
High Parasitic Loads Quantified in Sylvatic Triatoma melanica, a Chagas Disease Vector
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field-Collected Triatomines
2.2. T. cruzi Infection by Optical Microscopy and In Vitro Cultivation
3. Molecular Assays
3.1. DNA Extraction
3.2. T. cruzi Natural Infection by PCR
3.3. Genotyping of T. cruzi DTUs by Multilocus Conventional PCR
3.4. T. cruzi Quantification by Real-Time PCR (qPCR)
3.5. Identification of the Blood Meal Sources
3.6. Statistical Analysis
4. Results
4.1. Insect Capture
4.2. T. cruzi Natural Infection Detected by Optical Microscopy and In Vitro Cultivation
4.3. T. cruzi Natural Infection Detected by PCR
4.4. Comparison of T. cruzi Detection in T. melanica Using OM and Molecular Methods
4.5. Genotyping of T. cruzi DTUs
4.6. Comparison of DTUs in Abdominal Contents and Culture Samples
4.7. Quantification of T. cruzi Parasite Load by Real-Time PCR (qPCR)
4.8. Identification of Blood Meal Sources
4.9. T. cruzi Parasite Load According to DTUs and Blood Meal Source
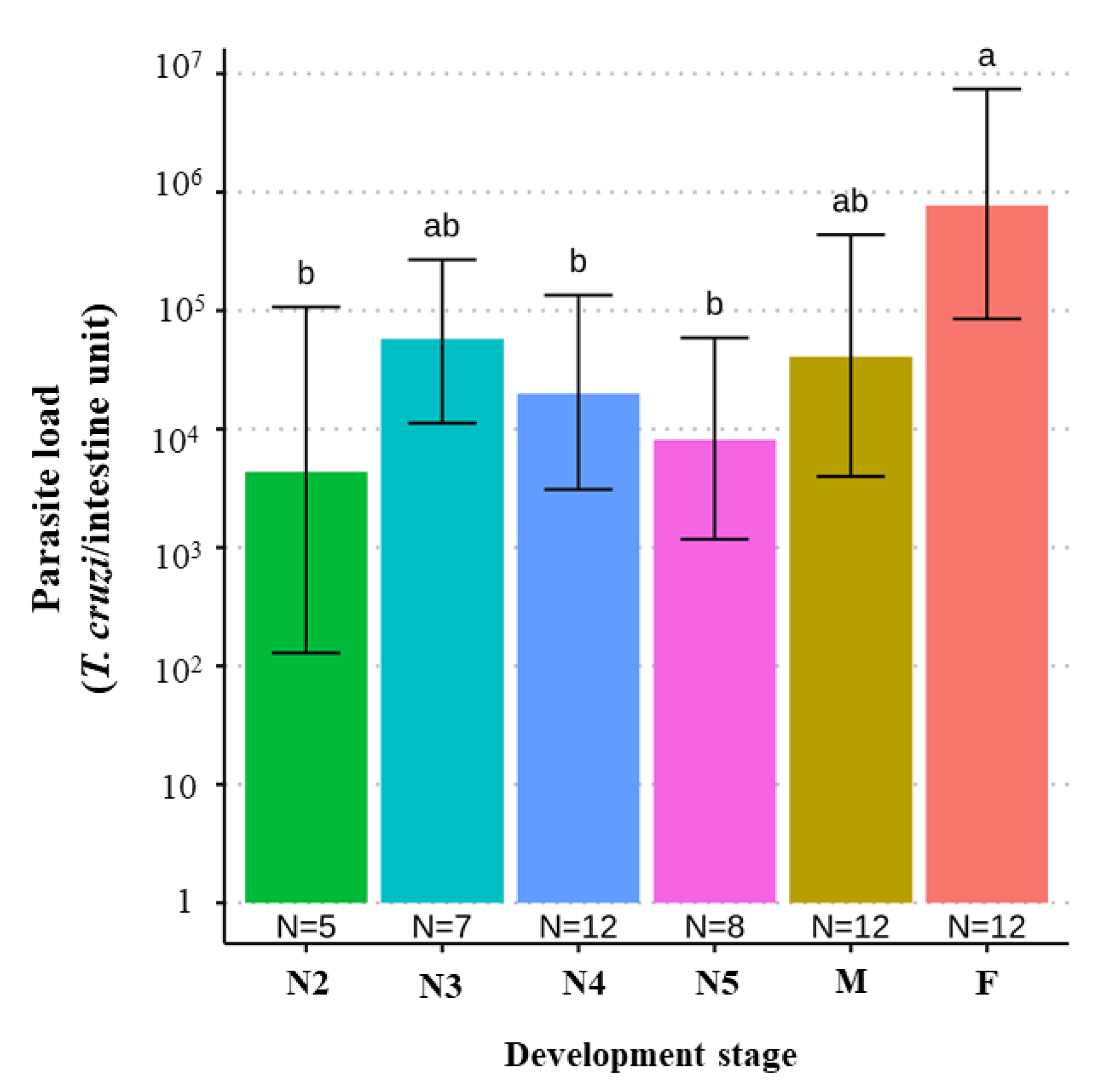
4.10. T. cruzi Parasite Load According to Triatomine Developmental Stage
4.11. Parasite Load According to T. cruzi Positivity Using OM
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zingales, B.; Bartholomeu, D.C. Trypanosoma cruzi genetic diversity: Impact on transmission cycles and Chagas disease. Mem. Inst. Oswaldo Cruz 2021, 116, e210193. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Available online: https://www.who.int/health-topics/chagas-disease (accessed on 7 October 2022).
- Dorn, P.L.; Justi, S.A.; Dale, C.; Stevens, L.; Galvão, C.; Lima-Cord´on, R.; Monroy, C. Description of Triatoma mopan sp. n. from a cave in Belize (Hemiptera, Reduviidae, Triatominae). ZooKeys 2018, 775, 69–95. [Google Scholar] [CrossRef] [PubMed]
- Lima-Cordon, R.A.; Monroy, M.C.; Stevens, L.; Rodas, A.; Rodas, G.A.; Dorn, P.L.; Justi, S.A. Description of Triatoma huehuetenanguensis sp. n., a potencial Chagas disease vector (Hemiptera, Reduviidae, Triatominae). ZooKeys 2019, 820, 51–70. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Galvão, C.; Cai, W. Rhodnius micki, a new species of Triatominae (Hemiptera, Reduviidae) from Bolivia. ZooKeys 2021, 1012, 71–93. [Google Scholar] [CrossRef] [PubMed]
- Gil-Santana, H.R.; Chavez, T.; Pita, S.; Panzera, F.; Galvão, C. Panstrongylus noireaui, a remarkable new species of Triatominae (Hemiptera, Reduviidae) from Bolivia. ZooKeys 2022, 1104, 203–225. [Google Scholar] [CrossRef]
- Diotaiuti, L.; Schofield, C.J.; Filho, O.F.F.; Carneiro, F.C.F.; Dias, J.C.P.; Pires, H.H.R. Aspectos operacionais do controle do Triatoma brasiliensis. Cad. Saude Publ. 2000, 16, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Abad-Franch, F.; Diotaiuti, L.; Gurgel-Gonçalves, R.; Gürtler, R.E. Certifying the interruption of Chagas disease transmission by native vectors: Cui bono? Mem. Inst. Oswaldo Cruz 2013, 108, 251–254. [Google Scholar] [CrossRef]
- Zingales, B.; Miles, M.A.; Campbell, D.A.; Tibayren, M.; Macedo, A.M.; Teixeira, M.M.G. The revised Trypanosoma cruzi subspecific nomenclature: Rationale, epidemiological relevance and research applications. Infect. Genet. Evol. 2012, 12, 240–253. [Google Scholar] [CrossRef]
- Zingales, B. Trypanosoma cruzi genetic diversity: Something new for something known about Chagas disease manifestations, serodiagnosis and drug sensitivity. Acta Trop. 2018, 184, 38–52. [Google Scholar] [CrossRef]
- Brenière, S.F.; Waleckx, E.; Barnabé, C. Over Six Thousand Trypanosoma cruzi Strains Classified into Discrete Typing Units (DTUs). PLoS Negl. Trop. Dis. 2016, 10, e0004792. [Google Scholar] [CrossRef]
- De Fuentes-Vicente, J.A.; Gutiérrez-Cabrera, A.E.; Flores-Villegas, A.L.; Lowenberger, C.; Benelli, G.; Salazar-Schettino, P.M.; Córdoba-Aguilar, A. What makes an effective Chagas disease vector? Factors underlying Trypanosoma cruzi-triatomine interactions. Acta Trop. 2018, 183, 23–31. [Google Scholar] [CrossRef]
- Verly, T.; Costa, S.; Lima, N.; Mallet, J.; Odêncio, F.; Pereira, M.; Moreira, C.J.C.; Britto, C.; Pavan, M.G. Vector competence and feeding-excretion behavior of Triatoma rubrovaria (Blanchard, 1843) (Hemiptera: Reduviidae) infected with Trypanosoma cruzi TcVI. PLoS Negl. Trop. Dis. 2020, 14, e0008712. [Google Scholar] [CrossRef]
- Moreira, O.C.; Verly, T.; Finamore-Araujo, P.; Gomes, S.A.O.; Lopes, C.M.; Sousa, D.M.; Azevedo, L.R.; Mota, F.F.; D’Avila-Levy, C.M.; Santos-Mallet, J.R.; et al. Development of conventional and real-time multiplex PCR-based assays for estimation of natural infection rates and Trypanosoma cruzi load in triatomine vectors. Parasites Vectors 2017, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Garcia, E.S.; Ratcliffe, N.A.; Whitten, M.M.; Gonzalez, M.S.; Azambuja, P. Exploring the role of insect host factors in the dynamics of Trypanosoma cruzi–Rhodnius prolixus interactions. J. Insect. Physiol. 2007, 53, 11–21. [Google Scholar] [CrossRef]
- Garcia, E.S.; Genta, F.A.; Azambuja, P.; Schaub, G. Interactions between intestinal compounds of triatomines and Trypanosoma cruzi. Trends Parasitol. 2010, 26, 499–505. [Google Scholar] [CrossRef]
- Valença-Barbosa, C.; Finamore-Araujo, P.; Moreira, O.C.; Vergara-Meza, J.G.; Alvarez, M.V.N.; Nascimento, J.R.; Borges-Veloso, A.; Viana, M.C.; Lilioso, M.; Miguel, D.C.; et al. Genotypic Trypanosoma cruzi distribution and parasite load differ ecotypically and according to parasite genotypes in Triatoma brasiliensis from endemic and outbreak areas in Northeastern Brazil. Acta Trop. 2021, 222, 106054. [Google Scholar] [CrossRef]
- Monteiro, F.A.; Donnelly, M.J.; Beard, C.B.; Costa, J. Nested clade and phylogeographic analyses of the Chagas disease vector Triatoma brasiliensis in Northeast Brazil. Mol. Phylogenet. Evol. 2004, 32, 46–56. [Google Scholar] [CrossRef]
- Almeida, C.E.; Oliveira, H.L.; Correia, N.; Dornak, L.L.; Gumiel, M.; Neiva, V.L.; Harry, M.; Mendonça, V.J.; Costa, J.; Galvão, C. Dispersion capacity of Triatoma sherlocki, Triatoma juazeirensis and laboratory-bred hybrids. Acta Trop. 2012, 122, 71–79. [Google Scholar] [CrossRef]
- Mendonça, V.J.; da Silva, M.T.; de Araujo, R.F.; Martins, J., Jr.; Bacci, M., Jr.; Almeida, C.E. Phylogeny of Triatoma sherlocki (Hemiptera: Reduviidae: Triatominae) inferred from two mitochondrial genes suggests its location within the Triatoma brasiliensis complex. Am. J. Trop. Med. Hyg. 2009, 81, 858–864. [Google Scholar] [CrossRef]
- Costa, J.; Dornak, L.L.; Almeida, C.D.; Townsend-Peterson, A. Distributional potential of the Triatoma brasiliensis species complex at present and under scenarios of future climate conditions. Parasite Vectors 2014, 7, 238. [Google Scholar] [CrossRef]
- Souza, R.C.M.; Campolina-Silva, G.H.; Bezerra, C.M.; Diotaiuti, L.; Gorla, D.E. Does Triatoma brasiliensis occupy the same environmental niche space as Triatoma melanica? Parasite Vectors 2015, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa-Silva, A.N.; Câmara, A.C.J.; Martins, K.; Nunes, D.F.; de Oliveira, P.I.C.; de Azevedo, P.R.M.; Chiari, E.; Galvão, L.M.C. Characteristics of triatomine infestation and natural Trypanosoma cruzi infection in the state of Rio Grande do Norte, Brazil. Rev. Soc. Bras. Med. Trop. 2016, 49, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, C.M.; Barbosa, S.E.; Souza, R.C.M.; Barezani, C.P.; Gürtler, R.E.; Ramos, A.N.; Diotaiuti, L. Triatoma brasiliensis Neiva, 1911: Food sources and diversity of Trypanosoma cruzi in wild and artificial environments of the semiarid region of Ceará, northeastern Brazil. Parasite Vectors 2018, 11, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezerra, C.M.; Barbosa, S.E.; Souza, R.C.M.; Feijão, L.X.; Gürtler, R.E.; Ramos, A.N.; Diotaiuti, L. Fast recovery of house infestation with Triatoma brasiliensis after residual insecticide spraying in a semiarid region of Northeastern Brazil. PLoS Negl. Trop. Dis. 2020, 14, e0008404. [Google Scholar] [CrossRef] [PubMed]
- Monsalve-Lara, J.; Lilioso, M.; Valença-Barbosa, C.; Thyssen, P.J.; Miguel, D.C.; Limeira, C.; Gadelha, F.R.; Fontes, F.V.H.; Pires-Silva, D.; Dornak, L.L.; et al. The risk of oral transmission in an area of a Chagas disease outbreak in the Brazilian northeast evaluated through entomological, socioeconomic and schooling indicators. Acta Trop. 2021, 215, 105803. [Google Scholar] [CrossRef]
- Costa, J.; Almeida, C.E.; Dotson, E.M.; Lins, A.; Vinhaes, M.; Silveira, A.C.; Beard, C.B. The epidemiologic importance of Triatoma brasiliensis as a Chagas disease vector in Brazil: A revision of domiciliary captures during 1993–1999. Mem. Inst. Oswaldo Cruz 2003, 98, 443–449. [Google Scholar] [CrossRef]
- Almeida, C.E.; Peterson, A.T.; Costa, J.; Duarte, R.; Lima, M.M.; Locks, M.; Beltrão, M.; Costa, J. Could the bug Triatoma sherlocki be vectoring Chagas disease in small mining communities in Bahia, Brazil. Med. Vet. Entomol. 2009, 23, 410–417. [Google Scholar] [CrossRef]
- Dale, C.; Almeida, C.E.; Mendonça, V.J.; Oliveira, J.; Rosa, J.A.; Galvão, C.; Costa, J. An updated and illustrated dichotomous key for the Triatoma brasiliensis species complex and their epidemiologic importance. Zookeys 2018, 805, 33–43. [Google Scholar] [CrossRef]
- Folly-Ramos, E.; Dornak, L.L.; Orsolon, G.; Gonçalves, T.C.M.; Lilioso, M.; Costa, J.; Almeida, C.E. Vector capacity of members of Triatoma brasiliensis species complex: The need to extend Chagas disease surveillance to Triatoma melanica. J. Vector Ecol. 2016, 41, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Ab’Saber, A.N. Espaços ocupados pela expansão dos climas secos na América do Sul, por ocasião dos períodos glaciais quaternários. Paleoclimas Inst. Geogr. Univ. São Paulo 1977, 3, 1–19. [Google Scholar]
- Chiari, E.; Camargo, E.P. Culture and cloning of Trypanosoma cruzi. In Genes and Antigens of Parasites, a Laboratory Manual; Morel, C.M., Ed.; Fundação Oswaldo Cruz: Rio de Janeiro, Brasil; World Health Organization: Geneva, Switzerland, 1984; pp. 23–26. [Google Scholar]
- Sturm, N.R.; Degrave, W.; Morel, C.; Simpson, L. Sensitive detection and schizodeme classification of Trypanosoma cruzi cells by amplification of kinetoplast minicircle DNA sequences: Use in diagnosis of Chagas’ disease. Mol. Biochem. Parasitol. 1989, 33, 205–214. [Google Scholar] [CrossRef]
- Wincker, P.; Britto, C.; Pereira, J.B.; Cardoso, M.A.; Oelemann, W.; Morel, C.M. Use of a Simplified Polymerase Chain Reaction Procedure to Detect Trypanosoma cruzi in Blood Samples from Chronic Chagasic Patients in a Rural Endemic Area. Am. J. Trop. Med. Hyg. 1994, 51, 771–777. [Google Scholar] [CrossRef]
- Moreira, O.C.; Ramirez, J.C. Genotyping of Trypanosoma cruzi from Clinical Samples by Multilocus Conventional PCR. Methods Mol. Biol. 2019, 1955, 227–238. [Google Scholar]
- Piron, M.; Fisa, R.; Casamitjana, N.; Lopez-Chejade, P.; Puig, L.; Verges, M.; Gascon, J.; Prat, J.G.; Portús, M.; Sauleda, S. Development of a real-time PCR assay for Trypanosoma cruzi detection in blood samples. Acta Trop. 2007, 103, 195–200. [Google Scholar] [CrossRef]
- Kitano, T.; Umetsu, K.; Tian, W.; Osawa, M. Two universal primer sets for species identification among vertebrates. Int. J. Legal Med. 2007, 21, 423–427. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 21 September 2022).
- R Studio Team. RStudio: Integrated Development for R; RStudio. Inc.: Boston, MA, USA, 2015; Available online: http://www.rstudio.com/ (accessed on 21 September 2022).
- Nagelkerke, N.J. A note on a general definition of the coefficient of determination. Biometrika 1991, 78, 691–692. [Google Scholar] [CrossRef]
- Landis, J.R.; Koch, G.G. An application of hierarchical kappa-type statistics in the assessment of majority agreement among multiple observers. Biometrics 1977, 33, 363–374. [Google Scholar] [CrossRef]
- Paula, R.C. Impact of Anthropogenic Activities on Mammal Diversity and Abundance in the Cerrado of Brazil. Ph.D. Thesis, Florida Atlantic University, Boca Raton, FL, USA, 2004. [Google Scholar]
- Oliveira, V.B.; Câmara, E.M.; Oliveira, L.C. Composição e caracterização da mastofauna de médio e grande porte do Parqu Nacional da Serra do Cipó, Minas Gerais, Brasil. Mastozool. Neotrop. 2009, 16, 355–364. [Google Scholar]
- SVS/Secretaria de Vigilância em Saúde/Ministério da Saúde (2020). Doença de Chagas. Available online: www.saude.gov.br/images/pdf/2020/April/23/boletim-especial-chagas20abr20.pdf (accessed on 15 October 2022).
- Araujo-Jorge, T.C.; Ferreira, R.R.; Rocha, R.C.M.; Vieira, T.M.; Costa, N.D.; Santos, L.L.; Silva, J.O.; Mendes, M.O.; Almeida-Silva, J.; Costa, E.J.; et al. Chagas Express XXI”: A new ArtScience social technology for health and science education—A case study in Brazilian endemic areas of Chagas disease with an active search of chronic cases. PLoS Negl. Trop. Dis. 2021, 15, e0009534. [Google Scholar] [CrossRef]
- Diotaiuti, L.; Pereira, A.S.; Loiola, C.F.; Fernandes, A.J.; Schofield, J.C.; Dujardin, J.P.; Dias, J.C.; Chiari, E. Inter-relation of sylvatic and domestic transmission of Trypanosoma cruzi in areas with and without domestic vectorial transmission in Minas Gerais, Brazil. Mem. Inst. Oswaldo Cruz 1995, 90, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.; Correia, N.C.; Neiva, V.L.; Gonçalves, T.C.M.; Felix, M. Revalidation and redescription of Triatoma brasiliensis macromelasoma Galvão, 1956 and an identification key for the Triatoma brasiliensis complex (Hemiptera: Reduviidae: Triatominae). Mem. Inst. Oswaldo Cruz 2013, 108, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, M.; Bacigalupo, A.; Victoria, M.; Vergara, M.J.; Alvarez-Duhart, B.; Muñoz-San, C.M.; Solis, R.; Cattan, P.E. Trypanosoma cruzi infection in the wild Chagas disease vector, Mepraia spinolai: Parasitic load, discrete typing units, and blood meal sources. Acta Trop. 2022, 229, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sarquis, O.; Borges-Pereira, J.; Mac Cord, J.R.; Gomes, T.F.; Cabello, P.H.; Lima, M.M. Epidemiology of Chagas disease in Jaguaruana, Ceara, Brazil. I. Presence of triatomines and index of Trypanosoma cruzi infection in four localities of a rural area. Mem. Inst. Oswaldo Cruz 2004, 99, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Lilioso, M.; Reigada, C.; Pires-Silva, D.; Fontes, F.V.H.M.; Limeira, C.; Monsalve-Lara, J.; Folly-Ramos, E.; Harry, M.; Costa, J.; Almeida, C.E. Dynamics of food sources, ecotypic distribution and Trypanosoma cruzi infection in Triatoma brasiliensis from the northeast of Brazil. PLoS Negl. Trop. Dis. 2020, 14, e0008735. [Google Scholar] [CrossRef] [PubMed]
- Herrera, L.; Andrea, P.S.D.; Xavier, S.C.C.; Mangia, R.H.; Fernandes, O.; Jansen, A.M. Trypanosoma cruzi infection in wild mammals of the National Park ‘Serra da Capivara’ and its surroundings (Piauí, Brazil), an area endemic for Chagas disease. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 379–388. [Google Scholar] [CrossRef]
- Roque, A.L.; D’Andrea, P.S.; Andrade, G.B.; Jansen, A.M. Trypanosoma cruzi: Distinct patterns of infection in the sibling caviomorph rodent species Thrichomys apereoides laurentius and Thrichomys pachyurus (Rodentia, Echimyidae). Exp. Parasitol. 2005, 111, 37–46. [Google Scholar] [CrossRef]
- Waniek, P.J.; Araújo, C.A.C.; Jansen, A.M.; Costa, J. First genotyping of Trypanosoma cruzi from naturally infected Triatoma juazeirensis, Triatoma melanica and Triatoma sherlocki from Bahia State, Brazil. Med. Vet. Entomol. 2021, 35, 134–140. [Google Scholar] [CrossRef]
- Valença-Barbosa, C.; Lima, M.M.; Sarquis, O.; Bezerra, C.M.; Abad-Franch, F. Modeling disease vector occurrence when detection is imperfect II: Drivers of site occupancy by synanthropic Triatoma brasiliensis in the Brazilian Northeast. PLoS Negl. Trop. Dis. 2014, 8, e2861. [Google Scholar] [CrossRef] [Green Version]
- Almeida, C.E.; Faucher, L.; Lavina, M.; Costa, J.; Harry, M. Molecular individual-based approach on Triatoma brasiliensis: Inferences on triatomine foci, Trypanosoma cruzi natural infection prevalence, parasite diversity and feeding sources. PLoS Negl. Trop. Dis. 2016, 10, e0004447. [Google Scholar] [CrossRef] [Green Version]
- Llewellyn, M.S.; Lewis, M.D.; Acosta, N.; Yeo, M.; Carrasco, H.J.; Segovia, M.; Vargas, J.; Torrico, F.; Miles, M.A.; Gaunt, M.W. Trypanosoma cruzi IIc: Phylogenetic and phylogeographic insights from sequence and microsatellite analysis and potential impact on emergent Chagas disease. PLoS. Negl. Trop. Dis. 2009, 3, e510. [Google Scholar] [CrossRef] [Green Version]
- Marcili, A.; Valente, V.C.; Valente, S.A.; Junqueira, A.C.V.; Silva, F.M.; Pinto, A.Y.N.; Naiff, R.D.; Campaner, M.; Coura, J.R.; Camargo, E.P.; et al. Trypanosoma cruzi in Brazilian Amazonia: Lineages TCI and TCIIa in wild primates, Rhodnius spp. and in humans with Chagas disease associated with oral transmission. Int. J. Parasitol. 2009, 39, 615–623. [Google Scholar] [CrossRef]
- Burgos, J.M.; Altcheh, J.; Bisio, M.; Duffy, T.; Valadares, H.M.; Seidenstein, M.E.; Piccinali, R.; Freitas, J.M.; Levin, M.J.; Macchi, L.; et al. Direct molecular profiling of minicircle signatures and lineages of Trypanosoma cruzi bloodstream populations causing congenital Chagas disease. Int. J. Parasitol. 2007, 37, 1319–1327. [Google Scholar] [CrossRef]
- Cardinal, M.V.; Lauricella, M.A.; Ceballos, L.A.; Lanati, L.; Marcet, P.L.; Levin, M.J.; Kitron, U.; Gurtler, R.; Alejandro, G.S. Molecular epidemiology of domestic and sylvatic Trypanosoma cruzi infection in rural northwestern Argentina. Int. J. Parasitol. 2008, 38, 1533–1543. [Google Scholar] [CrossRef]
- D’Avila, D.A.; Macedo, A.M.; Valadares, H.M.; Gontijo, E.D.; de Castro, A.M.; Machado, C.R.; Chiari, E.; Galvão, L.M. Probing population dynamics of Trypanosoma cruzi during progression of the chronic phase in chagasic patients. J. Clin. Microbiol. 2009, 47, 1718–1725. [Google Scholar] [CrossRef] [Green Version]
- Câmara, A.C.J.; Varela-Freire, A.A.; Valadares, H.M.S.; Macedo, A.M.; D’Avila, D.A.; Machado, C.R.; Lages-Silva, E.; Chiari, E.; Galvão, L.M.C. Genetic analyses of Trypanosoma cruzi isolates from naturally infected triatomines and humans in northeastern Brazil. Acta Trop. 2010, 115, 205–211. [Google Scholar] [CrossRef]
- Fernandes, O.; Sturm, N.R.; Derré, R.; Campbell, D.A. The mini-exon gene: A genetic marker for zymodeme III of Trypanosoma cruzi. Mol. Biochem. Parasitol. 1998, 95, 129–133. [Google Scholar] [CrossRef]
- Araújo, C.A.C.; Cabello, P.H.; Jansen, A.M. Growth behaviour of two Trypanosoma cruzi strains in single and mixed infections: In vitro and in the intestinal tract of the blood-sucking bug, Triatoma brasiliensis. Acta Trop. 2007, 101, 225–231. [Google Scholar] [CrossRef]
- Davey, K.G. The interaction of feeding and mating in the hormonal control of egg production in Rhodnius prolixus. J. Insect Physiol. 2007, 57, 208–215. [Google Scholar] [CrossRef]
- Vinhaes, M.C.; Oliveira, S.V.; Reis, P.O.; Sousa, A.C.L.; Silva, R.A.; Obara, M.T.; Bezerra, C.M.; Costa, V.M.; Alves, R.V.; Gurgel-Gonçalves, R. Assessing the vulnerability of Brazilian municipalities to the vectorial transmission of Trypanosoma cruzi using multi-criteria decision analysis. Acta Trop. 2014, 137, 105–110. [Google Scholar] [CrossRef]
- Minuzzi-Souza, T.T.C.; Silva, L.R.; Hagstrom, L.; Hecht, M.; Nitz, N.; Gurgel- Gonçalves, R. Molecular bloodmeal analyses reveal that Trypanosoma cruzi-infected, native triatomine bugs often feed on humans in houses in central Brazil. Med. Vet. Entomol. 2018, 32, 504–508. [Google Scholar] [CrossRef]
- Cominetti, M.C.; Almeida, R.F.; Gonçalves, G.M.; Andreotti, R. Monitoring Trypanosoma cruzi infection in triatomines using PCR in Mato Grosso do Sul, Brazil. Rev. Soc. Bras. Med. Trop. 2013, 3, 277–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima-Oliveira, T.M.; Fontes, F.V.H.M.; Lilioso, M.; Pires-Silva, D.; Teixeira, M.M.G.; Meza, J.G.V.; Harry, M.; Filee, J.; Costa, J.; Valenca-Barbosa, C.; et al. Molecular eco-epidemiology on the sympatric Chagas disease vectors Triatoma brasiliensis and Triatoma petrocchiae: Ecotopes, genetic variation, natural infection prevalence by trypanosomatids and parasite genotyping. Acta Trop. 2020, 201, 105188. [Google Scholar] [CrossRef] [PubMed]
- Abad-Franch, F.; Veja, M.C.; Rolon, M.S.; Santos, W.S.; Arias, A.R. Community Participation in Chagas Disease Vector Surveillance: Systematic Review. PLoS Negl. Trop. Dis. 2011, 5, e1207. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valença-Barbosa, C.; Finamore-Araujo, P.; Moreira, O.C.; Alvarez, M.V.N.; Borges-Veloso, A.; Barbosa, S.E.; Diotaiuti, L.; de Souza, R.d.C.M. High Parasitic Loads Quantified in Sylvatic Triatoma melanica, a Chagas Disease Vector. Pathogens 2022, 11, 1498. https://doi.org/10.3390/pathogens11121498
Valença-Barbosa C, Finamore-Araujo P, Moreira OC, Alvarez MVN, Borges-Veloso A, Barbosa SE, Diotaiuti L, de Souza RdCM. High Parasitic Loads Quantified in Sylvatic Triatoma melanica, a Chagas Disease Vector. Pathogens. 2022; 11(12):1498. https://doi.org/10.3390/pathogens11121498
Chicago/Turabian StyleValença-Barbosa, Carolina, Paula Finamore-Araujo, Otacílio Cruz Moreira, Marcus Vinicius Niz Alvarez, André Borges-Veloso, Silvia Ermelinda Barbosa, Liléia Diotaiuti, and Rita de Cássia Moreira de Souza. 2022. "High Parasitic Loads Quantified in Sylvatic Triatoma melanica, a Chagas Disease Vector" Pathogens 11, no. 12: 1498. https://doi.org/10.3390/pathogens11121498