
Virulence Profile, Antibiotic Resistance, and Phylogenetic Relationships among Escherichia coli Strains Isolated from the Feces and Urine of Hospitalized Patients
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients, Sample Collection, and Strain Identification
2.2. Clonal Analyses of the Isolates
2.3. Whole-Genome Sequencing and Genomic Analyses
2.4. Evaluation of Antimicrobial Susceptibility
2.5. Ethical Approval
3. Results
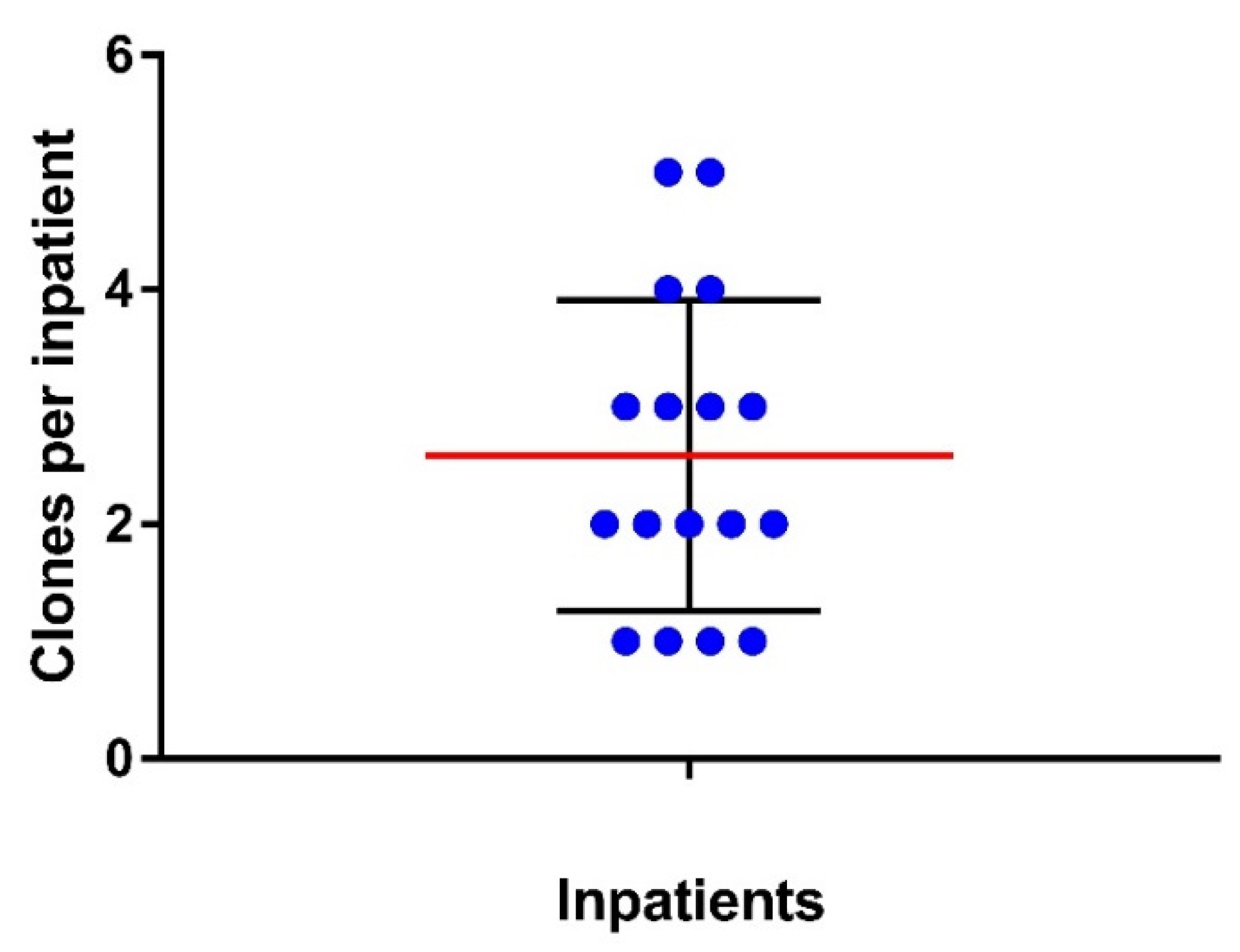
3.1. Clonal Diversity of E. coli Strains from the Gut of Inpatients
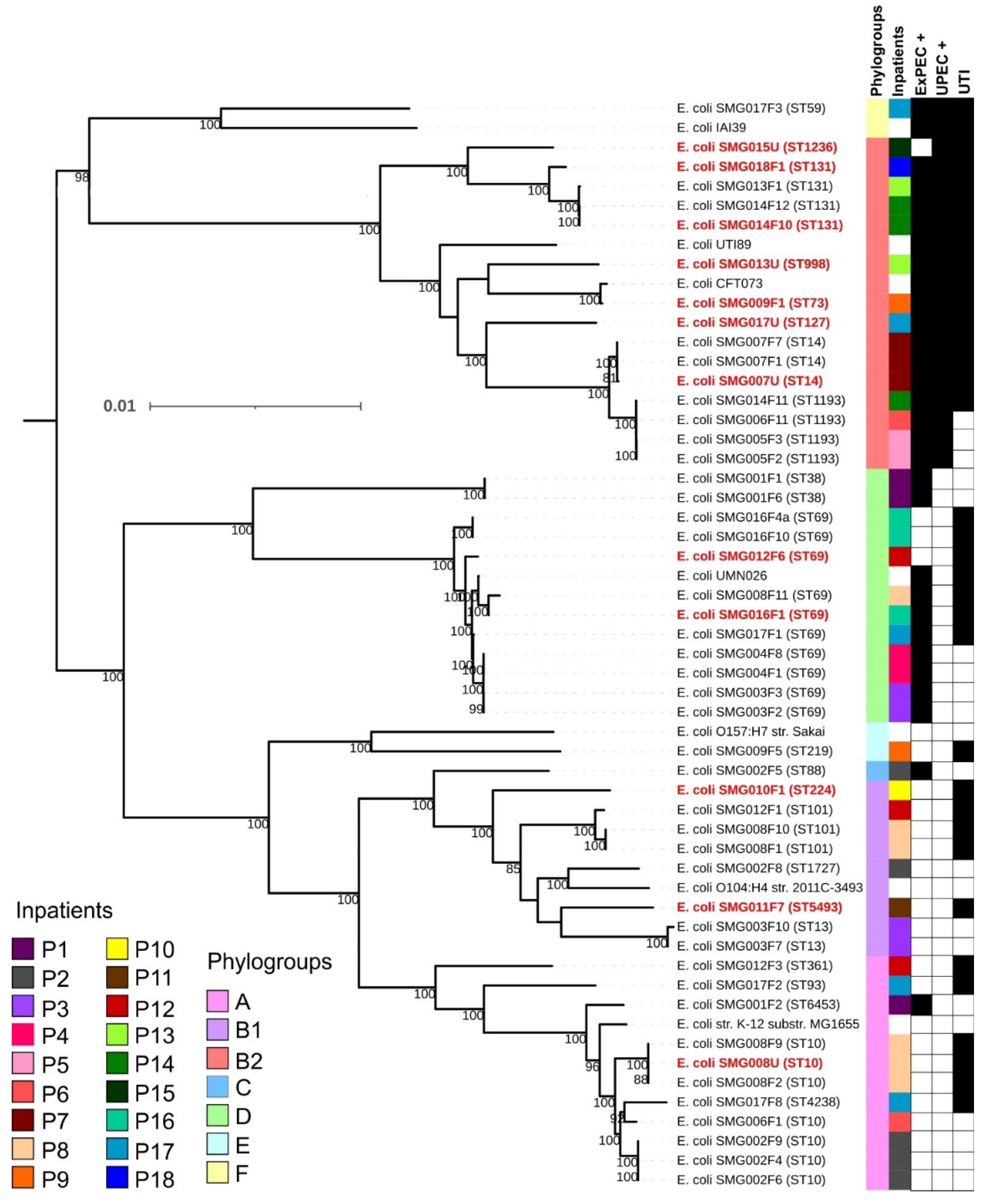
3.2. Phylogenetic Classification and Multilocus Sequence Types among Sequenced E. coli Clones
3.3. Virulence Profile, Intrinsic Virulence, and Uropathogenic Potential of E. coli Strains from the Gut and Urine
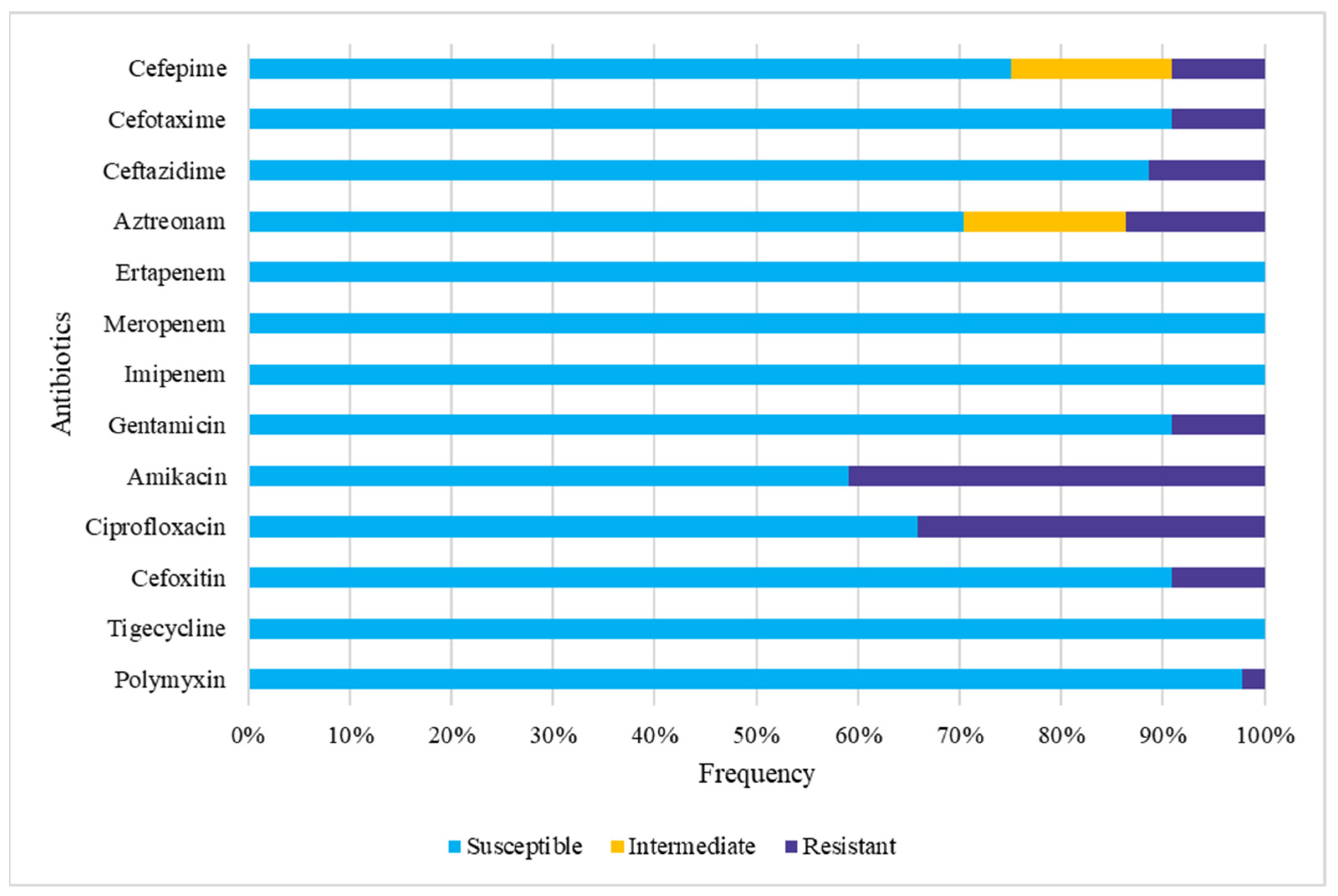
3.4. Gut Colonization by Multidrug-Resistant (MDR) E. coli
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Russo, T.A.; Johnson, J.R. Proposal for a New Inclusive Designation for Extraintestinal Pathogenic Isolates of Escherichia coli: ExPEC. J. Infect. Dis. 2000, 181, 1753–1754. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Russo, T.A. Extraintestinal pathogenic Escherichia coli: “The other bad E. coli”. J. Lab. Clin. Med. 2002, 139, 155–162. [Google Scholar] [CrossRef]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Dale, A.P.; Woodford, N. Extra-intestinal pathogenic Escherichia coli (ExPEC): Disease, carriage and clones. J. Infect. 2015, 71, 615–626. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Russo, T.A. Molecular Epidemiology of Extraintestinal Pathogenic Escherichia coli. EcoSal Plus 2018, 8, ecosalplus. [Google Scholar] [CrossRef]
- Johnson, J.R.; Murray, A.C.; Gajewski, A.; Sullivan, M.; Snippes, P.; Kuskowski, M.A.; Smith, K.E. Isolation and molecular characterization of nalidixic acid-resistant extraintestinal pathogenic Escherichia coli from retail chicken products. Antimicrob. Agents Chemother. 2003, 47, 2161–2168. [Google Scholar] [CrossRef] [Green Version]
- Spurbeck, R.R.; Dinh, P.C.; Walk, S.T.; Stapleton, A.E.; Hooton, T.M.; Nolan, L.K.; Kim, K.S.; Johnson, J.R.; Mobley, H.L.T. Escherichia coli Isolates That Carry vat, fyuA, chuA, and yfcV Efficiently Colonize the Urinary Tract. Infect. Immun. 2012, 80, 4115–4122. [Google Scholar] [CrossRef] [Green Version]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef]
- Abou Heidar, N.; Degheili, J.; Yacoubian, A.; Khauli, R. Management of urinary tract infection in women: A practical approach for everyday practice. Urol. Ann. 2019, 11, 339. [Google Scholar] [CrossRef]
- Forde, B.M.; Roberts, L.W.; Phan, M.D.; Peters, K.M.; Fleming, B.A.; Russell, C.W.; Lenherr, S.M.; Myers, J.B.; Barker, A.P.; Fisher, M.A.; et al. Population dynamics of an Escherichia coli ST131 lineage during recurrent urinary tract infection. Nat. Commun. 2019, 10, 3643. [Google Scholar] [CrossRef] [PubMed]
- Terlizzi, M.E.; Gribaudo, G.; Maffei, M.E. UroPathogenic Escherichia coli (UPEC) infections: Virulence factors, bladder responses, antibiotic, and non-antibiotic antimicrobial strategies. Front. Microbiol. 2017, 8, 1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, M.V.M. Phylogenetic Origin and Virulence Potential of Escherichia coli Belonging to the Intestinal Flora of Healthy Humans. Master’s Thesis, Universidade Federal de São Paulo, São Paulo, Brazil, 2011. [Google Scholar]
- Xie, J.; Foxman, B.; Zhang, L.; Marrs, C.F. Molecular Epidemiologic Identification of Escherichia coli Genes That Are Potentially Involved in Movement of the Organism from the Intestinal Tract to the Vagina and Bladder. J. Clin. Microbiol. 2006, 44, 2434–2441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Clabots, C.; Kuskowski, M.A. Multiple-Host Sharing, Long-Term Persistence, and Virulence of Escherichia coli Clones from Human and Animal Household Members. J. Clin. Microbiol. 2008, 46, 4078–4082. [Google Scholar] [CrossRef] [Green Version]
- Nowrouzian, F.L.; Wold, A.E.; Adlerberth, I. Escherichia coli Strains Belonging to Phylogenetic Group B2 Have Superior Capacity to Persist in the Intestinal Microflora of Infants. J. Infect. Dis. 2005, 191, 1078–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, C.W.; Richards, A.C.; Chang, A.S.; Mulvey, M.A. The Rhomboid Protease GlpG Promotes the Persistence of Extraintestinal Pathogenic Escherichia coli within the Gut. Infect. Immun. 2017, 85, e00866-16. [Google Scholar] [CrossRef] [Green Version]
- Toledo, M.; Fontes, C.; Trabulsi, L. EPM—Modificacao do meio de Rugai e Araujo para a realizacao simultanea dos testes de producao de gas a partir da glicose, H2S, urease e triptofano desaminase. Rev. Microbiol. 1982, 13, 309–315. [Google Scholar]
- Toledo, M.; Fontes, C.; Trabulsi, L. MILi—Um meio para a realizacao dos testes de motilidade, indol e lisina descarboxilase. Rev. Microbiol. 1982, 13, 230–235. [Google Scholar]
- Farmer, J.J.; Davis, B.R.; Hickman-Brenner, F.W.; McWhorter, A.; Huntley-Carter, G.P.; Asbury, M.A.; Riddle, C.; Wathen-Grady, H.G.; Elias, C.; Fanning, G.R. Biochemical identification of new species and biogroups of Enterobacteriaceae isolated from clinical specimens. J. Clin. Microbiol. 1985, 21, 46–76. [Google Scholar] [CrossRef] [Green Version]
- Starr, M.P. Edwards and Ewing’s Identification of Enterobacteriaceae. Int. J. Syst. Bacteriol. 1986, 36, 581–582. [Google Scholar] [CrossRef]
- Nielsen, K.L.; Dynesen, P.; Larsen, P.; Frimodt-Møller, N. Faecal Escherichia coli from patients with E. coli urinary tract infection and healthy controls who have never had a urinary tract infection. J. Med. Microbiol. 2014, 63, 582–589. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, K.L.; Godfrey, P.A.; Stegger, M.; Andersen, P.S.; Feldgarden, M.; Frimodt-Møller, N. Selection of unique Escherichia coli clones by random amplified polymorphic DNA (RAPD): Evaluation by whole genome sequencing. J. Microbiol. Methods 2014, 103, 101–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.C.M.; Fuga, B.; Esposito, F.; Cardoso, B.; Santos, F.F.; Valiatti, T.B.; Santos-Neto, J.F.; Gales, A.C.; Lincopan, N.; Silva, R.M.; et al. Unveiling the Virulent Genotype and Unusual Biochemical Behavior of Escherichia coli ST59. Appl. Environ. Microbiol. 2021, 87, e0074321. [Google Scholar] [CrossRef]
- Malberg Tetzschner, A.M.; Johnson, J.R.; Johnston, B.D.; Lund, O.; Scheutz, F. In Silico Genotyping of Escherichia coli Isolates for Extraintestinal Virulence Genes by Use of Whole-Genome Sequencing Data. J. Clin. Microbiol. 2020, 58, e01269-20. [Google Scholar] [CrossRef]
- Joensen, K.G.; Tetzschner, A.M.M.; Iguchi, A.; Aarestrup, F.M.; Scheutz, F. Rapid and easy in silico serotyping of Escherichia coli isolates by use of whole-genome sequencing data. J. Clin. Microbiol. 2015, 53, 2410–2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In Silico Detection and Typing of Plasmids using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- Roer, L.; Johannesen, T.B.; Hansen, F.; Stegger, M.; Tchesnokova, V.; Sokurenko, E.; Garibay, N.; Allesøe, R.; Thomsen, M.C.F.; Lund, O.; et al. CHTyper, a web tool for subtyping of extraintestinal pathogenic Escherichia coli based on the fumC and fimH alleles. J. Clin. Microbiol. 2018, 56, 63–81. [Google Scholar] [CrossRef] [Green Version]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.J.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Ponten, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.H.; McDermott, P.F.; et al. Validating the AMRFinder Tool and Resistance Gene Database by Using Antimicrobial Resistance Genotype-Phenotype Correlations in a Collection of Isolates. Antimicrob. Agents Chemother. 2019, 63, e00483-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beghain, J.; Bridier-Nahmias, A.; Le Nagard, H.; Denamur, E.; Clermont, O. ClermonTyping: An easy-to-use and accurate in silico method for Escherichia genus strain phylotyping. Microb. Genom. 2018, 4, e000192. [Google Scholar] [CrossRef]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Gordon, D.M.; Bauer, S.; Johnson, J.R. The genetic structure of Escherichia coli populations in primary and secondary habitats. Microbiology 2002, 148, 1513–1522. [Google Scholar] [CrossRef] [Green Version]
- Perez, L.R.R.; Carniel, E.; Narvaez, G.A.; Dias, C.G. Evaluation of a polymyxin drop test for polymyxin resistance detection among non-fermentative gram-negative rods and enterobacterales resistant to carbapenems. Apmis 2021, 129, 138–142. [Google Scholar] [CrossRef]
- Ewers, C.; de Jong, A.; Prenger-Berninghoff, E.; El Garch, F.; Leidner, U.; Tiwari, S.K.; Semmler, T. Genomic Diversity and Virulence Potential of ESBL- and AmpC-β-Lactamase-Producing Escherichia coli Strains From Healthy Food Animals Across Europe. Front. Microbiol. 2021, 12, 635. [Google Scholar] [CrossRef]
- EUCAST. European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters Version 11.0; EUCAST: Växjö, Sweden, 2021; Available online: http://www.eucast.org (accessed on 10 October 2021).
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Leimbach, A.; Hacker, J.; Dobrindt, U. E. coli as an all-rounder: The thin line between commensalism and pathogenicity. Curr. Top. Microbiol. Immunol. 2013, 358, 3–32. [Google Scholar] [CrossRef] [PubMed]
- Madigan, T.; Johnson, J.R.; Clabots, C.; Johnston, B.D.; Porter, S.B.; Slater, B.S.; Banerjee, R. Extensive Household Outbreak of Urinary Tract Infection and Intestinal Colonization due to Extended-Spectrum β-Lactamase–Producing Escherichia coli Sequence Type 131. Clin. Infect. Dis. 2015, 61, e5–e12. [Google Scholar] [CrossRef] [PubMed]
- Mosavie, M.; Blandy, O.; Jauneikaite, E.; Caldas, I.; Ellington, M.J.; Woodford, N.; Sriskandan, S. Sampling and diversity of Escherichia coli from the enteric microbiota in patients with Escherichia coli bacteraemia. BMC Res. Notes 2019, 12, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duriez, P.; Clermont, O.; Picard, B.; Denamur, E.; Bonacorsi, S.; Bingen, E.; Chaventré, A.; Elion, J. Commensal Escherichia coli isolates are phylogenetically distributed among geographically distinct human populations. Microbiology 2001, 147, 1671–1676. [Google Scholar] [CrossRef] [Green Version]
- Escobar-Páramo, P.; Grenet, K.; Le Menac’h, A.; Rode, L.; Salgado, E.; Amorin, C.; Gouriou, S.; Picard, B.; Rahimy, M.C.; Andremont, A.; et al. Large-scale population structure of human commensal Escherichia coli isolates. Appl. Environ. Microbiol. 2004, 70, 5698–5700. [Google Scholar] [CrossRef] [Green Version]
- Gordon, D.M.; Stern, S.E.; Collignon, P.J. Influence of the age and sex of human hosts on the distribution of Escherichia coli ECOR groups and virulence traits. Microbiology 2005, 151, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.; Hu, Y.; Sun, Q.; Hu, F.; Zhou, H.; Shu, L.; Ma, T.; Shen, Y.; Wang, Y.; Li, J.; et al. Emerging Carriage of NDM-5 and MCR-1 in Escherichia coli From Healthy People in Multiple Regions in China: A Cross Sectional Observational Study. EClinicalMedicine 2018, 6, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, R.; Friedman, C.R.; Rubin, J.; Suh, J.; Thys, E.; McDermott, P.; Hung-Fan, M.; Riley, L.W. A Population-Based Surveillance Study of Shared Genotypes of Escherichia coli Isolates from Retail Meat and Suspected Cases of Urinary Tract Infections. mSphere 2018, 3, e00179-18. [Google Scholar] [CrossRef] [Green Version]
- De Stoppe, N.C.; Silva, J.S.; Carlos, C.; Sato, M.I.Z.; Saraiva, A.M.; Ottoboni, L.M.M.; Torres, T.T. Worldwide Phylogenetic Group Patterns of Escherichia coli from Commensal Human and Wastewater Treatment Plant Isolates. Front. Microbiol. 2017, 8, 2512. [Google Scholar] [CrossRef] [Green Version]
- Olesen, B.; Kolmos, H.J.; Ørskov, F.; Ørskov, I. Cluster of Multiresistant Escherichia coli O78:H10 in Greater Copenhagen. Scand. J. Infect. Dis. 1994, 26, 406–410. [Google Scholar] [CrossRef]
- Gomig, F.; Galvão, C.W.; de Freitas, D.L.; Labas, L.; Etto, R.M.; Esmerino, L.A.; de Lima, M.A.; Appel, M.H.; Zanata, S.M.; Steffens, M.B.R.; et al. Quinolone resistance and ornithine decarboxylation activity in lactose-negative Escherichia coli. Braz. J. Microbiol. 2015, 46, 753–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczmarek, A.; Skowron, K.; Budzyńska, A.; Grudlewska, K.; Gospodarek-Komkowska, E. Virulence genes and antimicrobial susceptibility of lactose-negative and lactose-positive strains of Escherichia coli isolated from pregnant women and neonates. Folia Microbiol. 2017, 62, 363–371. [Google Scholar] [CrossRef]
- Johnson, J.R.; Johnston, B.D.; Porter, S.B.; Clabots, C.; Bender, T.L.; Thuras, P.; Trott, D.J.; Cobbold, R.; Mollinger, J.; Ferrieri, P.; et al. Rapid Emergence, Subsidence, and Molecular Detection of Escherichia coli Sequence Type 1193-fimH64, a New Disseminated Multidrug-Resistant Commensal and Extraintestinal Pathogen. J. Clin. Microbiol. 2019, 57, e01664-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanella, A.; Alborali, G.L.; Bardotti, M.; Candotti, P.; Guadagnini, P.F.; Martino, P.A.; Stonfer, M. Severe Escherichia coli O111 septicaemia and polyserositis in hens at the start of lay. Avian Pathol. 2000, 29, 311–317. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Johnston, B.D.; Gordon, D.M. Rapid and specific detection of the Escherichia coli sequence type 648 complex within phylogroup F. J. Clin. Microbiol. 2017, 55, 1116–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Östblom, A.; Adlerberth, I.; Wold, A.E.; Nowrouzian, F.L. Pathogenicity island markers, virulence determinants malX and usp, and the capacity of Escherichia coli to persist in infants’ commensal microbiotas. Appl. Environ. Microbiol. 2011, 77, 2303–2308. [Google Scholar] [CrossRef] [Green Version]
- Richter, T.K.S.; Hazen, T.H.; Lam, D.; Coles, C.L.; Seidman, J.C.; You, Y.; Silbergeld, E.K.; Fraser, C.M.; Rasko, D.A. Temporal Variability of Escherichia coli Diversity in the Gastrointestinal Tracts of Tanzanian Children with and without Exposure to Antibiotics. mSphere 2018, 3, e00558-18. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Saw, W.-Y.; Tan, L.W.L.; Moong, D.K.N.; Nagarajan, N.; Teo, Y.Y.; Seedorf, H. Extended-Spectrum β-Lactamase-Producing and mcr-1-Positive Escherichia coli from the Gut Microbiota of Healthy Singaporeans. Appl. Environ. Microbiol. 2021, 87, e0048821. [Google Scholar] [CrossRef]
- Carlos, C.; Pires, M.M.; Stoppe, N.C.; Hachich, E.M.; Sato, M.I.Z.; Gomes, T.A.T.; Amaral, L.A.; Ottoboni, L.M.M. Escherichia coli phylogenetic group determination and its application in the identification of the major animal source of fecal contamination. BMC Microbiol. 2010, 10, 161. [Google Scholar] [CrossRef] [Green Version]
- Hannah, E.L.; Johnson, J.R.; Angulo, F.; Haddadin, B.; Williamson, J.; Samore, M.H. Molecular analysis of antimicrobial-susceptible and -resistant Escherichia coli from retail meats and human stool and clinical specimens in a rural community setting. Foodborne Pathog. Dis. 2009, 6, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Unno, T.; Han, D.; Jang, J.; Lee, S.-N.; Ko, G.; Choi, H.Y.; Kim, J.H.; Sadowsky, M.J.; Hur, H.-G. Absence of Escherichia coli Phylogenetic Group B2 Strains in Humans and Domesticated Animals from Jeonnam Province, Republic of Korea. Appl. Environ. Microbiol. 2009, 75, 5659–5666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollmerhausen, T.L.; Ramos, N.L.; Gündoğdu, A.; Robinson, W.; Brauner, A.; Katouli, M. Population structure and uropathogenic virulence-associated genes of faecal Escherichia coli from healthy young and elderly adults. J. Med. Microbiol. 2011, 60, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Grude, N.; Potaturkina-Nesterova, N.I.; Jenkins, A.; Strand, L.; Nowrouzian, F.L.; Nyhus, J.; Kristiansen, B.E. A comparison of phylogenetic group, virulence factors and antibiotic resistance in Russian and Norwegian isolates of Escherichia coli from urinary tract infection. Clin. Microbiol. Infect. 2007, 13, 208–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and Simple Determination of the Escherichia coli Phylogenetic Group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [Green Version]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef]
- Massot, M.; Daubié, A.-S.; Clermont, O.; Jauréguy, F.; Couffignal, C.; Dahbi, G.; Mora, A.; Blanco, J.; Branger, C.; Mentré, F.; et al. Phylogenetic, virulence and antibiotic resistance characteristics of commensal strain populations of Escherichia coli from community subjects in the Paris area in 2010 and evolution over 30 years. Microbiology 2016, 162, 642–650. [Google Scholar] [CrossRef]
- Smati, M.; Clermont, O.; Le Gal, F.; Schichmanoff, O.; Jauréguy, F.; Eddi, A.; Denamur, E.; Picard, B. Real-time PCR for quantitative analysis of human commensal Escherichia coli populations reveals a high frequency of subdominant phylogroups. Appl. Environ. Microbiol. 2013, 79, 5005–5012. [Google Scholar] [CrossRef] [Green Version]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global Extraintestinal Pathogenic Escherichia coli (ExPEC) Lineages. Clin. Microbiol. Rev. 2019, 32, e00135-18. [Google Scholar] [CrossRef]
- Manges, A.R.; Johnson, J.R.; Foxman, B.; O’Bryan, T.T.; Fullerton, K.E.; Riley, L.W. Widespread Distribution of Urinary Tract Infections Caused by a Multidrug-Resistant Escherichia coli Clonal Group. N. Engl. J. Med. 2001, 345, 1007–1013. [Google Scholar] [CrossRef]
- Hertz, F.B.; Nielsen, J.B.; Schønning, K.; Littauer, P.; Knudsen, J.D.; Løbner-Olesen, A.; Frimodt-Møller, N. Population structure of Drug-Susceptible, -Resistant and ESBL-producing Escherichia coli from Community-Acquired Urinary Tract Infections. BMC Microbiol. 2016, 16, 63. [Google Scholar] [CrossRef] [Green Version]
- Kallonen, T.; Brodrick, H.J.; Harris, S.R.; Corander, J.; Brown, N.M.; Martin, V.; Peacock, S.J.; Parkhill, J. Systematic longitudinal survey of invasive Escherichia coli in England demonstrates a stable population structure only transiently disturbed by the emergence of ST131. Genome Res. 2017, 27, 1437–1449. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, R.; Rubin, J.; Thys, E.; Friedman, C.R.; Riley, L.W. Persistent Pandemic Lineages of Uropathogenic Escherichia coli in a College Community from 1999 to 2017. J. Clin. Microbiol. 2018, 56, e01834-17. [Google Scholar] [CrossRef] [PubMed]
- Dias, R.C.; Marangoni, D.V.; Riley, L.W.; Moreira, B.M. Identification of uropathogenic Escherichia coli clonal group A (CgA) in hospitalised patients. Mem. Inst. Oswaldo Cruz 2009, 104, 787–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, R.C.S.; Marangoni, D.V.; Smith, S.P.; Alves, E.M.; Pellegrino, F.L.P.C.; Riley, L.W.; Moreira, B.M. Clonal composition of Escherichia coli causing community-acquired urinary tract infections in the state of rio de janeiro, Brazil. Microb. Drug Resist. 2009, 15, 303–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, A.C.M.; Zidko, A.C.M.; Pignatari, A.C.; Silva, R.M. Assessing the diversity of the virulence potential of Escherichia coli isolated from bacteremia in São Paulo, Brazil. Braz. J. Med. Biol. Res. 2013, 46, 968–973. [Google Scholar] [CrossRef] [Green Version]
- Lara, F.B.M.; Nery, D.R.; de Oliveira, P.M.; Araujo, M.L.; Carvalho, F.R.Q.; Messias-Silva, L.C.F.; Ferreira, L.B.; Faria-Junior, C.; Pereira, A.L. Virulence Markers and Phylogenetic Analysis of Escherichia coli Strains with Hybrid EAEC/UPEC Genotypes Recovered from Sporadic Cases of Extraintestinal Infections. Front. Microbiol. 2017, 8, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, A.C.C.; Andrade, N.L.; Ferdous, M.; Chlebowicz, M.A.; Santos, C.C.; Correal, J.C.D.; Lo Ten Foe, J.R.; Rosa, A.C.P.; Damasco, P.V.; Friedrich, A.W.; et al. Comprehensive Molecular Characterization of Escherichia coli Isolates from Urine Samples of Hospitalized Patients in Rio de Janeiro, Brazil. Front. Microbiol. 2018, 9, 243. [Google Scholar] [CrossRef]
- Paiva, Y.; Nagano, D.S.; Cotia, A.L.F.; Guimarães, T.; Martins, R.C.R.; Perdigão Neto, L.V.; Côrtes, M.F.; Marchi, A.P.; Corscadden, L.; Machado, A.S.; et al. Colistin-resistant Escherichia coli belonging to different sequence types: Genetic characterization of isolates responsible for colonization, community- and healthcare-acquired infections. Rev. Inst. Med. Trop. Sao Paulo 2021, 63, e38. [Google Scholar] [CrossRef]
- Alghoribi, M.F.; Gibreel, T.M.; Farnham, G.; Al Johani, S.M.; Balkhy, H.H.; Upton, M. Antibiotic-resistant ST38, ST131 and ST405 strains are the leading uropathogenic Escherichia coli clones in Riyadh, Saudi Arabia. J. Antimicrob. Chemother. 2015, 70, 2757–2762. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Tian, G.-B.; Zhang, R.; Shen, Y.; Tyrrell, J.M.; Huang, X.; Zhou, H.; Lei, L.; Li, H.-Y.; Doi, Y.; et al. Prevalence, risk factors, outcomes, and molecular epidemiology of mcr-1-positive Enterobacteriaceae in patients and healthy adults from China: An epidemiological and clinical study. Lancet Infect. Dis. 2017, 17, 390–399. [Google Scholar] [CrossRef] [Green Version]
- Bozcal, E.; Eldem, V.; Aydemir, S.; Skurnik, M. The relationship between phylogenetic classification, virulence and antibiotic resistance of extraintestinal pathogenic Escherichia coli in İzmir province, Turkey. PeerJ 2018, 6, e5470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mbelle, N.M.; Feldman, C.; Osei Sekyere, J.; Maningi, N.E.; Modipane, L.; Essack, S.Y. The Resistome, Mobilome, Virulome and Phylogenomics of Multidrug-Resistant Escherichia coli Clinical Isolates from Pretoria, South Africa. Sci. Rep. 2019, 9, 16457. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.; Menard, M.; Lauderdale, T.-L.; Lauderdale, T.-L.; Kosmidis, C.; Gordon, D.; Collignon, P.; Maslow, J.; Andrasević, A.; Kuskowski, M. Global Distribution and Epidemiologic Associations of Escherichia coli Clonal Group A, 1998–2007. Emerg. Infect. Dis. 2011, 17, 2001–2009. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Zhang, Z.; Shen, H.; Ning, M.; Chen, J.; Wei, H.; Zhang, K. Genotypic characteristics of multidrug-resistant Escherichia coli isolates associated with urinary tract infections. APMIS 2014, 122, 1088–1095. [Google Scholar] [CrossRef]
- Silva, M.M.; Sellera, F.P.; Fernandes, M.R.; Moura, Q.; Garino, F.; Azevedo, S.S.; Lincopan, N. Genomic features of a highly virulent, ceftiofur-resistant, CTX-M-8-producing Escherichia coli ST224 causing fatal infection in a domestic cat. J. Glob. Antimicrob. Resist. 2018, 15, 252–253. [Google Scholar] [CrossRef]
- Schmitt, K.; Kuster, S.P.; Zurfluh, K.; Jud, R.S.; Sykes, J.E.; Stephan, R.; Willi, B. Transmission Chains of Extended-Spectrum Beta-Lactamase-Producing Enterobacteriaceae at the Companion Animal Veterinary Clinic–Household Interface. Antibiotics 2021, 10, 171. [Google Scholar] [CrossRef]
- Fuga, B.; Sellera, F.P.; Cerdeira, L.; Esposito, F.; Cardoso, B.; Fontana, H.; Moura, Q.; Cardenas-Arias, A.; Sano, E.; Ribas, R.M.; et al. WHO Critical Priority Escherichia coli as One Health Challenge for a Post-Pandemic Scenario: Genomic Surveillance and Analysis of Current Trends in Brazil. Microbiol. Spectr. 2022, 10, e0125621. [Google Scholar] [CrossRef]
- Roer, L.; Overballe-Petersen, S.; Hansen, F.; Schønning, K.; Wang, M.; Røder, B.L.; Hansen, D.S.; Justesen, U.S.; Andersen, L.P.; Fulgsang-Damgaard, D.; et al. Escherichia coli Sequence Type 410 Is Causing New International High-Risk Clones. mSphere 2018, 3, e00337-18. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.C.M.; Silva, R.M.; Valiatti, T.B.; Santos, F.F.; Santos-Neto, J.F.; Cayô, R.; Streling, A.P.; Nodari, C.S.; Gales, A.C.; Nishiyama, M.Y., Jr.; et al. Virulence Potential of a Multidrug-Resistant Escherichia coli Strain Belonging to the Emerging Clonal Group ST101-B1 Isolated from Bloodstream Infection. Microorganisms 2020, 8, 827. [Google Scholar] [CrossRef]
- Graham, J.P.; Amato, H.K.; Mendizabal-Cabrera, R.; Alvarez, D.; Ramay, B.M. Waterborne Urinary Tract Infections: Have We Overlooked an Important Source of Exposure? Am. J. Trop. Med. Hyg. 2021, 105, 12–17. [Google Scholar] [CrossRef]
- Manges, A.R.; Johnson, J.R. Reservoirs of Extraintestinal Pathogenic Escherichia coli. Microbiol. Spectr. 2015, 3, UTI-0006-2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludden, C.; Coll, F.; Gouliouris, T.; Restif, O.; Blane, B.; Blackwell, G.A.; Kumar, N.; Naydenova, P.; Crawley, C.; Brown, N.M.; et al. Defining nosocomial transmission of Escherichia coli and antimicrobial resistance genes: A genomic surveillance study. Lancet Microbe 2021, 2, e472–e480. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.; Hang, X.; Li, X.; Ye, N.; Tang, W.; Zhang, Y.; Yang, X.; Yang, M.; Wang, Y.; Liu, Z.; et al. Bloodstream Infections in Patients with Rectal Colonization by Carbapenem-Resistant Enterobacteriaceae: A Prospective Cohort Study. Infect. Drug Resist. 2022, 15, 6051. [Google Scholar] [CrossRef] [PubMed]
- Freire, S.; Grilo, T.; Teixeira, M.L.; Fernandes, E.; Poirel, L.; Aires-de-Sousa, M. Screening and Characterization of Multidrug-Resistant Enterobacterales among Hospitalized Patients in the African Archipelago of Cape Verde. Microorganisms 2022, 10, 1426. [Google Scholar] [CrossRef] [PubMed]
- Spoto, S.; Daniel Markley, J.; Valeriani, E.; Abbate, A.; Argemi, J.; Markley, R.; Fogolari, M.; Locorriere, L.; Anguissola, G.B.; Battifoglia, G.; et al. Active Surveillance Cultures and Procalcitonin in Combination with Clinical Data to Guide Empirical Antimicrobial Therapy in Hospitalized Medical Patients with Sepsis. Front. Microbiol. 2022, 13, 797932. [Google Scholar] [CrossRef]
- Farrell, J.J.; Hicks, J.L.; Wallace, S.E.; Seftel, A.D. Impact of preoperative screening for rectal colonization with fluoroquinolone-resistant enteric bacteria on the incidence of sepsis following transrectal ultrasound guided prostate biopsy. Res. Rep. Urol. 2017, 9, 37–41. [Google Scholar] [CrossRef] [Green Version]
- Ceccarelli, G.; Alessandri, F.; Moretti, S.; Borsetti, A.; Maggiorella, M.T.; Fabris, S.; Russo, A.; Ruberto, F.; De Meo, D.; Ciccozzi, M.; et al. Clinical Impact of Colonization with Carbapenem-Resistant Gram-Negative Bacteria in Critically Ill Patients Admitted for Severe Trauma. Pathogens 2022, 11, 1295. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | N° (%) | |
|---|---|---|
| Gender | ||
| Female | 15 (83.3) | |
| Male | 3 (16.7) | |
| Antibiotic use | 12 (66.7) | |
| UTI agent | ||
| E. coli | 12 (66.7) | |
| Another pathogen a | 1 (5.6) | |
| Strains | SAM | ATM | CRO | CAZ | FEP | ETP | IPM | MEM | TGC | CIP | LVX | AMK | GEN | CST | PMB |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SMG008F2 | >128/4 | 64 | 256 | 128 | 64 | <0.06 | <0.06 | <0.06 | 16 | 64 | 32 | 128 | 16 | <0.25 | <0.25 |
| SMG008F9 | >128/4 | 64 | 256 | 64 | 64 | <0.06 | 0.25 | <0.06 | 2 | >64 | 64 | 128 | 64 | <0.25 | <0.25 |
| SMG008U | >128/4 | 32 | 256 | 128 | 32 | <0.06 | <0.06 | <0.06 | 1 | >64 | 32 | 8 | <0.125 | <0.25 | <0.25 |
| SMG010F1 | 16/4 | 32 | 128 | 32 | 64 | 0.25 | 0.125 | 0.125 | 1 | 64 | 64 | 32 | 16 | <0.25 | <0.25 |
| SMG010U | >128/4 | 128 | 128 | 32 | 16 | <0.06 | <0.06 | <0.06 | 0.25 | >64 | >64 | 32 | 64 | <0.25 | <0.25 |
| SMG014F10 | >128/4 | 128 | 256 | 64 | 64 | 0.125 | 0.25 | 0.125 | 0.25 | >64 | 32 | 16 | 32 | <0.25 | <0.25 |
| SMG014F12 | >128/4 | 128 | >256 | 128 | 128 | <0.06 | 0.25 | <0.06 | 0.5 | >64 | 64 | 16 | 128 | <0.25 | <0.25 |
| SMG014U | >128/4 | 32 | 256 | 64 | 16 | <0.06 | <0.06 | <0.06 | 4 | >64 | 16 | 128 | 32 | <0.25 | <0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos-Neto, J.F.; Santos, A.C.M.; Nascimento, J.A.S.; Trovão, L.O.; Santos, F.F.; Valiatti, T.B.; Gales, A.C.; Marques, A.L.V.R.; Pinaffi, I.C.; Vieira, M.A.M.; et al. Virulence Profile, Antibiotic Resistance, and Phylogenetic Relationships among Escherichia coli Strains Isolated from the Feces and Urine of Hospitalized Patients. Pathogens 2022, 11, 1528. https://doi.org/10.3390/pathogens11121528
Santos-Neto JF, Santos ACM, Nascimento JAS, Trovão LO, Santos FF, Valiatti TB, Gales AC, Marques ALVR, Pinaffi IC, Vieira MAM, et al. Virulence Profile, Antibiotic Resistance, and Phylogenetic Relationships among Escherichia coli Strains Isolated from the Feces and Urine of Hospitalized Patients. Pathogens. 2022; 11(12):1528. https://doi.org/10.3390/pathogens11121528
Chicago/Turabian StyleSantos-Neto, José F., Ana C. M. Santos, Júllia A. S. Nascimento, Liana O. Trovão, Fernanda F. Santos, Tiago B. Valiatti, Ana C. Gales, Ana L. V. R. Marques, Isabel C. Pinaffi, Mônica A. M. Vieira, and et al. 2022. "Virulence Profile, Antibiotic Resistance, and Phylogenetic Relationships among Escherichia coli Strains Isolated from the Feces and Urine of Hospitalized Patients" Pathogens 11, no. 12: 1528. https://doi.org/10.3390/pathogens11121528