

Viral Metagenomics Reveals a Putative Novel HPV Type in Anogenital Wart Tissues


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.2. Viral Metagenomic and Bioinformatics Analysis
2.3. Acquiring of Complete Genome and PCR Reactions
2.4. Phylogenetic Analysis
3. Results
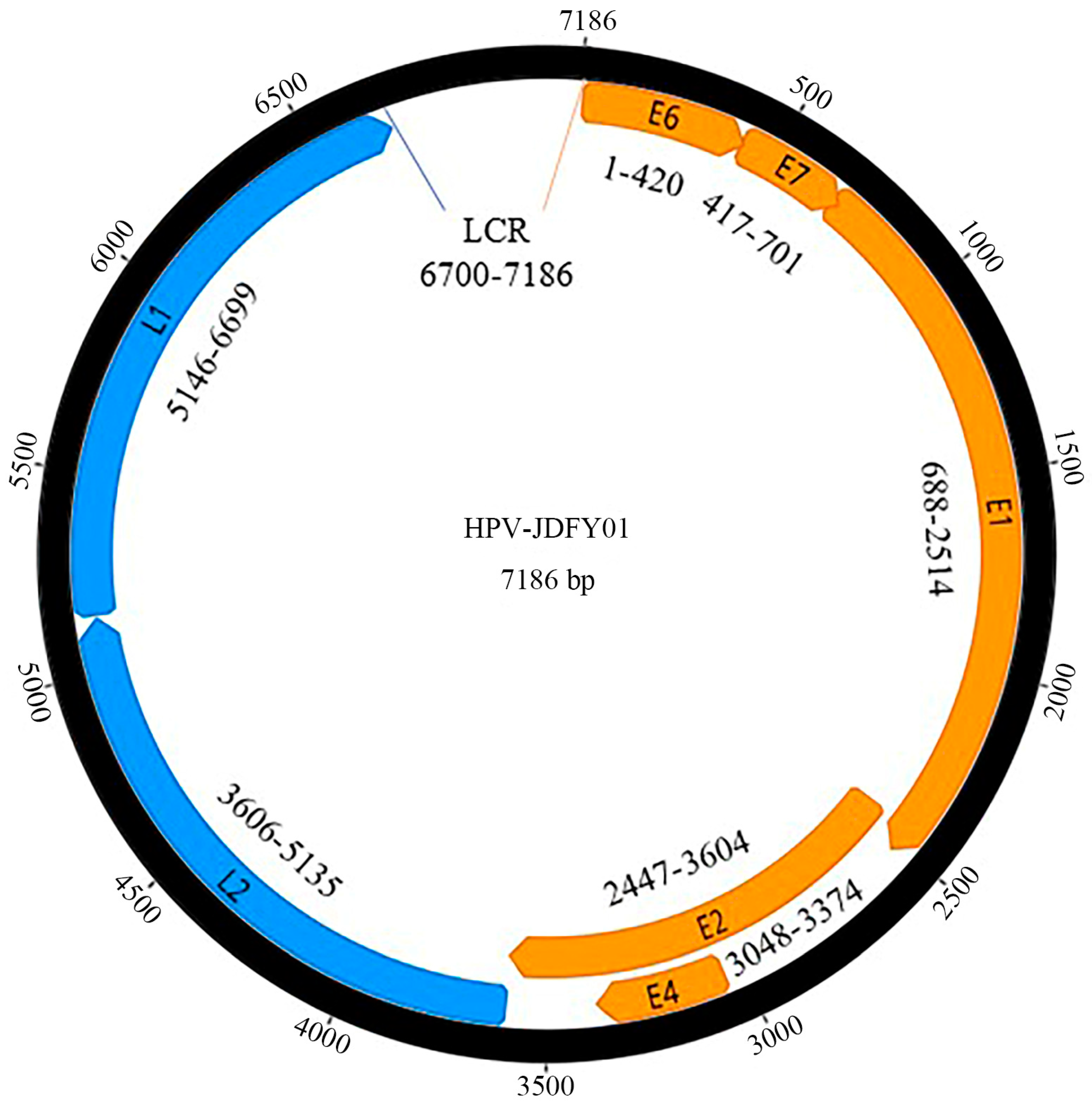
3.1. Complete Genomic Structure of HPV-JDFY01
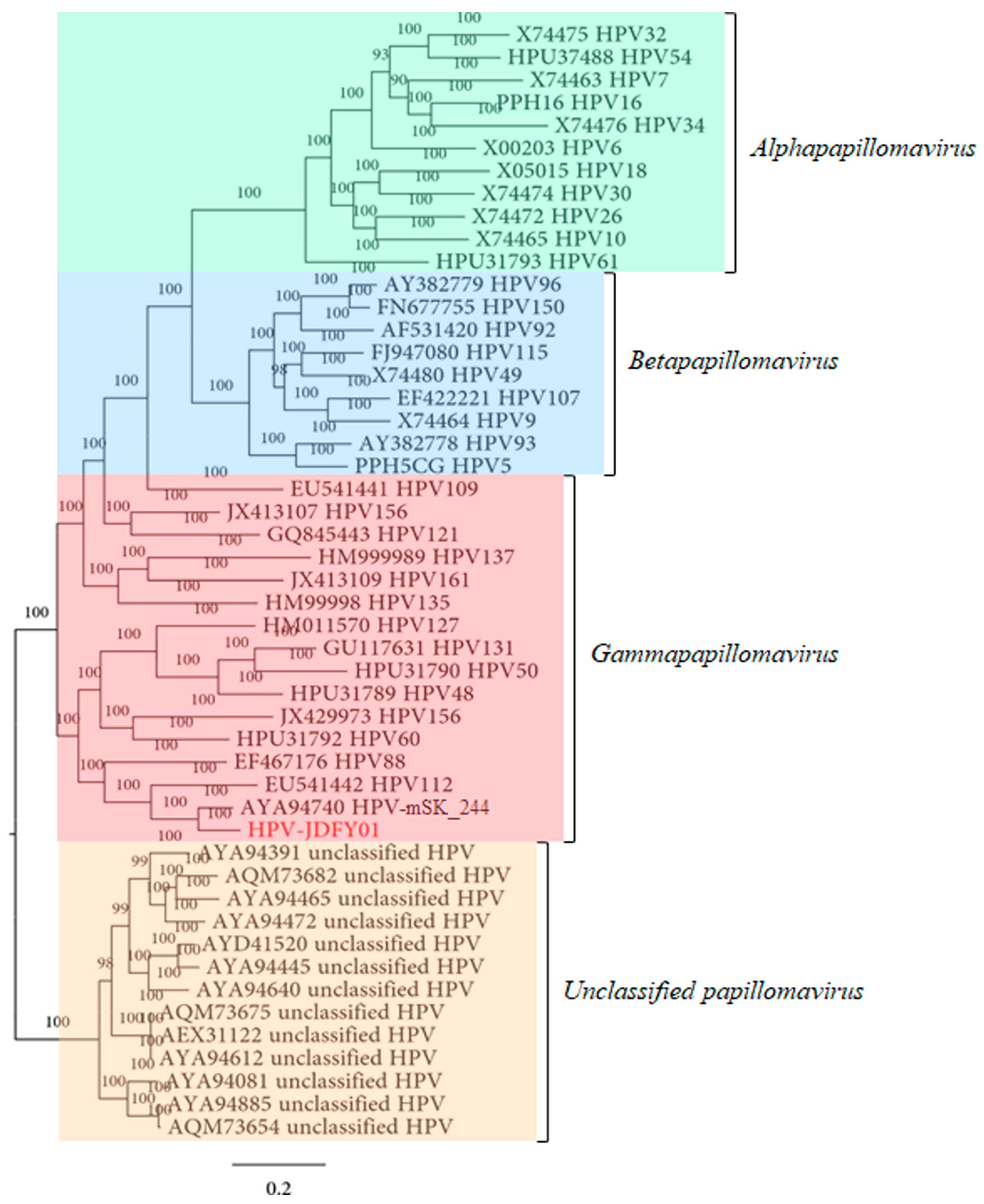
3.2. Phylogenetic Analysis of HPV-JDFY01
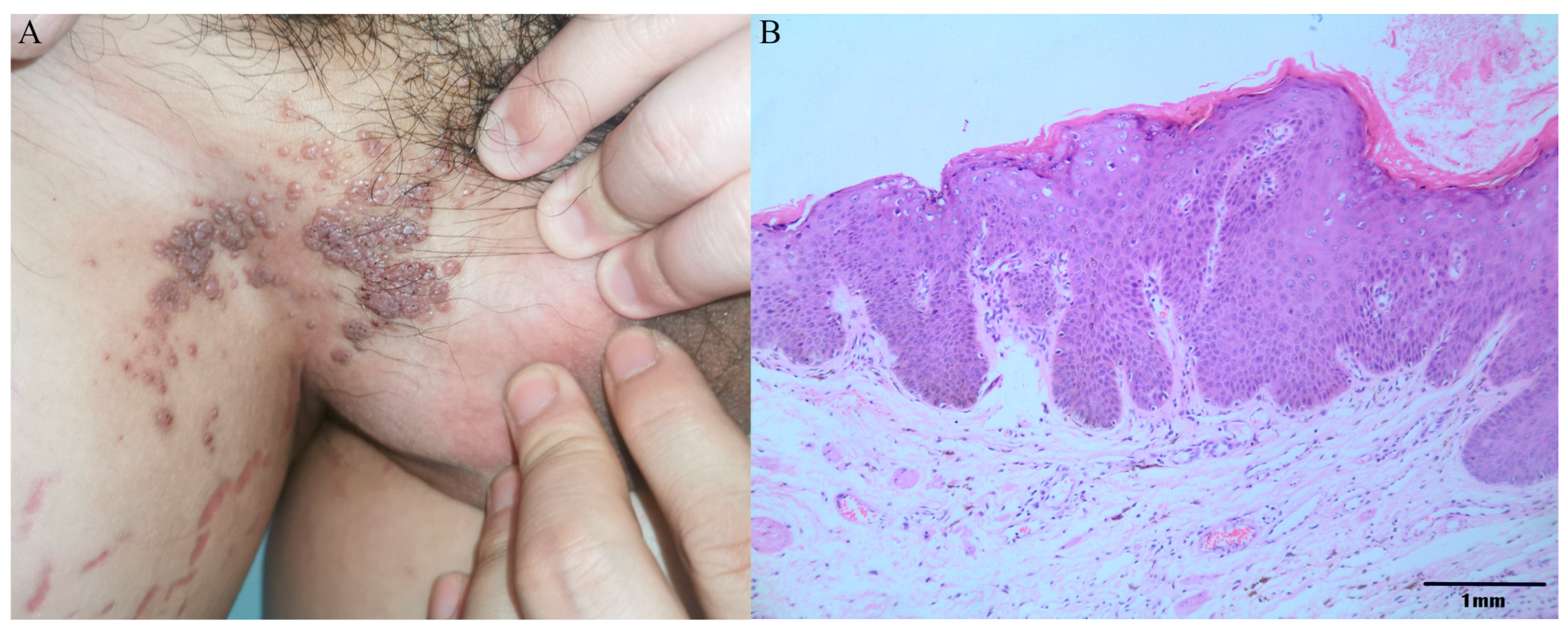
3.3. Epidemiological Analysis of HPV-JDFY01
3.4. GenBank Accession Number
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Villiers, E.M.; Fauquet, C.; Broker, T.R.; Bernard, H.U.; zur Hausen, H. Classification of papillomaviruses. Virology 2004, 324, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seedorf, K.; Krammer, G.; Durst, M.; Suhai, S.; Rowekamp, W.G. Human papillomavirus type 16 DNA sequence. Virology 1985, 145, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Bernard, H.-U.; Burk, R.D.; Chen, Z.; van Doorslaer, K.; Hausen, H.z.; de Villiers, E.-M. Classification of papillomaviruses (PVs) based on 189 PV types and proposal of taxonomic amendments. Virology 2010, 401, 70–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, C.C.; Phelps, W.C.; Lindgren, V.; Braun, M.J.; Gonda, M.A.; Howley, P.M. Structural and transcriptional analysis of human papillomavirus type 16 sequences in cervical carcinoma cell lines. J. Virol. 1987, 61, 962–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howley, P.M.; Munger, K.; Werness, B.A.; Phelps, W.C.; Schlegel, R. Molecular mechanisms of transformation by the human papillomaviruses. Princess Takamatsu Symp. 1989, 20, 199–206. [Google Scholar] [PubMed]
- Gheit, T. Mucosal and Cutaneous Human Papillomavirus Infections and Cancer Biology. Front. Oncol. 2019, 9, 355. [Google Scholar] [CrossRef] [Green Version]
- Tirosh, O.; Conlan, S.; Deming, C.; Lee-Lin, S.Q.; Huang, X.; Program, N.C.S.; Su, H.C.; Freeman, A.F.; Segre, J.A.; Kong, H.H. Expanded skin virome in DOCK8-deficient patients. Nat. Med. 2018, 24, 1815–1821. [Google Scholar] [CrossRef]
- Muhr, L.S.A.; Eklund, C.; Dillner, J. Towards quality and order in human papillomavirus research. Virology 2018, 519, 74–76. [Google Scholar] [CrossRef]
- Van Doorslaer, K.; McBride, A.A. Molecular archeological evidence in support of the repeated loss of a papillomavirus gene. Sci. Rep. 2016, 6, 33028. [Google Scholar] [CrossRef]
- Pastrana, D.V.; Peretti, A.; Welch, N.L.; Borgogna, C.; Olivero, C.; Badolato, R.; Notarangelo, L.D.; Gariglio, M.; FitzGerald, P.C.; McIntosh, C.E.; et al. Metagenomic Discovery of 83 New Human Papillomavirus Types in Patients with Immunodeficiency. mSphere 2018, 3, e00645-18. [Google Scholar] [CrossRef]
- Latsuzbaia, A.; Arbyn, M.; Dutta, S.; Fischer, M.; Gheit, T.; Tapp, J.; Tommasino, M.; Weyers, S.; Mossong, J. Complete Genome Sequence of a Novel Human Gammapapillomavirus Isolated from a Cervical Swab in Luxembourg. Genome Announc. 2018, 6, e00114-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brancaccio, R.N.; Robitaille, A.; Dutta, S.; Rollison, D.E.; Tommasino, M.; Gheit, T. Isolation of a Novel Beta-2 Human Papillomavirus from Skin. Microbiol. Resour. Announc. 2019, 8, e01628-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Yang, S.; Wang, Y.; Shen, Q.; Yang, Y.; Deng, X.; Zhang, W.; Delwart, E. Identification of a novel human papillomavirus by metagenomic analysis of vaginal swab samples from pregnant women. Virol. J. 2016, 13, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, Y.; Wang, J.; Yin, J.; Xu, J.; Wu, Y.; Zhou, R.; Lu, J.; Yang, S.; Wang, X.; Shen, Q.; et al. Genomic organization of a Gamma-6 papillomavirus metagenomic discovered from vaginal swab samples of Chinese pregnant women. Virol. J. 2020, 17, 44. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Li, L.; Deng, X.; Blumel, J.; Nubling, C.M.; Hunfeld, A.; Baylis, S.A.; Delwart, E. Viral nucleic acids in human plasma pools. Transfusion 2016, 56, 2248–2255. [Google Scholar] [CrossRef]
- Zhang, W.; Yang, S.; Shan, T.; Hou, R.; Liu, Z.; Li, W.; Guo, L.; Wang, Y.; Chen, P.; Wang, X.; et al. Virome comparisons in wild-diseased and healthy captive giant pandas. Microbiome 2017, 5, 90. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, S.; Mahmood, A.; Yang, S.; Wang, X.; Shen, Q.; Shan, T.; Deng, X.; Li, J.; Hua, X.; et al. Plasma virome of cattle from forest region revealed diverse small circular ssDNA viral genomes. Virol. J. 2018, 15, 11. [Google Scholar] [CrossRef] [Green Version]
- Sabatier, M.; Bal, A.; Destras, G.; Regue, H.; Queromes, G.; Cheynet, V.; Lina, B.; Bardel, C.; Brengel-Pesce, K.; Navratil, V.; et al. Comparison of Nucleic Acid Extraction Methods for a Viral Metagenomics Analysis of Respiratory Viruses. Microorganisms 2020, 8, 1539. [Google Scholar] [CrossRef]
- Xiao, Y.; Wang, H.; Feng, L.; Pan, J.; Chen, Z.; Wang, H.; Yang, S.; Shen, Q.; Wang, X.; Shan, T.; et al. Fecal, oral, blood and skin virome of laboratory rabbits. Arch. Virol. 2020, 165, 2847–2856. [Google Scholar] [CrossRef]
- Pichler, M.; Coskun, O.K.; Ortega-Arbulu, A.S.; Conci, N.; Worheide, G.; Vargas, S.; Orsi, W.D. A 16S rRNA gene sequencing and analysis protocol for the Illumina MiniSeq platform. Microbiologyopen 2018, 7, e00611. [Google Scholar] [CrossRef]
- McGinnis, S.; Madden, T.L. BLAST: At the core of a powerful and diverse set of sequence analysis tools. Nucleic Acids Res. 2004, 32, W20–W25. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Naccache, S.; Ng, T.; Federman, S.; Li, L.; Chiu, C.; Delwart, E. An ensemble strategy that significantly improves de novo assembly of microbial genomes from metagenomic next-generation sequencing data. Nucleic Acids Res. 2015, 43, e46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bibi, Z.; Fatima, A.; Rani, R.; Maqbool, A.; Khan, S.; Naz, S.; Waseem, S. Genetic characterization of Plasmodium vivax isolates from Pakistan using circumsporozoite protein (pvcsp) and merozoite surface protein-1 (pvmsp-1) genes as genetic markers. Malar. J. 2021, 20, 112. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wang, H.; Ling, Y.; Yang, S.X.; Wang, X.C.; Zhou, R.; Xiao, Y.Q.; Chen, X.; Yang, J.; Fu, W.G.; et al. Viral metagenomics revealed diverse CRESS-DNA virus genomes in faeces of forest musk deer. Virol. J. 2020, 17, 61. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Grennan, D. Genital Warts. JAMA 2019, 321, 520. [Google Scholar] [CrossRef]
- Domenech-Vinolas, M.; Leon-Maldonado, L.; Ramirez-Palacios, P.; Flores, Y.N.; Granados-Garcia, V.; Brown, B.; Corona, E.; Liu, S.; Lazcano-Ponce, E.; Salmeron, J. Incidence, psychosocial burden, and economic impact of genital warts in Mexico. Salud Publica Mex. 2018, 60, 624–632. [Google Scholar] [CrossRef]
- Ogawa, T.; Tomita, Y.; Okada, M.; Shirasawa, H. Complete genome and phylogenetic position of bovine papillomavirus type 7. J. Gen. Virol. 2007, 88, 1934–1938. [Google Scholar] [CrossRef]
- Bernard, H.U. Regulatory elements in the viral genome. Virology 2013, 445, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Kohler, A.; Gottschling, M.; Manning, K.; Lehmann, M.D.; Schulz, E.; Kruger-Corcoran, D.; Stockfleth, E.; Nindl, I. Genomic characterization of ten novel cutaneous human papillomaviruses from keratotic lesions of immunosuppressed patients. J. Gen. Virol. 2011, 92, 1585–1594. [Google Scholar] [CrossRef]
- McLaughlin-Drubin, M.E.; Munger, K. The human papillomavirus E7 oncoprotein. Virology 2009, 384, 335–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergvall, M.; Melendy, T.; Archambault, J. The E1 proteins. Virology 2013, 445, 35–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.W.; Roden, R.B. L2, the minor capsid protein of papillomavirus. Virology 2013, 445, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lace, M.J.; Isacson, C.; Anson, J.R.; Lorincz, A.T.; Wilczynski, S.P.; Haugen, T.H.; Turek, L.P. Upstream regulatory region alterations found in human papillomavirus type 16 (HPV-16) isolates from cervical carcinomas increase transcription, ori function, and HPV immortalization capacity in culture. J. Virol. 2009, 83, 7457–7466. [Google Scholar] [CrossRef] [Green Version]
- Gu, W.; Miller, S.; Chiu, C.Y. Clinical Metagenomic Next-Generation Sequencing for Pathogen Detection. Annu. Rev. Pathol. 2019, 14, 319–338. [Google Scholar] [CrossRef]
- Liao, Z.H.; Yang, S.; Li, Q.; Lu, H.Z. The role of metagenomic next-generation sequencing as a promising technology for diagnosing HIV-TB coinfection. Zoonoses 2022, 2, 32. [Google Scholar] [CrossRef]
- Slavov, S.N. Viral Metagenomics for Identification of Emerging Viruses in Transfusion Medicine. Viruses 2022, 14, 2448. [Google Scholar] [CrossRef]
- Miller, S.; Chiu, C. The Role of Metagenomics and Next-Generation Sequencing in Infectious Disease Diagnosis. Clin. Chem. 2021, 68, 115–124. [Google Scholar] [CrossRef]
- Ekstrom, J.; Forslund, O.; Dillner, J. Three novel papillomaviruses (HPV109, HPV112 and HPV114) and their presence in cutaneous and mucosal samples. Virology 2010, 397, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Pan, Y.; Deng, Q.; Cai, H.; Ke, Y. Identification and characterization of eleven novel human gamma-papillomavirus isolates from healthy skin, found at low frequency in a normal population. PLoS ONE 2013, 8, e77116. [Google Scholar] [CrossRef]
- Li, J.; Cai, H.; Xu, Z.; Wang, Q.; Hang, D.; Shen, N.; Liu, M.; Zhang, C.; Abliz, A.; Ke, Y. Nine complete genome sequences of cutaneous human papillomavirus genotypes isolated from healthy skin of individuals living in rural He Nan province, China. J. Virol. 2012, 86, 11936. [Google Scholar] [CrossRef] [Green Version]
- Hatterschide, J.; Bohidar, A.E.; Grace, M.; Nulton, T.J.; Kim, H.W.; Windle, B.; Morgan, I.M.; Munger, K.; White, E.A. PTPN14 degradation by high-risk human papillomavirus E7 limits keratinocyte differentiation and contributes to HPV-mediated oncogenesis. Proc. Natl. Acad Sci. USA 2019, 116, 7033–7042. [Google Scholar] [CrossRef] [Green Version]
- Zou, N.; Lin, B.Y.; Duan, F.; Lee, K.Y.; Jin, G.; Guan, R.; Yao, G.; Lefkowitz, E.J.; Broker, T.R.; Chow, L.T. The hinge of the human papillomavirus type 11 E2 protein contains major determinants for nuclear localization and nuclear matrix association. J. Virol. 2000, 74, 3761–3770. [Google Scholar] [CrossRef] [Green Version]
- Nagasaka, K.; Kawana, K.; Osuga, Y.; Fujii, T. PDZ domains and viral infection: Versatile potentials of HPV-PDZ interactions in relation to malignancy. Biomed. Res. Int. 2013, 2013, 369712. [Google Scholar] [CrossRef] [Green Version]
- Huber, B.; Wang, J.W.; Roden, R.B.S.; Kirnbauer, R. RG1-VLP and Other L2-Based, Broad-Spectrum HPV Vaccine Candidates. J. Clin. Med. 2021, 10, 1044. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Primer ID | Application | Primer Sequences (5′-3′) |
|---|---|---|
| Screening WF | First round | AGGCCTCCATTACCTCTTCTG |
| Screening WR | AGCACAGATGGTTCAGTTGT | |
| Screening NF | Second round | AGAGTTTGATCTGCGCCATTTGAT |
| Screening NR | AGATGATGGCGAGCTGG | |
| Gap1 WF | First round | TTGCGCGCAAAAATGAGAGG |
| Gap1 WR | GAATTCCCCTGCTCCAGCTC | |
| Gap1 NF | Second round | TTGCGCGCAAAAATGAGAGG |
| Gap1 NR | GCACACCGTGGACAAAGAAG | |
| Gap2 WF | First round | AATCTGTCCCGTCTCCTGGA |
| Gap2 WR | TGGAACCCCGAATGTGGATG | |
| Gap2 NF | Second round | AATCTGTCCCGTCTCCTGGA |
| Gap2 NR | TGCCAGCAGCTTTACAATGC | |
| Gap3 WF | First round | TGGTCCACCAGATACCGGAA |
| Gap3 WR | TGGAAGTAGGAGTGGGCTGT | |
| Gap3 NF | Second round | GCAAAACACAAACATCCACAGC |
| Gap3 NR | TGGAAGTAGGAGTGGGCTGT | |
| Gap4 WF | First round | AGCTCAGCGCATGTAGTTTTG |
| Gap4 WR | ACCCGAAACTGAGAACCTGAC | |
| Gap4 NF | Second round | CTCAGCGCATGTAGTTTTGCA |
| Gap4 NR | CCACTGGTTTTTGGGGAGGT | |
| Gap5 WF | First round | TGGCAAACGAGAACAAGGTT |
| Gap5 WR | TCCTCCTCCTCTGCTTCAGT | |
| Gap5 NF | Second round | TGCCAGACATTACTTTGCCAG |
| Gap5 NR | TGCAAGCTTTCATTCCGACA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, H.; Ling, Y.; Wang, X.; Wang, H.; Zhu, N.; Li, Y.; Xu, H. Viral Metagenomics Reveals a Putative Novel HPV Type in Anogenital Wart Tissues. Pathogens 2022, 11, 1452. https://doi.org/10.3390/pathogens11121452
Hu H, Ling Y, Wang X, Wang H, Zhu N, Li Y, Xu H. Viral Metagenomics Reveals a Putative Novel HPV Type in Anogenital Wart Tissues. Pathogens. 2022; 11(12):1452. https://doi.org/10.3390/pathogens11121452
Chicago/Turabian StyleHu, Huimin, Yu Ling, Xuan Wang, Hao Wang, Niannan Zhu, Yumei Li, and Hui Xu. 2022. "Viral Metagenomics Reveals a Putative Novel HPV Type in Anogenital Wart Tissues" Pathogens 11, no. 12: 1452. https://doi.org/10.3390/pathogens11121452