
Modeling Effects of Vertebrate Host Exclosures and Host-Targeted Acaricides on Lone Star Tick (Amblyomma americanum, L.) Infestations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
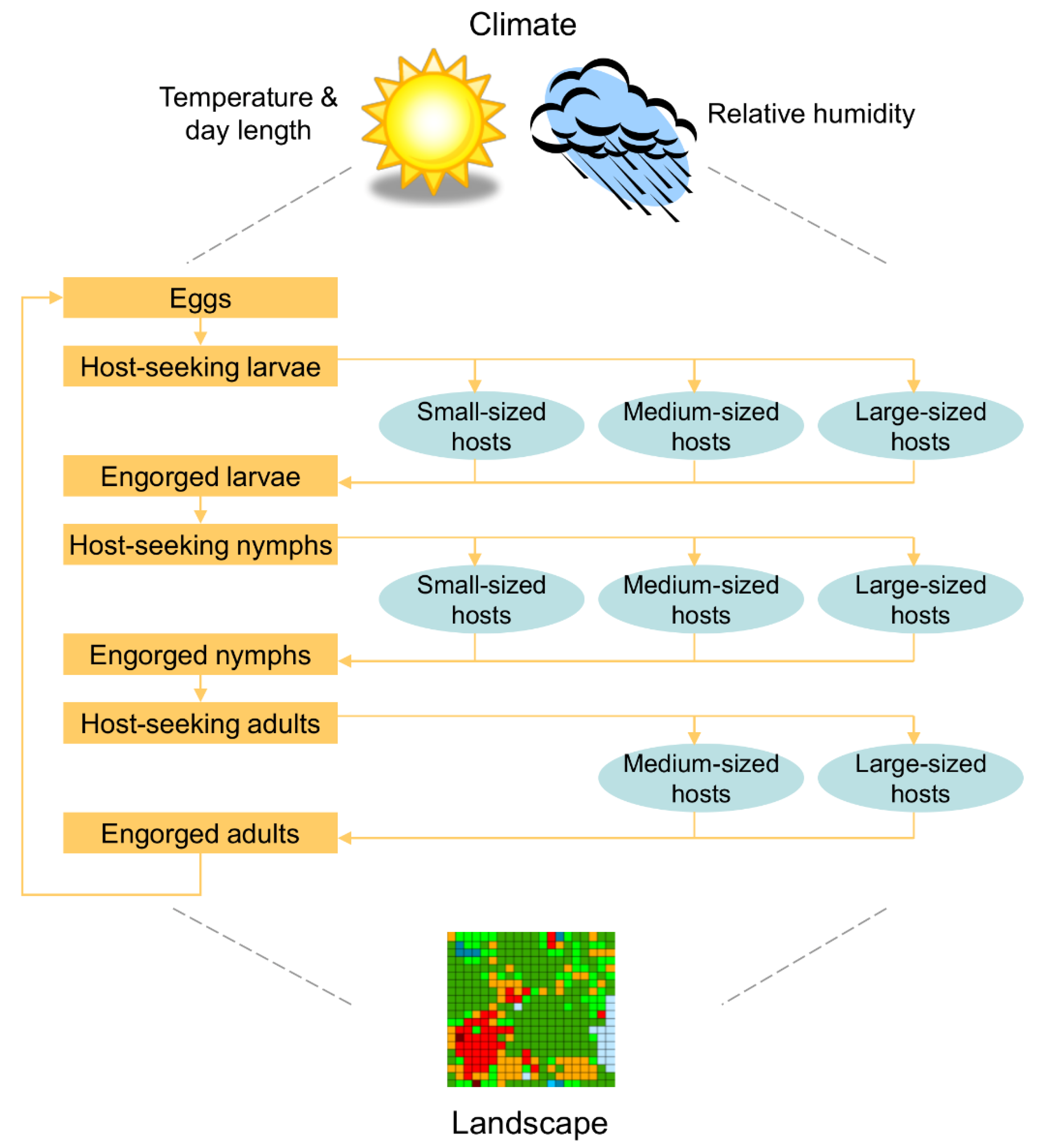
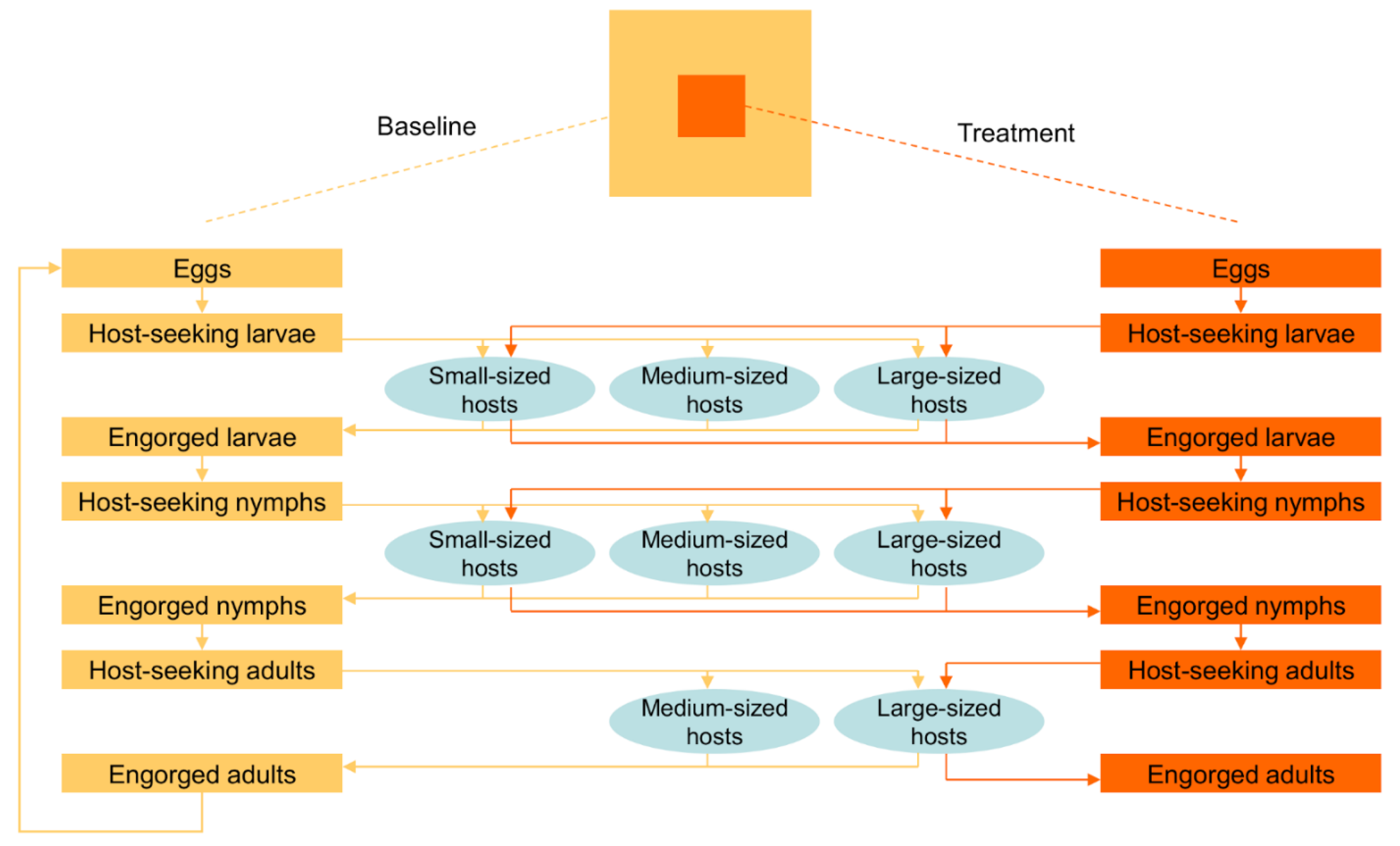
2. Materials and Methods
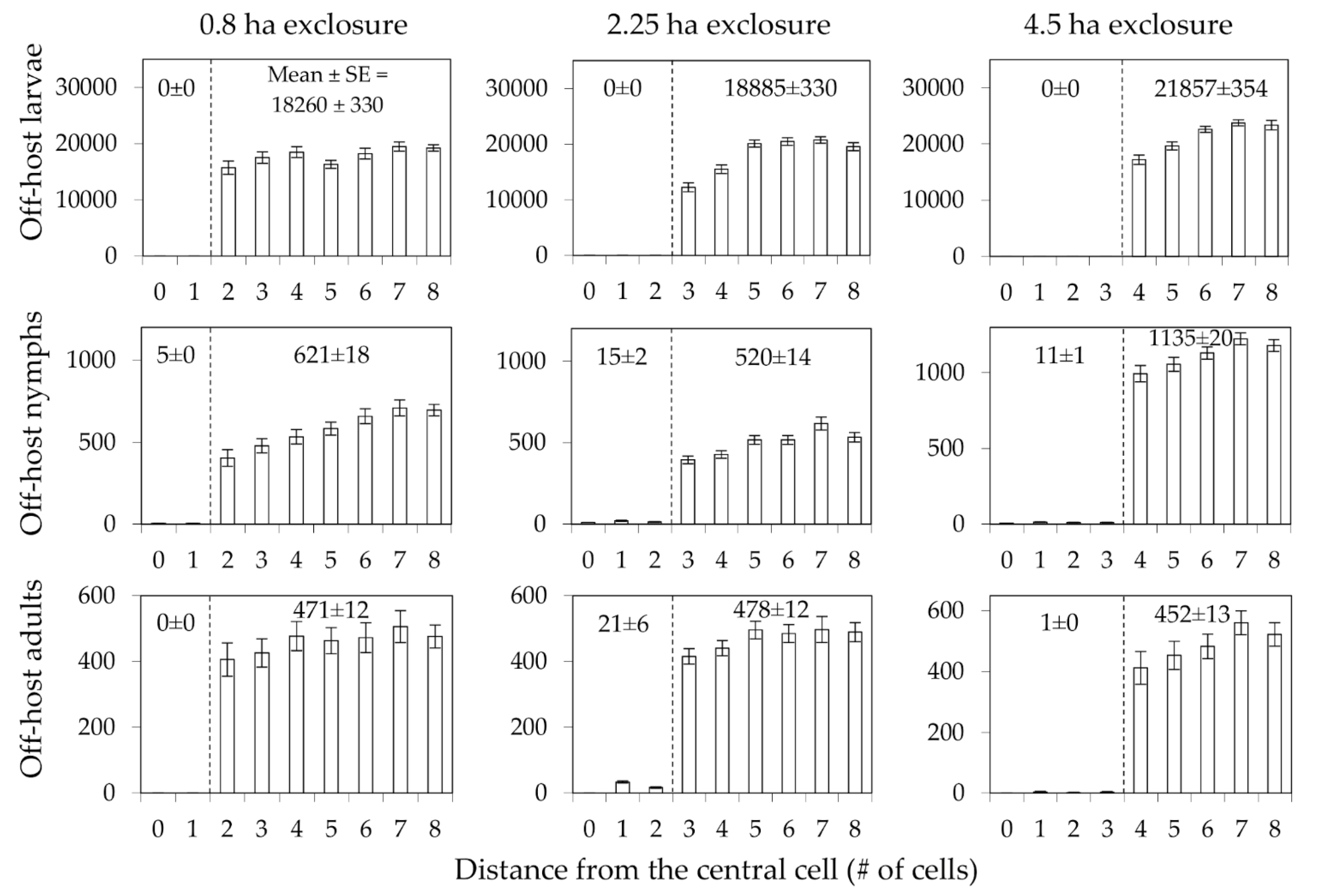
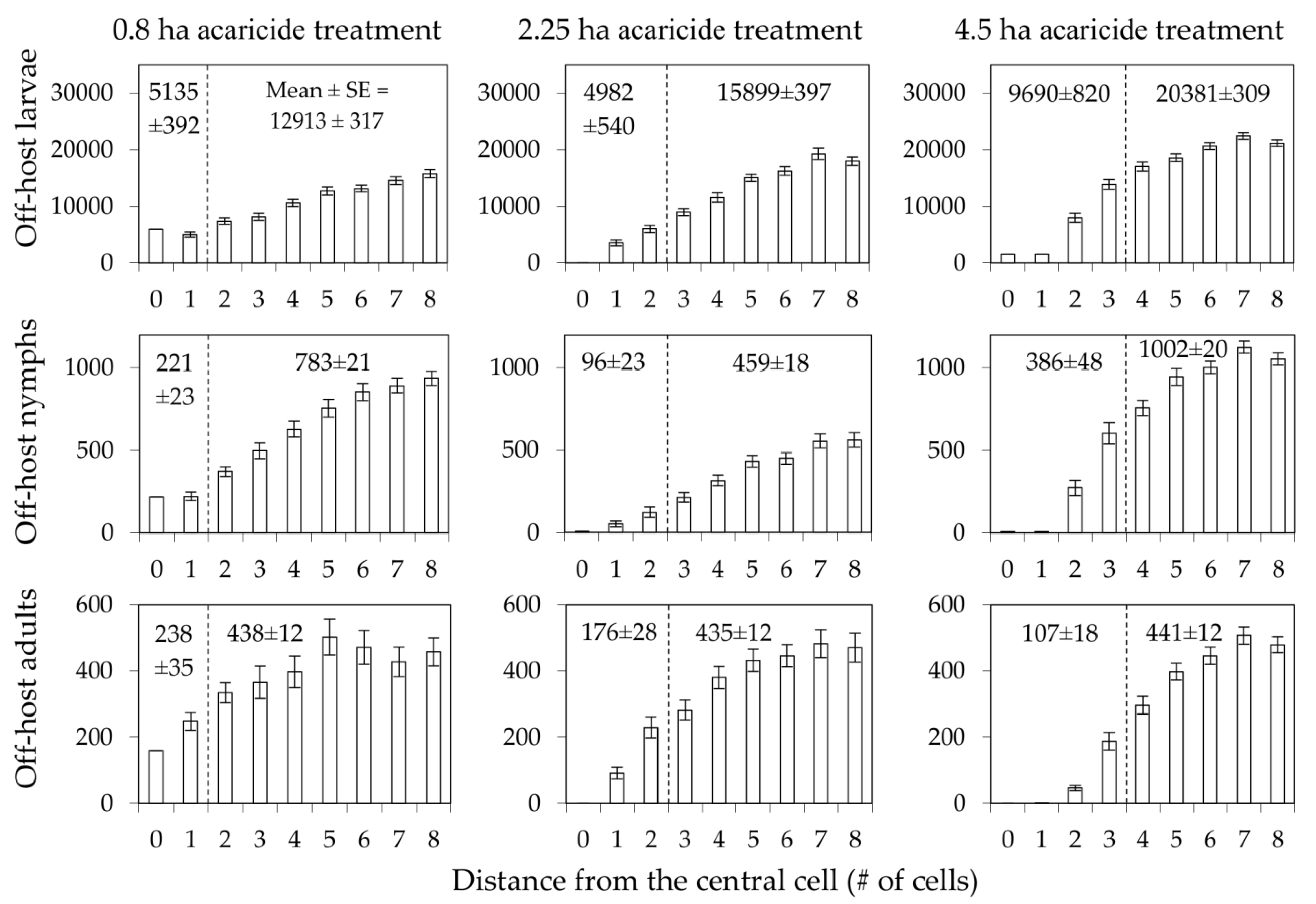
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Childs, J.E.; Paddock, C.D. The ascendancy of Amblyomma americanum as a vector of pathogens affecting humans in the United States. Annu. Rev. Entomol. 2003, 48, 307–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloemer, S.R.; Snoddy, E.L.; Cooney, J.C.; Fairbanks, K. Influence of deer exclusion on populations of lone star ticks and American dog ticks (Acari: Ixodidae). J. Econ. Entomol. 1986, 79, 679–683. [Google Scholar] [CrossRef] [Green Version]
- Perkins, S.E.; Cattadori, I.M.; Tagliapietra, V.; Rizzoli, A.P.; Hudson, P.J. Localized deer absence leads to tick amplification. Ecology 2006, 87, 1981–1986. [Google Scholar] [CrossRef]
- Cooney, J.C.; Burgdorfer, W. Zoonotic potential (Rocky Mountain spotted fever and tularemia) in the Tennessee Valley region: I. Ecologic studies of ticks infesting mammals in land between the lakes. Am. J. Trop. Med. Hyg. 1974, 23, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Bishop, A.; Borski, J.; Wang, H.-H.; Donaldson, T.G.; Michalk, A.; Montgomery, A.; Heldman, S.; Mogg, M.; Derouen, Z.; Grant, W.E. Increasing incidence of spotted fever group rickettsioses in the United States, 2010–2018. Vector-Borne Zoonotic Dis. 2022, 22, 491–497. [Google Scholar] [CrossRef]
- Parker, R.R.; Kohls, G.M. American Q fever: The occurrence of Rickettsia diaporica in Amblyomma ammerucanum in eastern Texas. Public Health Rep. 1943, 58, 1510–1511. [Google Scholar] [CrossRef]
- Schulze, T.A.; Bowen, G.S.; Bosler, E.M.; Lakat, M.F.; Parkin, W.E.; Altman, R.; Ormiston, B.G.; Shisler, J.K. Amblyomma americanum: A potential vector of Lyme disease in New Jersey. Science 1984, 224, 601–603. [Google Scholar] [CrossRef]
- James, A.M.; Liveris, D.; Wormser, G.P.; Schwartz, I.; Montecalvo, M.A.; Johnson, B.J.B. Borrelia lonestari infection after a bite by an Amblyomma americanum tick. J. Infect. Dis. 2001, 183, 1810–1814. [Google Scholar] [CrossRef] [Green Version]
- Loftis, A.D.; Levin, M.L.; Spurlock, J.P. Two USA Ehrlichia spp. cause febrile illness in goats. Vet. Microbiol. 2008, 130, 398–402. [Google Scholar] [CrossRef]
- Mogg, M.; Wang, H.-H.; Baker, A.; Derouen, Z.; Borski, J.; Grant, W.E. Increased incidence of Ehrlichia chaffeensis infections in the United States, 2012 through 2016. Vector-Borne Zoonotic Dis. 2020, 20, 547–550. [Google Scholar] [CrossRef]
- Sonenshine, D.E.; Kocan, K.M.; de la Fuente, J. Tick control: Further thoughts on a research agenda. Trends Parasitol. 2006, 22, 550–551. [Google Scholar] [CrossRef] [PubMed]
- Pound, J.M.; Miller, J.A.; George, J.E.; Fish, D.; Carroll, J.F.; Schulze, T.L.; Daniels, T.J.; Falco, R.C.; StaffordIII, K.C.; Mather, T.N. The United States Department of Agriculture’s Northeast Area-Wide Tick Control Project: Summary and Conclusions. Vector-Borne Zoonotic Dis. 2009, 9, 439–448. [Google Scholar] [CrossRef] [Green Version]
- Piesman, J. Strategies for reducing the risk of Lyme borreliosis in North America. Int. J. Med. Microbiol. 2006, 296, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Stafford, K.C. Reduced abundance of Ixodes scapularis (Acari: Ixodidae) and the tick parasitoid Ixodiphagus hookeri (Hymenoptera: Encyrtidae) with reduction of white-tailed deer. J. Med. Entomol. 2003, 40, 642–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rand, P.W.; Lubelczyk, C.; Holman, M.S.; Lacombe, E.H.; Smith, R.P. Abundance of Ixodes scapularis (Acari: Ixodidae) after the complete removal of deer from an isolated offshore island, endemic for Lyme disease. J. Med. Entomol. 2004, 41, 779–784. [Google Scholar] [CrossRef]
- Wilson, M.L. Distribution and abundance of Ixodes scapularis (Acari: Ixodidae) in North America: Ecological processes and spatial analysis. J. Med. Entomol. 1998, 35, 446–457. [Google Scholar] [CrossRef]
- Stafford, K.C. Reduced abundance of Ixodes scapularis (Acari: Ixodidae) with exclusion of deer by electric fencing. J. Med. Entomol. 1993, 30, 987–996. [Google Scholar] [CrossRef]
- Ginsberg, H.S.; Zhioua, E. Influence of deer abundance on the abundance of questing adult Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 1999, 36, 376–381. [Google Scholar] [CrossRef]
- Ginsberg, H.S.; Butler, M.; Zhioua, E. Effect of deer exclusion by fencing on abundance of Amblyomma americanum (Acari: Ixodidae) on Fire Island, New York, USA. J. Vector Ecol. 2002, 27, 215–221. [Google Scholar]
- Pound, J.M.; Miller, J.A.; George, J.E.; Lemeilleur, C.A. The 4-Poster passive topical treatment device to apply acaricide for controlling ticks (Acari: Ixodidae) feeding on white-tailed deer. J. Med. Entomol. 2000, 37, 588–594. [Google Scholar] [CrossRef]
- White, A.; Gaff, H. Review: Application of tick control technologies for blacklegged, lone star, and american dog ticks. J. Integr. Pest Manag. 2018, 9, 12. [Google Scholar] [CrossRef]
- Hüe, T.; Berger, A.; Wang, H.-H.; Grant, W.E.; Teel, P.D.; de León, A.A.P. Integrated control of the cattle tick, Rhipicephalus australis (Acari: Ixodidae), in New Caledonia through the pasture and cattle management method. Parasitol. Res. 2021, 120, 2749–2758. [Google Scholar] [CrossRef] [PubMed]
- Hüe, T.; Fontfreyde, C.; Wang, H.-H.; Grant, W.E.; Teel, P.D.; de León, A.A.P. Optimizing long-acting acaricide use for integrated tick management of Rhipicephalus australis-infesting cattle in New Caledonia. Trop. Anim. Health Prod. 2021, 53, 384. [Google Scholar] [CrossRef] [PubMed]
- Gaff, H.; Gross, L. Modeling tick-borne disease: A metapopulation model. Bull. Math. Biol. 2007, 69, 265–288. [Google Scholar] [CrossRef]
- Wang, H.H.; Corson, M.S.; Grant, W.E.; Teel, P.D. Quantitative models of Rhipicephalus (Boophilus) ticks: Historical review and synthesis. Ecosphere 2017, 8, e01942. [Google Scholar] [CrossRef]
- Schmidt, K.A.; Ostfeld, R.S. Biodiversity and the dilution effect in disease ecology. Ecology 2001, 82, 609–619. [Google Scholar] [CrossRef]
- Wang, H.-H.; Teel, P.D.; Grant, W.E.; Schuster, G.; de León, A.A.P. Simulated interactions of white-tailed deer (Odocoileus virginianus), climate variation and habitat heterogeneity on southern cattle tick (Rhipicephalus (Boophilus) microplus) eradication methods in south Texas, USA. Ecol. Model. 2016, 342, 82–96. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.-H.; Teel, P.D.; Grant, W.E.; Soltero, F.; Urdaz, J.; Ramírez, A.E.P.; Miller, R.J.; de León, A.A.P. Simulation tools for assessment of tick suppression treatments of Rhipicephalus (Boophilus) microplus on non-lactating dairy cattle in Puerto Rico. Parasites Vectors 2019, 12, 185. [Google Scholar] [CrossRef]
- Wang, H.-H.; Grant, W.E.; Teel, P.D.; Lohmeyer, K.H.; de León, A.P. Enhanced biosurveillance of high-consequence invasive pests: Southern cattle fever ticks, Rhipicephalus (Boophilus) microplus, on livestock and wildlife. Parasites Vectors 2020, 13, 487. [Google Scholar] [CrossRef]
- Agudelo, M.S.; Grant, W.E.; Wang, H.H. Effects of white-tailed deer habitat use preferences on southern cattle fever tick eradication: Simulating impact on “pasture vacation” strategies. Parasites Vectors 2021, 14, 102. [Google Scholar] [CrossRef]
- Wang, H.-H.; Grant, W.E.; Teel, P.D.; Lohmeyer, K.H.; de León, A.A.P. Simulated dynamics of southern cattle fever ticks (Rhipicephalus (Boophilus) microplus) in south Texas, USA: Investigating potential wildlife-mediated impacts on eradication efforts. Parasites Vectors 2021, 14, 231. [Google Scholar] [CrossRef] [PubMed]
- Clymer, B.C.; Howell, D.; Hair, J.A. Animal hosts of economically important ticks (Acarina) in east-central Oklahoma. Ann. Entomol. Soc. Am. 1970, 63, 612–613. [Google Scholar] [CrossRef]
- Wang, H.-H.; Grant, W.E.; Teel, P.D. Simulation of climate-host-parasite-landscape interactions: A spatially explicit model for ticks (Acari: Ixodidae). Ecol. Model. 2012, 243, 42–62. [Google Scholar] [CrossRef]
- Wang, H.H.; Grant, W.; Teel, P.; Hamer, S. Simulation of climate-tick-host-landscape interactions: Effects of shifts in the seasonality of host population fluctuations on tick densities. J. Vector Ecol. 2015, 40, 247–255. [Google Scholar] [CrossRef]
- Wang, H.-H.; Grant, W.; Teel, P.; Hamer, S. Tick-borne infectious agents in nature: Simulated effects of changes in host density on spatial-temporal prevalence of infected ticks. Ecol. Model. 2016, 323, 77–86. [Google Scholar] [CrossRef]
- Goddard, J.; Varela-Stokes, A.S. Role of the lone star tick, Amblyomma americanum (L.), in human and animal diseases. Vet. Parasitol. 2009, 160, 1–12. [Google Scholar] [CrossRef]
- Schmidly, D.J. Texas Mammals East of the Balcones Fault Zone; Texas A&M University: College Station, TX, USA, 1983; p. 400. [Google Scholar]
- Piesman, J.; Eisen, L. Prevention of tick-borne diseases. Annu. Rev. Entomol. 2008, 53, 323–343. [Google Scholar] [CrossRef] [PubMed]
- Drummond, R.O. Seasonal activity of ticks (Acarina: Metastigmata) on cattle in southwestern Texas. Ann. Entomol. Soc. Am. 1967, 60, 439–447. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.-H.; Grant, W.E.; Donaldson, T.G.; Teel, P.D. Modeling Effects of Vertebrate Host Exclosures and Host-Targeted Acaricides on Lone Star Tick (Amblyomma americanum, L.) Infestations. Pathogens 2022, 11, 1412. https://doi.org/10.3390/pathogens11121412
Wang H-H, Grant WE, Donaldson TG, Teel PD. Modeling Effects of Vertebrate Host Exclosures and Host-Targeted Acaricides on Lone Star Tick (Amblyomma americanum, L.) Infestations. Pathogens. 2022; 11(12):1412. https://doi.org/10.3390/pathogens11121412
Chicago/Turabian StyleWang, Hsiao-Hsuan, William E. Grant, Taylor G. Donaldson, and Pete D. Teel. 2022. "Modeling Effects of Vertebrate Host Exclosures and Host-Targeted Acaricides on Lone Star Tick (Amblyomma americanum, L.) Infestations" Pathogens 11, no. 12: 1412. https://doi.org/10.3390/pathogens11121412