

Progress of the COVID-19: Persistence, Effectiveness, and Immune Escape of the Neutralizing Antibody in Convalescent Serum
 ,
,
Abstract
:1. Introduction
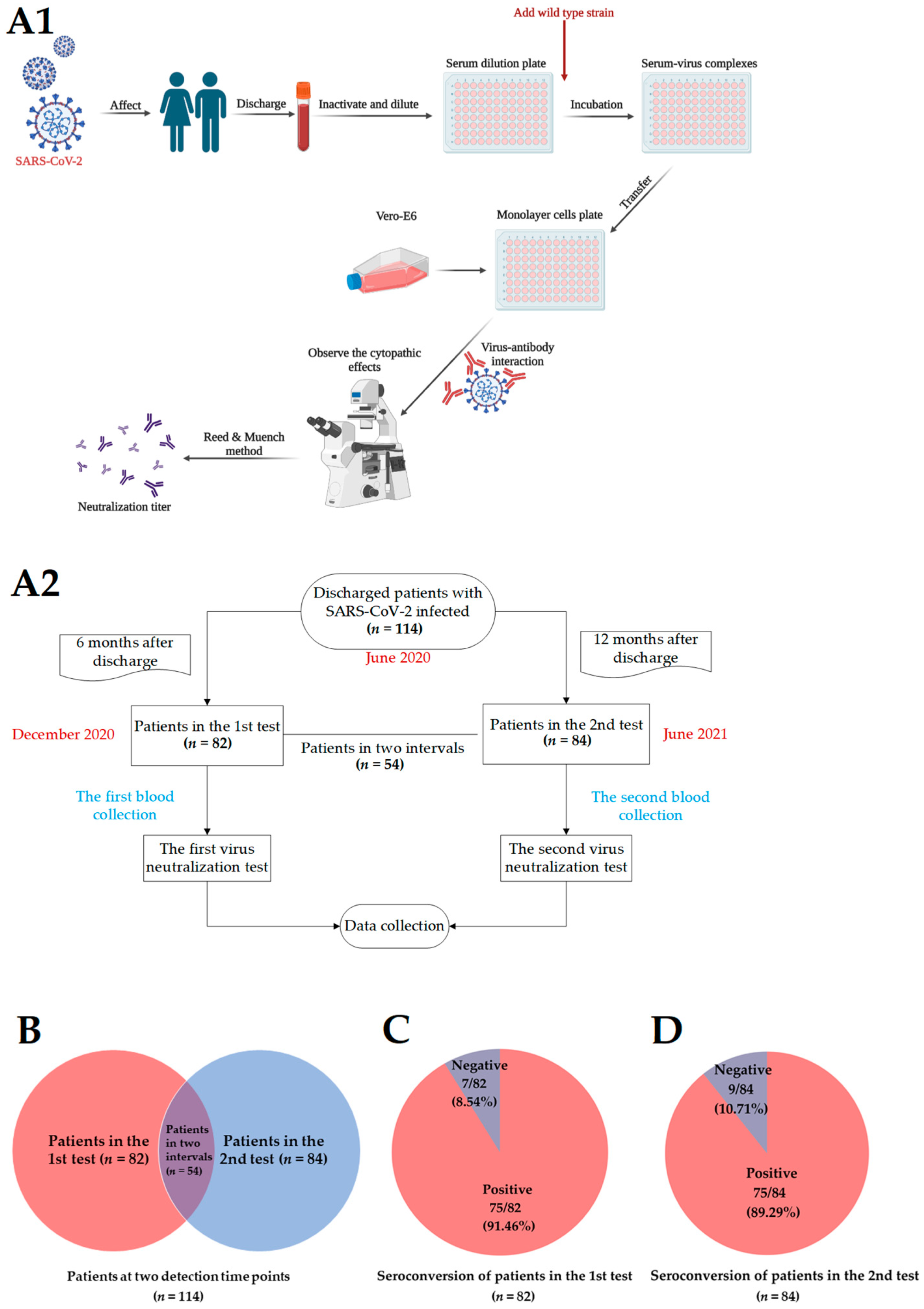
2. Materials and Methods
2.1. Ethics Approval Statement
2.2. Serum Specimen
2.3. Vero-E6 Cells
2.4. SARS-CoV-2 Isolates
2.5. Virus Neutralization Test
2.6. Statistical Analyses
3. Results
3.1. Basic Information on Patients with COVID-19
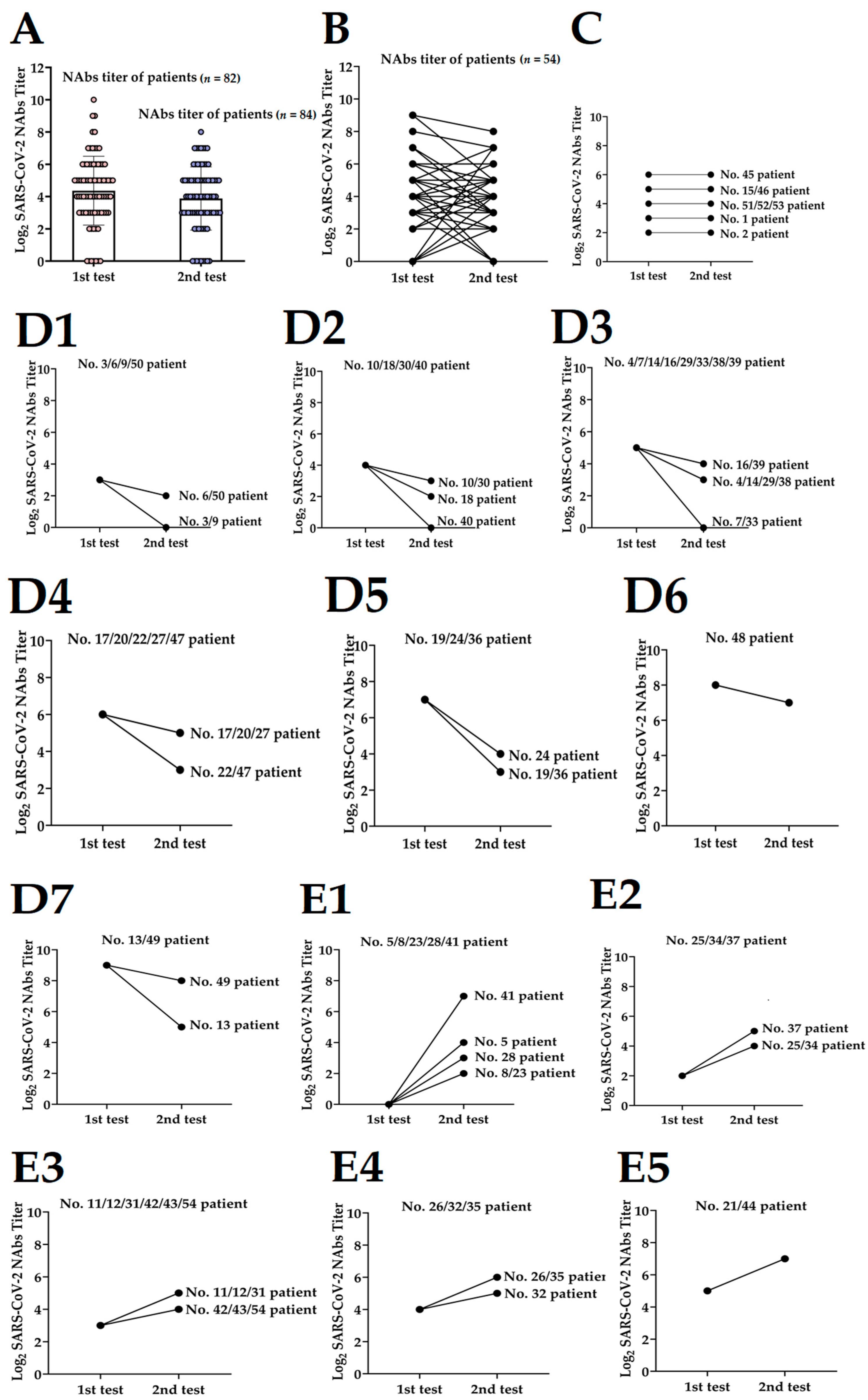
3.2. Dynamics of NAbs Level to Discharged Patients
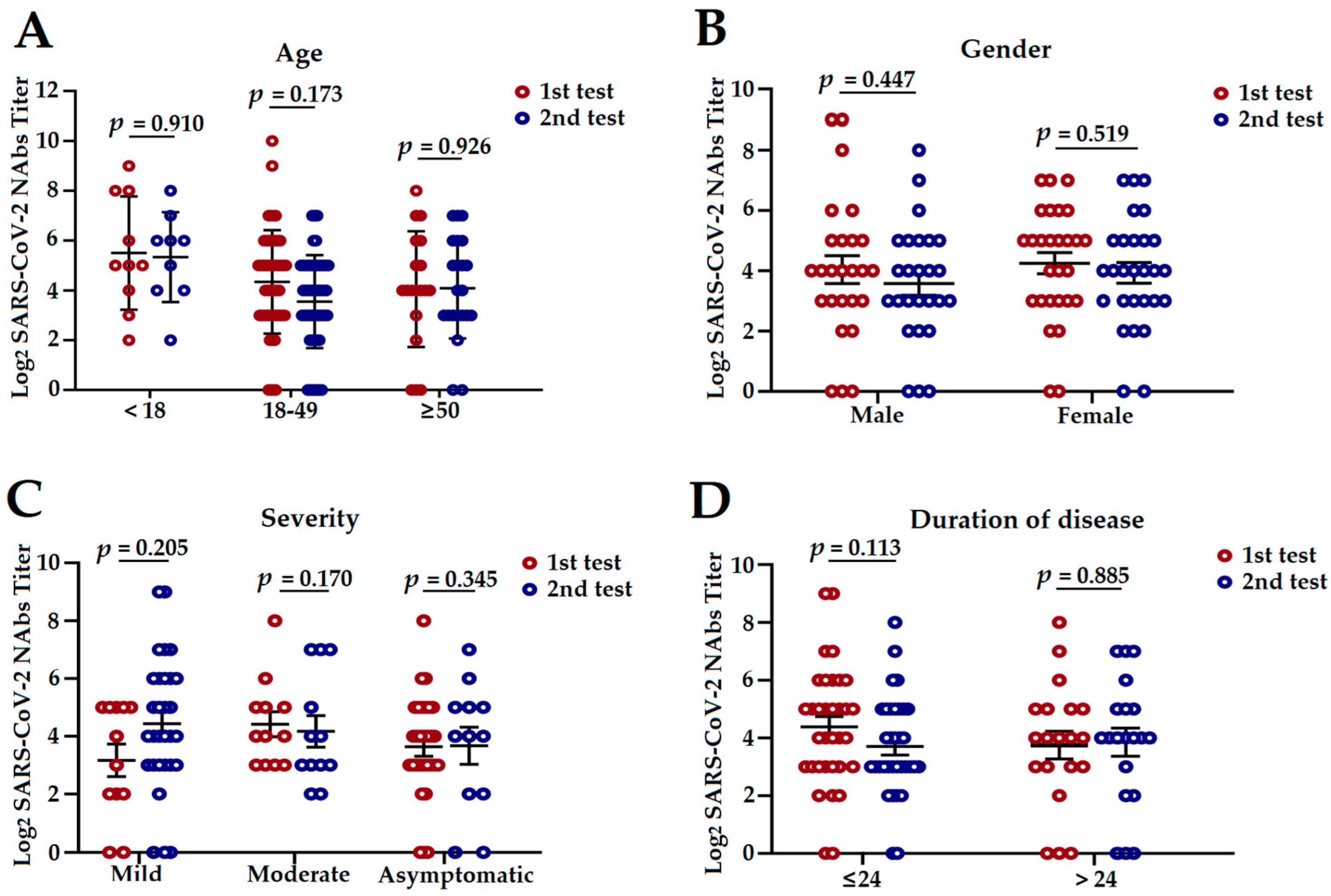
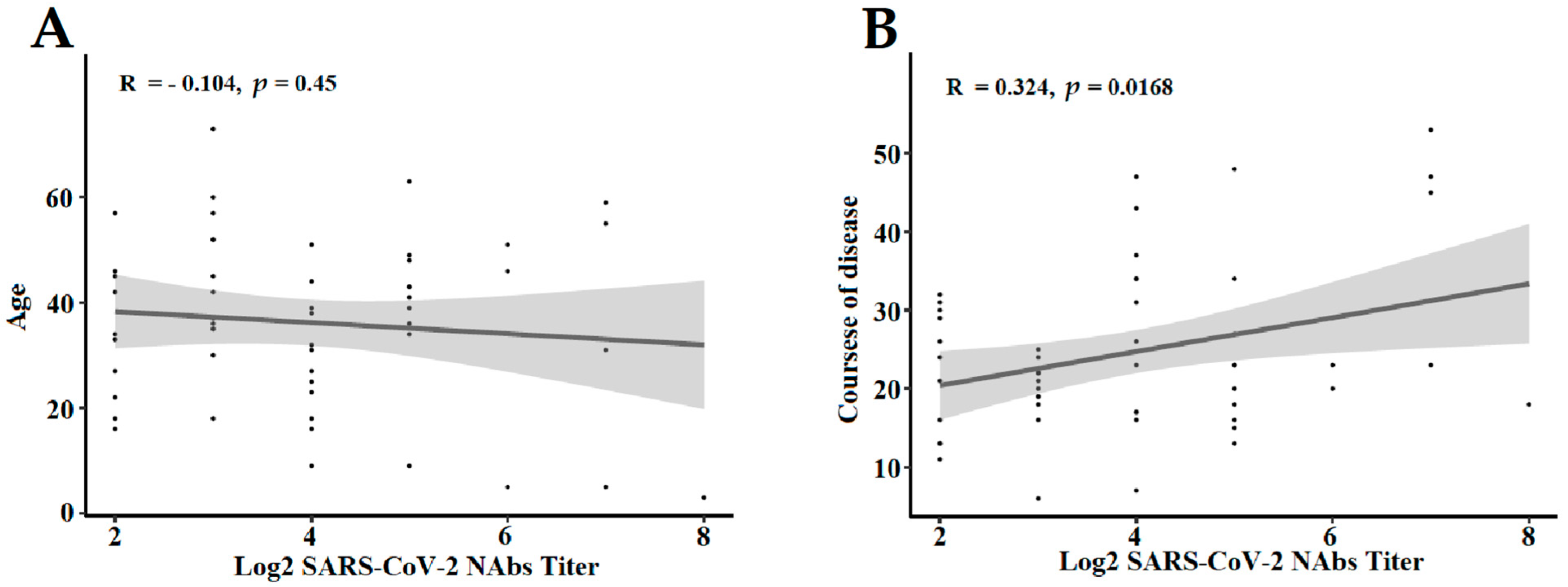
3.3. The Influencing Factors of NAbs Level in COVID-19 Convalescent Patients
3.4. The NAbs against Other Variants in COVID-19 Convalescent Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- V’Kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Hirabara, S.M.; Serdan, T.D.A.; Gorjao, R.; Masi, L.N.; Pithon-Curi, T.C.; Covas, D.T.; Curi, R.; Durigon, E.L. SARS-CoV-2 Variants: Differences and Potential of Immune Evasion. Front. Cell. Infect. Microbiol. 2021, 11, 781429. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. World Health Organization (WHO)/Home/Diseases/ Coronavirus Disease (COVID-19)/Number at a Glance (last updated: 13 April 2022). Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 (accessed on 14 April 2022).
- Lumley, S.F.; O’Donnell, D.; Stoesser, N.E.; Matthews, P.C.; Howarth, A.; Hatch, S.B.; Marsden, B.D.; Cox, S.; James, T.; Warren, F.; et al. Antibody Status and Incidence of SARS-CoV-2 Infection in Health Care Workers. N. Engl. J. Med. 2021, 384, 533–540. [Google Scholar] [CrossRef]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef]
- Muecksch, F.; Weisblum, Y.; Barnes, C.O.; Schmidt, F.; Schaefer-Babajew, D.; Wang, Z.J.; Lorenzi, J.C.C.; Flyak, A.I.; Delaitsch, A.T.; Huey-Tubman, K.E.; et al. Affinity maturation of SARS-CoV-2 neutralizing antibodies confers potency, breadth, and resilience to viral escape mutations. Immunity 2021, 54, 1853–1868.e7. [Google Scholar] [CrossRef] [PubMed]
- Matusali, G.; Colavita, F.; Lapa, D.; Meschi, S.; Bordi, L.; Piselli, P.; Galiardini, R.; Corpolongo, A.; Nicastri, E.; Antinori, A.; et al. SARS-CoV-2 Serum Neutralization Assay: A Traditional Tool for a Brand-New Virus. Viruses 2021, 13, 655. [Google Scholar] [CrossRef] [PubMed]
- Gaebler, C.; Wang, Z.; Lorenzi, J.C.C.; Muecksch, F.; Finkin, S.; Tokuyama, M.; Cho, A.; Jankovic, M.; Schaefer-Babojew, D.; Oliveira, T.Y.; et al. Evolution of antibody immunity to SARS-CoV-2. Nature 2021, 591, 639–644. [Google Scholar] [CrossRef]
- Dan, J.M.; Mateus, J.; Kato, Y.; Hastie, K.M.; Yu, E.D.; Faliti, C.E.; Grifon, A.; Ramirez, S.I.; Haupt, S.; Frazier, A.; et al. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef]
- Liu, W.; Fontanet, A.; Zhang, P.H.; Zhan, L.; Xin, Z.T.; Baril, L.; Tang, F.; Lv, H.; Cao, W.C. Two-year prospective study of the humoral immune response of patients with severe acute respiratory syndrome. J. Infect. Dis. 2006, 193, 792–795. [Google Scholar] [CrossRef]
- De Giorgi, V.; West, K.A.; Henning, A.N.; Chen, L.N.; Holbrook, M.R.; Gross, R.; Liang, J.; Postnikova, E.; Trenbeath, J.; Pogue, S. Naturally Acquired SARS-CoV-2 Immunity Persists for Up to 11 Months Following Infection. J. Infect. Dis. 2021, 224, 1294–1304. [Google Scholar] [CrossRef]
- Post, N.; Eddy, D.; Huntley, C.; van Schalkwyk, M.C.I.; Shrotri, M.; Leeman, D.; Rigby, S.; Williams, S.V.; Bermingham, W.H.; Kellam, P. Antibody response to SARS-CoV-2 infection in humans: A systematic review. PLoS ONE 2020, 15, e0244126. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.; Zhang, Y.; Tang, G.; Luo, Y.; Liu, W.; Cheng, C.; Jiang, Y.H.; Xiong, Z.G.; Wu, S.J.; Sun, Z.Y. Immunologic memory to SARS-CoV-2 in convalescent COVID-19 patients at 1 year postinfection. J. Allergy Clin. Immunol. 2021, 148, 1481–1492.e2. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Nie, K.; Li, K.; Hu, Y.; Song, Y.; Kang, M.; Zhang, M.; Deng, X.L.; Yuan, J.; Xu, W.B. Genome Characterization of the First Outbreak of COVID-19 Delta Variant B.1.617.2—Guangzhou City, Guangdong Province, China, May 2021. China CDC Wkly. 2021, 3, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Zheng, Z.; Zhao, X.; Zeng, Q.; Zhou, T.; Guo, Q.; Hu, Y.; Xu, W.B.; Zhang, Z.B.; Li, B.S. An Imported Case and an Infected Close Contact of the Omicron Variant of SARS-CoV-2—Guangdong Province, China, December 13, 2021. China CDC Wkly. 2022, 4, 96–97. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.Y.; Wang, W.B.; Gao, R.D.; Zhou, A.M. Omicron Variant (B.1.1.529) of SARS-CoV-2: Mutation, infectivity, transmission, and vaccine resistance. World J. Clin. Cases 2022, 10, 1–11. [Google Scholar] [CrossRef]
- University S Coronavirus Antiviral & Resistance Database. Available online: https://covdb.stanford.edu/page/susceptibility-data/ (accessed on 2 December 2022).
- Zhang, M. Omicron Now Account for 35 of US Coronavirus Cases. China Daily Global. Available online: https://global.chinadaily.com.cn/a/202112/15/WS61b95f63a310cdd39bc7b7f8.html (accessed on 2 December 2022).
- Huang, C.; Huang, L.; Wang, Y.; Li, X.; Ren, L.; Gu, X.; Kang, L.; Guo, L.; Liu, M.; Zhou, X.; et al. 6-month consequences of COVID-19 in patients discharged from hospital: A cohort study. Lancet 2021, 397, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Battaglia, F.; Terlecky, S.R. Discharged COVID-19 patients testing positive again for SARS-CoV-2 RNA: A minireview of published studies from China. J. Med. Virol. 2021, 93, 262–274. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, H.Y.; Zou, L.R.; Liu, Z.; Liang, L.J.; Peng, X.F.; Zhang, W.; Ke, C.W.; Wu, J. First Isolation and Identification of SARS-CoV-2 in Guangdong Province, China. Chin. J. Virol. 2020, 36, 155–159. (In Chinese) [Google Scholar] [CrossRef]
- Németh, B.; Fasseeh, A.; Molnár, A.; Bitter, I.; Horváth, M.; Kóczián, K.; Götze, Á.; Nagy, B. A systematic review of health economic models and utility estimation methods in schizophrenia. Expert Rev. Pharm. Outcomes Res. 2018, 18, 267–275. [Google Scholar] [CrossRef]
- Marklund, E.; Leach, S.; Axelsson, H.; Nyström, K.; Norder, H.; Bemark, M.; Aneletti, D.; Lundgren, A.N.; Nilsson, S.; Andersson, L.M.; et al. Serum-IgG responses to SARS-CoV-2 after mild and severe COVID-19 infection and analysis of IgG non-responders. PLoS ONE 2020, 15, e0241104. [Google Scholar] [CrossRef]
- Bongen, E.; Lucian, H.; Khatri, A.; Fragiadakis, G.K.; Bjornson, Z.B.; Nolan, G.P.; Utz, P.J.; Khatri, P. Sex Differences in the Blood Transcriptome Identify Robust Changes in Immune Cell Proportions with Aging and Influenza Infection. Cell Rep. 2019, 29, 1961–1973.e4. [Google Scholar] [CrossRef] [PubMed]
- Schurz, H.; Salie, M.; Tromp, G.; Hoal, E.G.; Kinnear, C.J.; Möller, M. The X chromosome and sex-specific effects in infectious disease susceptibility. Hum. Genom. 2019, 13, 2. [Google Scholar] [CrossRef] [PubMed]
- Steeg, L.G.V.; Klein, S.L. SeXX Matters in Infectious Disease Pathogenesis. PLoS Pathog. 2016, 12, e1005374. [Google Scholar]
- Márquez, E.J.; Chung, C.H.; Marches, R.; Rossi, R.J.; Nehar-Belaid, D.; Eroglu, A.; Mellert, D.J.; Kuchel, G.A.; Banchereau, J.; Ucar, D. Sexual-dimorphism in human immune system aging. Nat. Commun. 2020, 11, 751. [Google Scholar] [CrossRef]
- Ober, C.; Loisel, D.A.; Gilad, Y. Sex-specific genetic architecture of human disease. Nat. Rev. Genet. 2008, 9, 911–922. [Google Scholar] [CrossRef]
- Schmiedel, B.J.; Singh, D.; Madrigal, A.; Valdovino-Gonzalez, A.G.; White, B.M.; Zapardiel-Gonzalo, J.; Ha, B.; Altay, G.; Greenbaum, J.A.; McVicker, G.; et al. Impact of Genetic Polymorphisms on Human Immune Cell Gene Expression. Cell. 2018, 175, 1701–1715.e16. [Google Scholar] [CrossRef]
- Piasecka, B.; Duffy, D.; Urrutia, A.; Quach, H.; Patin, E.; Posseme, C.; Bergstedt, J.; Charbit, B.; Rouilly, V.; MacPherson, C.R.; et al. Distinctive roles of age, sex, and genetics in shaping transcriptional variation of human immune responses to microbial challenges. Proc. Natl. Acad. Sci. USA 2018, 115, E488–E497. [Google Scholar] [CrossRef]
- Xia, S.; Zhang, Y.; Wang, Y.; Wang, H.; Yang, Y.; Gao, G.F.; Tan, W.J.; Wu, G.Z.; Xu, M.; Lou, Z.Y.; et al. Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBIBP-CorV: A randomised, double-blind, placebo-controlled, phase 1/2 trial. Lancet Infect. Dis. 2021, 21, 39–51. [Google Scholar] [CrossRef]
- Zhang, J.; Xing, S.; Liang, D.; Hu, W.; Ke, C.; He, J.; Yuan, R.Y.; Huang, Y.L.; Li, Y.Z.; Liu, X.D.; et al. Differential Antibody Response to Inactivated COVID-19 Vaccines in Healthy Subjects. Front. Cell. Infect. Microbiol. 2021, 11, 791660. [Google Scholar] [CrossRef]
- Walsh, E.E.; Frenck, R.W., Jr.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based COVID-19 Vaccine Candidates. N. Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef]
- Li, X.; Pang, L.; Yin, Y.; Zhang, Y.; Xu, S.; Xu, D.; Shen, T. Patient and Clinical Factors at Admission Affect the Levels of Neutralizing Antibodies Six Months after Recovering from COVID-19. Viruses 2022, 14, 80. [Google Scholar] [CrossRef] [PubMed]
- Tregoning, J.S.; Flight, K.E.; Higham, S.L.; Wang, Z.; Pierce, B.F. Progress of the COVID-19 vaccine effort: Viruses, vaccines and variants versus efficacy, effectiveness and escape. Nat. Rev. Immunol. 2021, 21, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Quan, Y.; Xin, Z.T.; Wrammert, J.; Ma, M.J.; Lv, H.; Wang, T.B.; Yang, H.; Richardus, J.H.; Liu, W. Lack of peripheral memory B cell responses in recovered patients with severe acute respiratory syndrome: A six-year follow-up study. J. Immunol. 2011, 186, 7264–7268. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.T.; Peng, H.; Zhu, Z.L.; Li, G.; Huang, Z.T.; Zhao, Z.X.; Koup, R.A.; Bailer, R.T.; Wu, C.Y. Long-lived effector/central memory T-cell responses to severe acute respiratory syndrome coronavirus (SARS-CoV) S antigen in recovered SARS patients. Clin. Immunol. 2006, 120, 171–178. [Google Scholar] [CrossRef]
- Long, Q.X.; Liu, B.Z.; Deng, H.J.; Wu, G.C.; Deng, K.; Chen, Y.K.; Liao, P.; Qie, J.F.; Lin, Y.; Cai, X.F. Antibody responses to SARS-CoV-2 in patients with COVID-19. Nat. Med. 2020, 26, 845–848. [Google Scholar] [CrossRef]
- Cohen, K.W.; Linderman, S.L.; Moodie, Z.; Czartoski, J.; Lai, L.; Mantus, G.; Norwood, C.; Nyhoff, L.E.; Edara, V.V.; Floyd, K.; et al. Longitudinal analysis shows durable and broad immune memory after SARS-CoV-2 infection with persisting antibody responses and memory B and T cells. Cell Rep. Med. 2021, 2, 100354. [Google Scholar] [CrossRef]
- Rodda, L.B.; Netland, J.; Shehata, L.; Pruner, K.B.; Morawski, P.A.; Thouvenel, C.D.; Takehara, K.K.; Eggenberger, J.; Hemann, E.A.; Waterman, H.R.; et al. Functional SARS-CoV-2-Specific Immune Memory Persists after Mild COVID-19. Cell 2021, 184, 169–183.e17. [Google Scholar] [CrossRef]
- Xiang, T.; Liang, B.; Fang, Y.; Lu, S.; Li, S.; Wang, H.; Li, H.d.; Yang, X.L.; Shen, S.; Zhu, S.; et al. Declining Levels of Neutralizing Antibodies Against SARS-CoV-2 in Convalescent COVID-19 Patients One Year Post Symptom Onset. Front. Immunol. 2021, 12, 708523. [Google Scholar] [CrossRef]
- Beaudoin-Bussières, G.; Laumaea, A.; Anand, S.P.; Prévost, J.; Gasser, R.; Goyette, G.; Medjahed, H.; Perreaut, J.; Tremblay, T.; Lewin, A.; et al. Decline of Humoral Responses against SARS-CoV-2 Spike in Convalescent Individuals. mBio 2020, 11, e02590-20. [Google Scholar] [CrossRef]
- Seow, J.; Graham, C.; Merrick, B.; Acors, S.; Pickering, S.; Steel, K.J.A.; Hemmings, O.; O’Byrne, A.; Kouphou, N.; Galao, R.P.; et al. Longitudinal observation and decline of neutralizing antibody responses in the three months following SARS-CoV-2 infection in humans. Nat. Microbiol. 2020, 5, 1598–1607. [Google Scholar] [CrossRef]
- Thomopoulos, T.P.; Rosati, M.; Terpos, E.; Stellas, D.; Hu, X.; Karaliota, S.; Bouchla, A.; Katagas, L.; Antoniadou, A.; Mentis, A.; et al. Kinetics of Nucleocapsid, Spike and Neutralizing Antibodies, and Viral Load in Patients with Severe COVID-19 Treated with Convalescent Plasma. Viruses 2021, 13, 1844. [Google Scholar] [CrossRef]
- Addetia, A.; Crawford, K.H.D.; Dingens, A.; Zhu, H.; Roychoudhury, P.; Huang, M.L.; Jerome, K.R.; Bloom, J.D.; Greninger, A.L. Neutralizing Antibodies Correlate with Protection from SARS-CoV-2 in Humans during a Fishery Vessel Outbreak with a High Attack Rate. J. Clin. Microbiol. 2020, 58, e02107-20. [Google Scholar] [CrossRef]
- Guo, Y.; Huang, L.; Zhang, G.; Yao, Y.; Zhou, H.; Shen, S.; Shen, B.Q.; Li, B.; Li, X.; Zhang, Q.; et al. A SARS-CoV-2 neutralizing antibody with extensive Spike binding coverage and modified for optimal therapeutic outcomes. Nat. Commun. 2021, 12, 2623. [Google Scholar] [CrossRef] [PubMed]
- China NHCotPsRo. Trial 8th Edition Diagnosis and Treatment Program of Novel Coronavirus Pneumonia (Last Updated: 15 April 2021). Available online: http://www.nhc.gov.cn/xcs/zhengcwj/202104/7de0b3837c8b4606a0594aeb0105232b.shtml (accessed on 15 April 2022).
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L.; Bird, B.H.; Glass, G.E. Sex differences in immune responses and viral shedding following Seoul virus infection in Norway rats. Am. J. Trop. Med. Hyg. 2001, 65, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Park, M.D. Sex differences in immune responses in COVID-19. Nat. Rev. Immunol. 2020, 20, 461. [Google Scholar] [CrossRef]
- Fathi, A.; Addo, M.M.; Dahlke, C. Sex Differences in Immunity: Implications for the Development of Novel Vaccines Against Emerging Pathogens. Front. Immunol. 2020, 11, 601170. [Google Scholar] [CrossRef]
- Selvi, V. Convalescent Plasma: A Challenging Tool to Treat COVID-19 Patients-A Lesson from the Past and New Perspectives. BioMed Res. Int. 2020, 2020, 2606058. [Google Scholar] [CrossRef]
- Rössler, A.; Riepler, L.; Bante, D.; von Laer, D.; Kimpel, J. SARS-CoV-2 Omicron Variant Neutralization in Serum from Vaccinated and Convalescent Persons. N. Engl. J. Med. 2022, 386, 698–700. [Google Scholar] [CrossRef]
- Khoury, J.; Najjar-Debbiny, R.; Hanna, A.; Jabbour, A.; Abu Ahmad, Y.; Saffuri, A.; Abu-Sinni, M.; Shkeiri, R.; Elemy, A.; Hakim, F. COVID-19 vaccine -Long term immune decline and breakthrough infections. Vaccine 2021, 39, 6984–6989. [Google Scholar] [CrossRef]
- Choi, J.Y.; Smith, D.M. SARS-CoV-2 Variants of Concern. Yonsei Med. J. 2021, 62, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Sanyaolu, A.; Okorie, C.; Marinkovic, A.; Haider, N.; Abbasi, A.F.; Jaferi, U.; Prakash, S.; Balendra, V. The emerging SARS-CoV-2 variants of concern. Ther. Adv. Infect. Dis. 2021, 8, 20499361211024372. [Google Scholar] [CrossRef] [PubMed]
- Campbell, F.; Archer, B.; Laurenson-Schafer, H.; Jinnai, Y.; Konings, F.; Batra, N.; Pavlin, B.; Vandemaele, K.; Kerkhove, M.D.V.; Jombart, T.; et al. Increased transmissibility and global spread of SARS-CoV-2 variants of concern as at June 2021. Eurosurveillance 2021, 26, 2100509. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Li, B.; Hao, P.; Song, Y.; Xu, W.; Liu, N.; Lei, C.L.; Ke, C.W. A Case of New Variant COVID-19 First Emerging in South Africa Detected in Airplane Pilot—Guangdong Province, China, January 6, 2021. China CDC Wkly. 2021, 3, 28–29. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Male (n = 56) | Female (n = 58) | Total (n = 114) | p-Value | |
|---|---|---|---|---|
| Age, years | 0.192 | |||
| <18 | 10 | 4 | 14 | |
| 18–50 | 33 | 37 | 70 | |
| ≥50 | 13 | 17 | 30 | |
| Median age (IQR) | 39.00 (27.00–48.75) | 39.00 (29.75–52.25) | 39.00 (27.75–50.25) | |
| Clinical classification | 1.000 | |||
| Asymptomatic case | 9 | 9 | 18 | |
| Mild case | 7 | 9 | 16 | |
| Moderate case | 40 | 40 | 80 | |
| NAbs titer | ||||
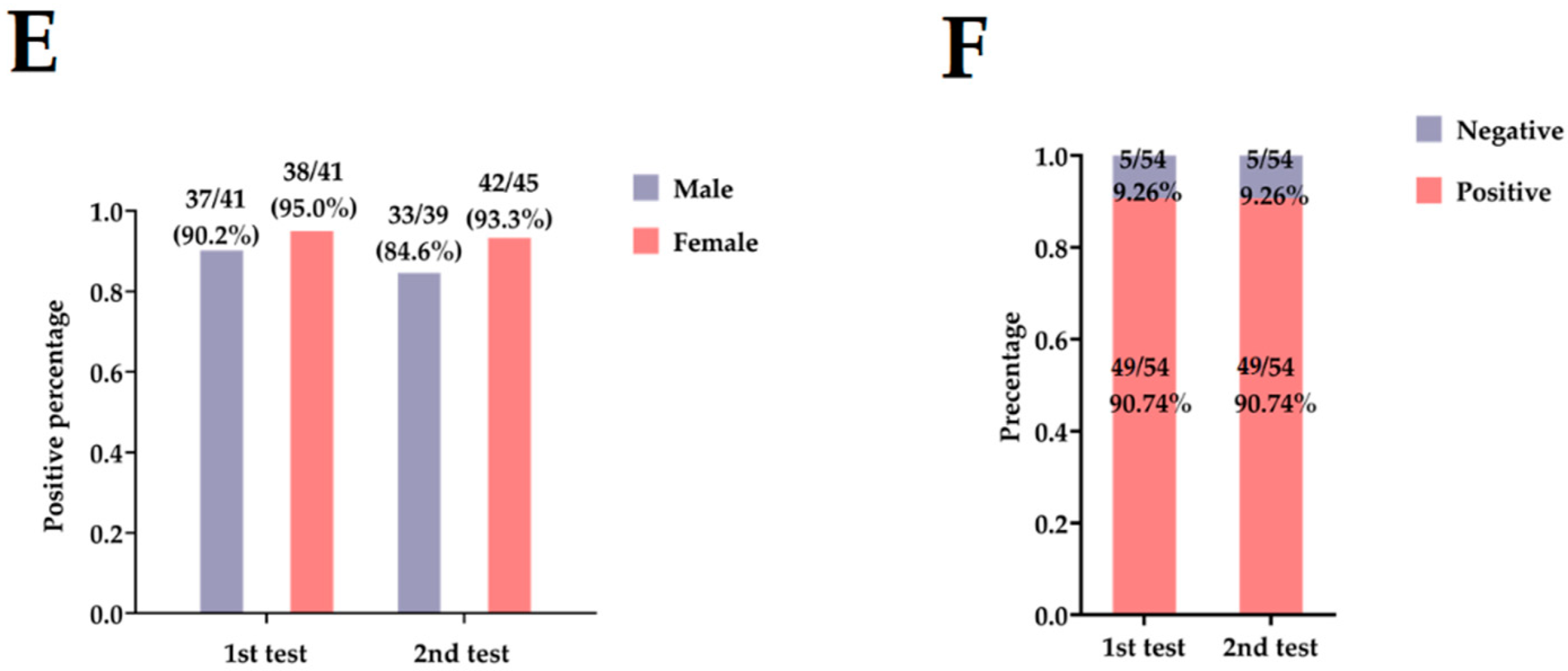
| 1st test (n = 82) | 41 | 41 | 82 | 1.000 |
| Seronegative (<1:4) | 4 | 3 | 7 | |
| Seropositive (≥1:4) | 37 | 38 | 75 | |
| GMT1st test | 1:23.21 | |||
| 2nd test (n = 84) | 39 | 45 | 84 | 0.292 |
| Seronegative (<1:4) | 6 | 3 | 9 | |
| Seropositive (≥1:4) | 33 | 42 | 75 | |
| GMT2nd test | 1:17.09 | |||
| Average duration of disease (days) | 25.06 | 22.87 | 23.90 | 0.258 |
| Downtrend Curve 1 | Uptrend Curve 2 | Unconverted Curve 3 | p-Value | ||
|---|---|---|---|---|---|
| Total | 10 | 27 | 17 | 0.302 | |
| Male | 7 | 12 | 7 | ||
| Female | 3 | 15 | 10 | ||
| Age (years) | 39.00 (30.00–49.00) | 43.00 (31.50–51.00) | 22.50 (16.00–36.25) | 0.026 a* | |
| Clinical classification | Asymptomatic case | 6 | 5 | 1 | 0.003 b* |
| Mild case | 2 | 3 | 5 | ||
| Moderate case | 1 | 18 | 11 | ||
| Duration of disease | 20.69 | 27.88 | 29.89 | 0.015 c* |
| SAlpha (n = 3) | SBeta (n = 3) | SDelta (n = 31) | SOmicron (n = 25) | |
|---|---|---|---|---|
| Median Age, years | 39 | 47 | 44 | 35 |
| Gender | ||||
| Male | 2 | 3 | 12 | 4 |
| Female | 1 | 0 | 19 | 21 |
| Vaccination history | ||||
| Non-vaccinees | 3 | 3 | 19 | 1 |
| 1st dose | 0 | 0 | 7 | 1 |
| 2nd dose | 0 | 0 | 5 | 9 |
| 3rd dose | 0 | 0 | 0 | 14 |
| GMTs | ||||
| VWild | 50.4 | 5.85 | 25.60 | 263.55 |
| VAlpha | 25.20 | 1.00 | 16.62 | 256.34 |
| VBeta | 31.75 | 7.94 | 12.06 | 164.50 |
| VGamma | 80.00 | 40.00 | 87.48 | 572.82 |
| VDelta | 31.75 | 3.42 | 101.57 | 223.16 |
| VOmicron | 2.15 | 1.71 | 3.19 | 102.67 |
| Vaccination History | GMTs of SDelta (n = 31) | Vaccination History | GMTs of SOmicron (n = 25) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| VWild | VAlpha | VBeta | VGamma | VDelta | VOmicron | VWild | VAlpha | VBeta | VGamma | VDelta | VOmicron | ||
| Non-vaccinees (n = 19) | 18.84 | 24.32 | 19.54 | 115.22 | 160 | 4.78 | Non-vaccinees (n = 1) | 160 | 160 | 20 | 160 | 160 | 40 |
| 1st dose (n = 7) | 63.57 | 5.72 | 3.38 | 24.38 | 23.61 | 1.26 | 1st dose (n = 1) | 10 | 160 | 160 | 320 | 80 | 40 |
| 2nd dose (n = 5) | 22.97 | 17.41 | 11.49 | 183.79 | 139.29 | 2.51 | 2nd dose (n = 9) | 296.28 | 320 | 217.73 | 507.97 | 217.73 | 93.32 |
| 3rd dose (n = 0) | 0 | 0 | 0 | 0 | 0 | 0 | 3rd dose (n = 14) | 320 | 237.76 | 160 | 706.62 | 249.83 | 124.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, D.; Zhang, G.; Huang, M.; Wang, L.; Hong, W.; Li, A.; Liang, Y.; Wang, T.; Lu, J.; Ou, M.; et al. Progress of the COVID-19: Persistence, Effectiveness, and Immune Escape of the Neutralizing Antibody in Convalescent Serum. Pathogens 2022, 11, 1531. https://doi.org/10.3390/pathogens11121531
Liang D, Zhang G, Huang M, Wang L, Hong W, Li A, Liang Y, Wang T, Lu J, Ou M, et al. Progress of the COVID-19: Persistence, Effectiveness, and Immune Escape of the Neutralizing Antibody in Convalescent Serum. Pathogens. 2022; 11(12):1531. https://doi.org/10.3390/pathogens11121531
Chicago/Turabian StyleLiang, Dan, Guanting Zhang, Mingxing Huang, Li Wang, Wenshan Hong, An’an Li, Yufeng Liang, Tao Wang, Jiahui Lu, Mengdang Ou, and et al. 2022. "Progress of the COVID-19: Persistence, Effectiveness, and Immune Escape of the Neutralizing Antibody in Convalescent Serum" Pathogens 11, no. 12: 1531. https://doi.org/10.3390/pathogens11121531