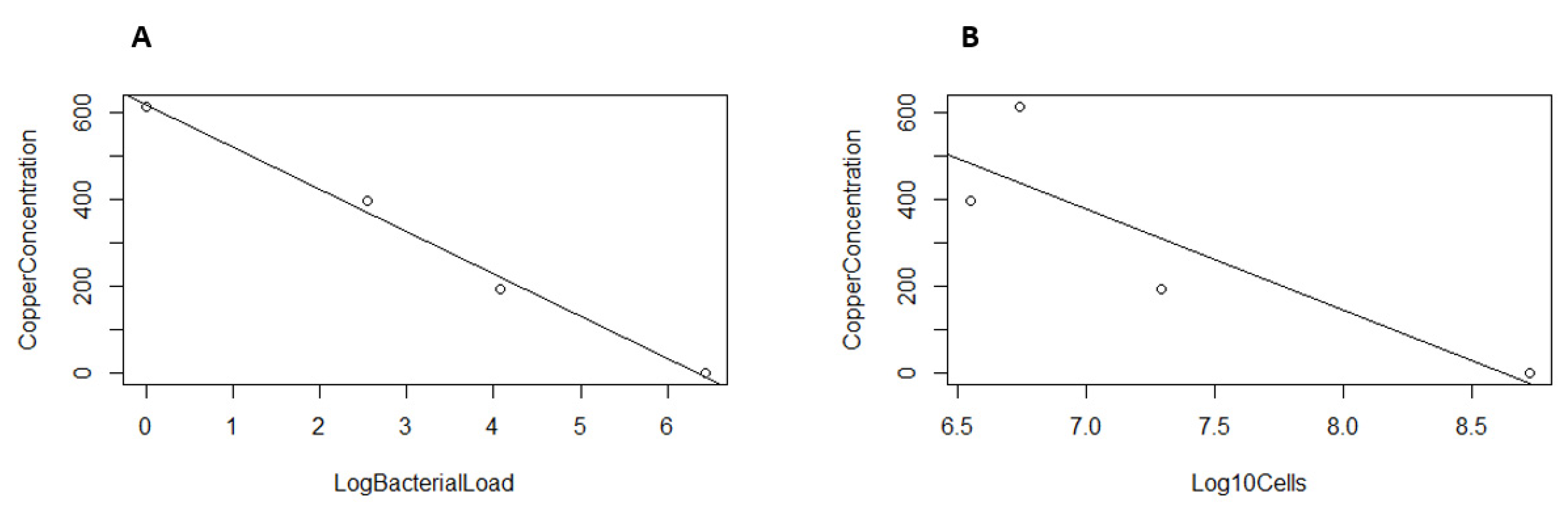
2.4. Correlation between Copper Concentration and Bacterial Load
2.5. Physicochemical Properties of the Treated Buffer
3. Discussion
4. Materials and Methods
4.1. Preparation of Pure MAP Cultures
4.2. Copper Ion Treatment Device
4.3. Estimation of Bacterial Load Using BACTEC–MGIT 960 Culture after the Application of Copper Ion Treatment
4.4. Estimation of Bacterial Load Using qPCR after the Application of Copper Ion Treatment
4.5. Evaluation of the Change in Physicochemical Composition after the Application of Copper Ion Treatment
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Linder, M.C.; Hazegh-Azam, M. Copper biochemistry and molecular biology. Am. J. Clin. Nutr. 1996, 63, 797S–811S. [Google Scholar] [PubMed]
- Dollwet, H.H.; Sorenson, J.R. Historic uses of copper compounds in medicine. Trace Elem. Biol. 1985, 2, 80–87. [Google Scholar]
- Grass, G.; Rensing, C.; Solioz, M. Metallic copper as an antimicrobial surface. Appl Environ Microbiol 2011, 77, 1541–1547. [Google Scholar] [CrossRef] [Green Version]
- Borkow, G.; Gabbay, J. Copper as a biocidal tool. Curr. Med. Chem. 2005, 12, 2163–2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilks, S.A.; Michels, H.; Keevil, C.W. Survival of Escherichia coli O157 on a range of metal surfaces. Int. J. Food Microbiol. 2005, 105, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Noyce, J.O.; Michels, H.; Keevil, C.W. Potential use of copper surfaces to reduce survival of epidemic meticillin resistant Staphylococcus aureus in the healthcare environment. J. Hosp. Infect. 2006, 63, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Warnes, S.L.; Green, S.M.; Michels, H.T.; Keevil, C.W. Biocidal efficacy of copper alloys against pathogenic Enterococci involves degradation of genomic and plasmid DNAs. Appl. Environ. Microbiol. 2010, 76, 5390–5401. [Google Scholar] [CrossRef] [Green Version]
- Espirito Santo, C.; Lam, E.W.; Elowsky, C.G.; Quaranta, D.; Domaille, D.W.; Chang, C.J.; Grass, G. Bacterial killing by dry metallic copper surfaces. Appl. Environ. Microbiol. 2011, 77, 794–800. [Google Scholar] [CrossRef] [Green Version]
- Weaver, L.; Michels, H.T.; Keevil, C.W. Survival of Clostridium difficile on copper and steel: Futuristic options for hospital hygiene. J. Hosp. Infect. 2008, 68, 145–151. [Google Scholar] [CrossRef]
- Mehtar, S.; Wiid, I.; Todorov, S.D. The antimicrobial activity of copper and copper alloys against nosocomial pathogens and Mycobacterium tuberculosis isolated from healthcare facilities in the Western Cape: An invitro study. J. Hosp. Infect. 2008, 68, 45–51. [Google Scholar] [CrossRef]
- Souli, M.; Galani, I.; Plachouras, D.; Panagea, T.; Armaganidis, A.; Petrikkos, G.; Giamarellou, H. Antimicrobial activity of copper surfaces against carbapenemase-producing contemporary Gram-negative clinical isolates. J. Antimicrob. Chemother. 2013, 68, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Benhalima, L.; Amri, S.; Bensouilah, M.; Ouzrout, R. Antibacterial effect of copper sulfate against multi-drug resistant nosocomial pathogens isolated from clinical simples. Pak. J. Med. Sci. 2019, 35, 1322–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usman, M.S.; El Zowalaty, M.E.; Shameli, K.; Zainuddin, N.; Salama, M.; Ibrahim, N.A. Synthesis, characterization, and antimicrobial properties of copper nanoparticles. Int. J. Nanomed. 2013, 8, 4467–4479. [Google Scholar]
- Steuer, P.; Avilez, C.; Tejeda, C.; Gonzalez, N.; Ramirez-Reveco, A.; Ulloa, F.; Mella, A.; Grant, I.; Collins, M.T.; Salgado, M. In Vitro inactivation of Mycobacterium avium subsp. paratuberculosis (MAP) by use of copper ions. BMC Microbiol. 2018, 18, 172. [Google Scholar]
- Steuer, P.; Tejeda, C.; Martinez, O.; Ramirez-Reveco, A.; Gonzalez, N.; Grant, I.R.; Foddai, A.C.G.; Collins, M.T.; Salgado, M. Effectiveness of copper ions against Mycobacterium avium subsp. paratuberculosis and bacterial communities in naturally contaminated raw cow’s milk. J. Appl. Microbiol. 2020, 131, 146–154. [Google Scholar]
- Grant, I.R.; Williams, A.G.; Rowe, M.T.; Muir, D.D. Efficacy of various pasteurization time-temperature conditions in combination with homogenization on inactivation of Mycobacterium avium subsp. paratuberculosis in milk. Appl. Environ. Microbiol. 2005, 71, 2853–2861. [Google Scholar] [CrossRef] [Green Version]
- Whan, L.B.; Grant, I.R.; Ball, H.J.; Scott, R.; Rowe, M.T. Bactericidal effect of chlorine on Mycobacterium paratuberculosis in drinking water. Lett. Appl. Microbiol. 2001, 33, 227–231. [Google Scholar] [CrossRef]
- Adékambi, T.; Ben Salah, S.; Khlif, M.; Raoult, D.; Drancourt, M. Survival of environmental mycobacteria in Acanthamoeba polyphaga. Appl. Environ. Microbiol. 2006, 72, 5974–5981. [Google Scholar] [CrossRef] [Green Version]
- Rowe, M.T.; Grant, I.R. Mycobacterium avium subsp. paratuberculosis and its potential survival tactics. Lett. App. Microbiol. 2006, 42, 305–311. [Google Scholar] [CrossRef]
- Harris, N.B.; Barletta, R.G. Mycobacterium avium subsp. paratuberculosis in veterinary medicine. Clin. Microbiol. Rev. 2001, 14, 489–512. [Google Scholar] [CrossRef] [Green Version]
- Sechi, L.A.; Mura, M.; Tanda, F.; Lissia, A.; Solinas, A.; Fadda, G.; Zanetti, S. Identification of Mycobacterium avium subsp. paratuberculosis in Biopsy Specimens from Patients with Crohn’s Disease Identified by In Situ Hybridization. J. Clin. Microbiol. 2001, 39, 4514–4517. [Google Scholar] [PubMed] [Green Version]
- Versoza, M.; Jung, W.; Barabad, M.L.; Ko, S.; Kim, M.; Park, D. Reduction of Escherichia coli using metal plates with the influenced of applied low current and physical barrier of filter layers. Int. J. Environ. Res. Public Health 2019, 16, 3887. [Google Scholar] [CrossRef] [Green Version]
- Hall, C.W.; Mah, T.F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef] [PubMed]
- Steuer, P.; Tejeda, C.; Moroni, M.; Verdugo, C.; Collins, M.T.; Salgado, M. Attempted Control of Paratuberculosis in Dairy Calves by Only Changing the Quality of Milk Fed to Calves. Animals 2021, 11, 2569. [Google Scholar] [CrossRef] [PubMed]
- Hosseiniporgham, S.; Biet, F.; Ganneau, C.; Bannantine, J.P.; Bay, S.; Sechi, L.A. A Comparative Study on the Efficiency of Two Mycobacterium avium subsp. paratuberculosis (MAP)-Derived Lipopeptides of L3P and L5P as Capture Antigens in an In-House Milk ELISA Test. Vaccines 2021, 9, 99. [Google Scholar]
- Bagherian, G.; Chamjangali, M.A.; Evari, H.S.; Ashrafi, M. Determination of copper (II) by flame atomic absorption spectrometry after its perconcentration by a highly selective and environmentally friendly dispersive liquid—liquid microextraction technique. J. Anal. Sci. Technol. 2019, 10, 3. [Google Scholar] [CrossRef]
- Lamont, E.A.; Bannantine, J.P.; Armién, A.; Ariyakumar, D.S.; Sreevatsan, S. Identification and Characterization of a Spore-Like Morphotype in Chronically Starved Mycobacterium avium subsp. paratuberculosis Cultures. PLoS ONE 2012, 7, e30648. [Google Scholar] [CrossRef] [Green Version]
- Solioz, M.; Stoyanov, J.V. Copper homeostasis in Enterococcus hirae. FEMS Microbiol. Rev. 2003, 27, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Wolschendorf, F.; Ackart, D.; Shrestha, T.B.; Hascall-Dove, L.; Nolan, S.; Lamichhane, G.; Wang, Y.; Bossmann, S.H.; Basaraba, R.J.; Niederweis, M. Copper resistance is essential for virulence of Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2011, 108, 1621–1626. [Google Scholar]
- Shi, X.; Darwin, K.H. Copper homeostasis in Mycobacterium tuberculosis. Metallomics 2015, 7, 929–934. [Google Scholar] [CrossRef] [Green Version]
- Salgado, M.; Alfaro, M.; Salazar, F.; Troncoso, E.; Mitchell, R.M.; Ramirez, L.; Niguil, A.; Zamorano, P.; Collins, M.T. Effect of soil slope on appearance of Mycobacterium avium subsp. paratuberculosis in water running off grassland soil after contaminated slurry application. Appl. Environ. Microbiol. 2013, 79, 3544–3552. [Google Scholar] [PubMed] [Green Version]
- Reyes-Jara, A.; Cordero, N.; Aguirre, J.; Troncoso, M.; Figueroa, G. Antibacterial Effect of Copper on Microorganisms Isolated from Bovine Mastitis. Front. Microbiol. 2016, 7, 626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herthnek, D.; Englund, S.; Willemsen, P.T.J.; Bölske, G. Sensitive detection of Mycobacterium avium subsp. paratuberculosis in bovine semen by real-time PCR. J. Appl. Microbiol. 2006, 100, 1095–1102. [Google Scholar]
- Sweeney, R.W.; Whitlock, R.H.; McAdams, S.C. Comparison of three DNA preparation methods for real-time polymerase chain reaction confirmation of Mycobacterium avium subsp. paratuberculosis growth in an automated broth culture system. J. Vet. Diagn. Investig. 2006, 18, 587–590. [Google Scholar]
- Salgado, M.; Verdugo, C.; Heuer, C.; Castillo, P.; Zamorano, P. A novel low-cost method for Mycobacterium avium subsp. paratuberculosis DNA extraction from an automated broth culture system for a real time PCR confirmation. J. Vet. Sci. 2013, 15, 233–239. [Google Scholar]
- Shin, S.J.; Han, J.H.; Manning, E.J.B.; Collins, M.T. Rapid and reliable method for quantification of Mycobacterium paratuberculosis by use of the BACTEC MGIT 960 System. J. Clin. Microbiol. 2007, 45, 1941–1948. [Google Scholar] [CrossRef] [Green Version]
- Borkow, G.; Gabbay, J. Copper, An Ancient Remedy Returning to Fight Microbial, Fungal and Viral Infections. Curr. Med. Chem. 2009, 3, 272–278. [Google Scholar]
- Hans, M.; Mathews, S.; Mücklich, F.; Solioz, M. Physicochemical properties of copper important for its antibacterial activity and development of a unified model. Biointerphases 2016, 11, 018902. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Wang, H.; Zhang, X.; Qian, Y. Effect of Direct Electric Current on the Cell Surface Properties of Phenol-Degrading Bacteria. Appl. Environ. Microbiol. 2005, 423–427. [Google Scholar] [CrossRef] [Green Version]
- Weaver, J.C.; Chizmadzhev, Y.A. Theory of electroporation: A review. Bioelectrochem. Bioenerg. 1996, 41, 135–160. [Google Scholar] [CrossRef]
- Dreesa, K.P.; Abbaszadegan, M.; Maiera, R.M. Comparative electrochemical inactivation of bacteria and bacteriophage. Water Res. 2003, 37, 2291–2300. [Google Scholar] [CrossRef]
- Wells, S.J.; Wagner, B.A. Herd-level risk factors for infection with Mycobacterium paratuberculosis in US dairies and association between familiarity of the herd manager with the disease or prior diagnosis of the disease in that herd and use of preventive measures. J. Am. Vet. Med. Assoc. 2000, 216, 1450–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salgado, M.; Alfaro, M.; Salazar, F.; Badilla, X.; Troncoso, E.; Zambrano, A.; Gonzalez, M.; Mitchell, R.M.; Collins, M.T. Application of cattle slurry containing Mycobacterium avium subsp. paratuberculosis (MAP) to grassland soil and its effect on the relationship between MAP and free-living amoeba. Vet. Microbiol. 2015, 175, 26–34. [Google Scholar]
- Steuer, P.; Collado, B.; Avilez, C.; Tejeda, C.; Soto, J.P.; Salgado, M. Is the transmission of Mycobacterium avium subspecies paratuberculosis (MAP) infection through milk intended to feed calves an overlooked item in paratuberculosis control programs? Trop. Anim. Health Prod. 2019, 52, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Dzieciol, M.; Volgger, P.; Khol, J.; Baumgartner, W.; Wagner, M.; Hein, I. A novel real-time PCR assay for specific detection and quantification of Mycobacterium avium subsp. paratuberculosis in milk with the inherent possibility of differentiation between viable and dead cells. BMC Res. Notes 2010, 3, 251. [Google Scholar]
- Crawley, M.J. The R Book, 2nd ed.; Imperial College London: Silwood Park, UK, 2013; Available online: http://www.bio.ic.ac.uk/research/mjcraw/therbook/index.htm (accessed on 10 January 2022).



{kind=link}
{kind=link}
| Cu Exposure Time (min) | TTD Days (Mean ± SD) | Log10 Cells (Mean ± SD) | ||||
|---|---|---|---|---|---|---|
| 106 | 104 | 102 | 106 | 104 | 102 | |
| MAP Dilution (Cells/mL) (Mean ± SD) | MAP Dilution (Cells/mL) (Mean ± SD) | |||||
| 0 | 0.46 ± 0.00 | 0.67 ± 0.00 | 1.05 ± 0.01 | 8.72 ± 0.00 | 8.61 ± 0.00 | 8.42 ± 0.01 |
| 5 | 3.63 ± 3.35 | 8.34 ± 1.59 | 15.4 ± 0.02 | 7.29 ± 1.45 | 5.49 ± 0.50 | 3.74 ± 0.01 |
| 15 | 5.25 ± 1.06 | 7.61 ± 1.85 | 16.6 ± 4.16 | 6.55 ± 0.40 | 5.73 ± 0.60 | 3.57 ± 0.74 |
| 30 | 4.75 ± 0.35 | 8.80 ± 0.29 | 16.1 ± 4.74 | 6.74 ± 0.13 | 5.34 ± 0.09 | 3.68 ± 0.88 |
| MAP | Load | ||
|---|---|---|---|
| Cu Exposure Time (min) | No TT | Complete Cu TT | Cu TT w/o E |
| (mean ± SD) | (mean ± SD) | (mean ± SD) | |
| 0 | 3.34 × 106 ± 340,293 a | 2.71 × 106 ± 74,833 a | 4.97 × 106 ± 79,302 a |
| 5 | 3.46 × 106 ± 43,204 a | 1.20 × 104 ± 499 b | 5.18 × 105 ± 10,801 a |
| 15 | 3.87 × 106 ± 101,980 a | 3.46 × 102 ± 4.92 c | 3.43 × 105 ± 18,018 a |
| 30 | 3.8 × 106 ± 82,596 a | 0.00 × 100 ± 0.00 c | 7.82 × 104 ± 589 a |
| Cu Exposure Time (min) | pH with No TT | pH with Complete Cu TT | pH with Cu TT w/o E | EC with No TT | EC with Complete Cu TT | EC with Cu TT w/o E | [O2] with No TT | [O2] with Complete Cu TT | [O2] with Cu TT w/o E |
|---|---|---|---|---|---|---|---|---|---|
| (mean ± SD) | (mean ± SD) | (mean ± SD) | (mean ± SD) | (mean ± SD) | (mean ± SD) | (mean ± SD) | (mean ± SD) | (mean ± SD) | |
| 0 | 7.5 ± 0.04 | 7.5 ± 0.04 | 7.5 ± 0.07 | 4.7 ± 0.09 | 4.7 ± 0.08 | 4.8 ± 0.02 | 7.3 ± 0.12 | 7.4 ± 0.39 | 7.6 ± 0.02 |
| 5 | 7.5 ± 0.02 | 11.0 ± 0,09 | 7.4 ± 0.02 | 4.7 ± 0.06 | 4.8 ± 0.07 | 4.9 ± 0.03 | 7.8 ± 0.18 | 4.3 ± 1.38 | 7.5 ± 0.03 |
| 15 | 7.6 ± 0.04 | 11.8 ± 0.09 | 7.2 ± 0.02 | 4.7 ± 0.07 | 5.5 ± 0.08 | 4.9 ± 0.03 | 7.6 ± 0.21 | 3.3 ± 1.50 | 7.3 ± 0.03 |
| 30 | 7.6 ± 0.04 | 11.9 ± 0.04 | 7.2 ± 0.03 | 4.7 ± 0.04 | 5.8 ± 0.28 | 4.9 ± 0.02 | 7.5 ± 0.21 | 2.5 ± 1.31 | 7.0 ± 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tejeda, C.; Steuer, P.; Villegas, M.; Reyes-Jara, A.; Iranzo, E.C.; Umaña, R.; Salgado, M. More Insights about the Efficacy of Copper Ion Treatment on Mycobacterium avium subsp. paratuberculosis (MAP): A Clue for the Observed Tolerance. Pathogens 2022, 11, 272. https://doi.org/10.3390/pathogens11020272
Tejeda C, Steuer P, Villegas M, Reyes-Jara A, Iranzo EC, Umaña R, Salgado M. More Insights about the Efficacy of Copper Ion Treatment on Mycobacterium avium subsp. paratuberculosis (MAP): A Clue for the Observed Tolerance. Pathogens. 2022; 11(2):272. https://doi.org/10.3390/pathogens11020272
Chicago/Turabian StyleTejeda, Carlos, Pamela Steuer, Marcela Villegas, Angelica Reyes-Jara, Esperanza C. Iranzo, Reydoret Umaña, and Miguel Salgado. 2022. "More Insights about the Efficacy of Copper Ion Treatment on Mycobacterium avium subsp. paratuberculosis (MAP): A Clue for the Observed Tolerance" Pathogens 11, no. 2: 272. https://doi.org/10.3390/pathogens11020272