
Phytobacter diazotrophicus from Intestine of Caenorhabditis elegans Confers Colonization-Resistance against Bacillus nematocida Using Flagellin (FliC) as an Inhibition Factor

 and
and 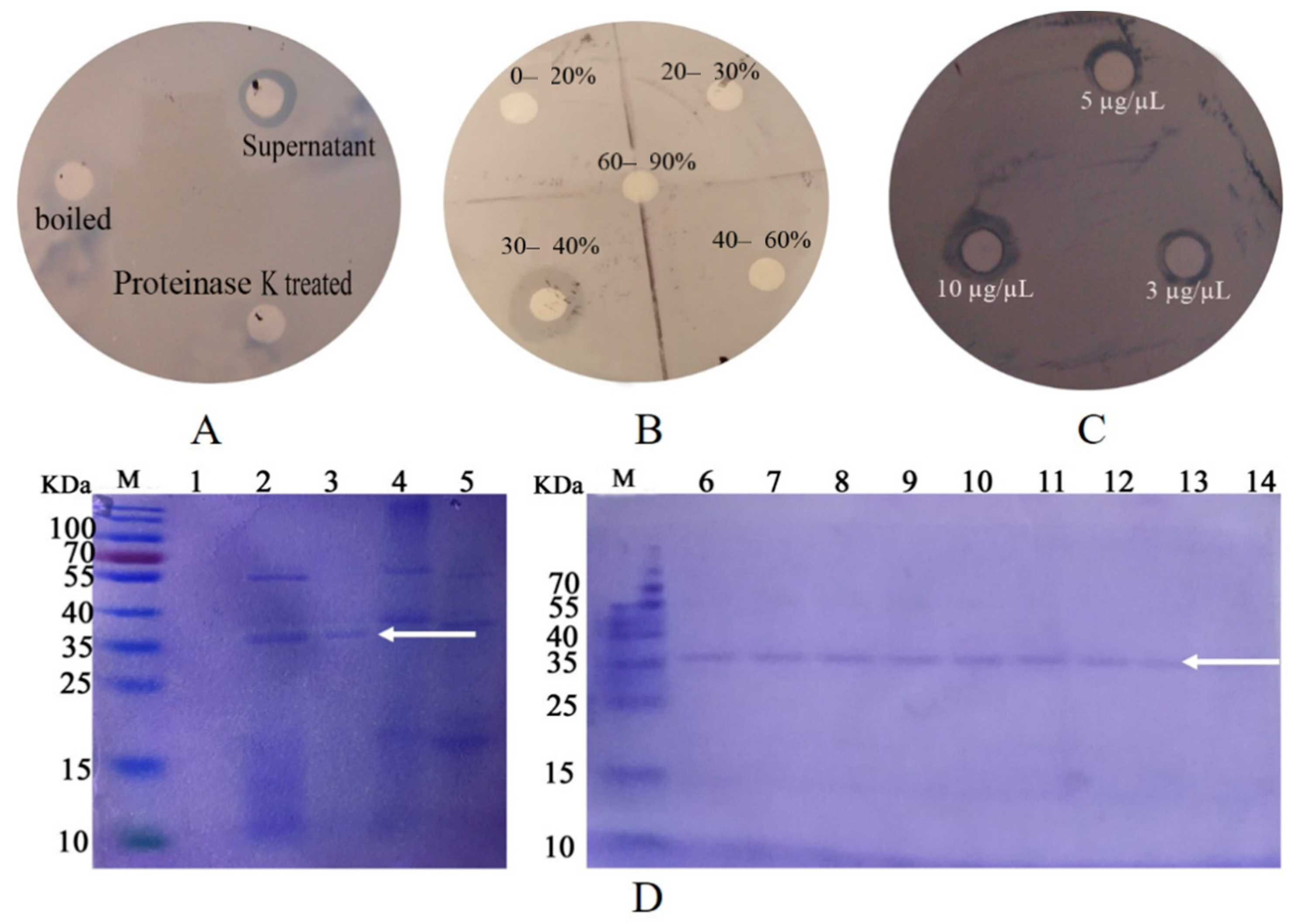{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification Analysis
2.2. Purification of the Inhibition Factor and Identification
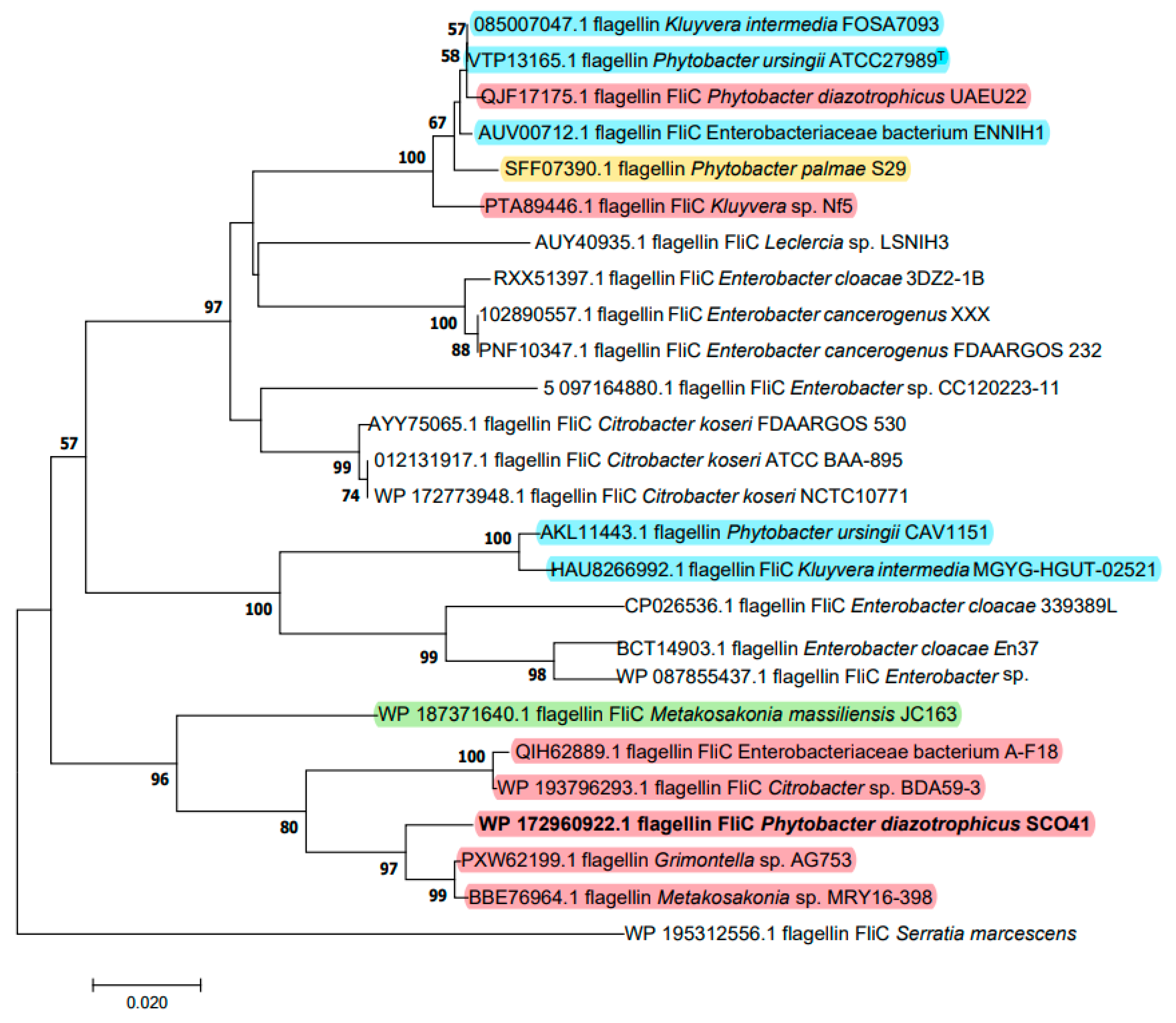
2.3. Cloning of the Gene and Sequence Analysis
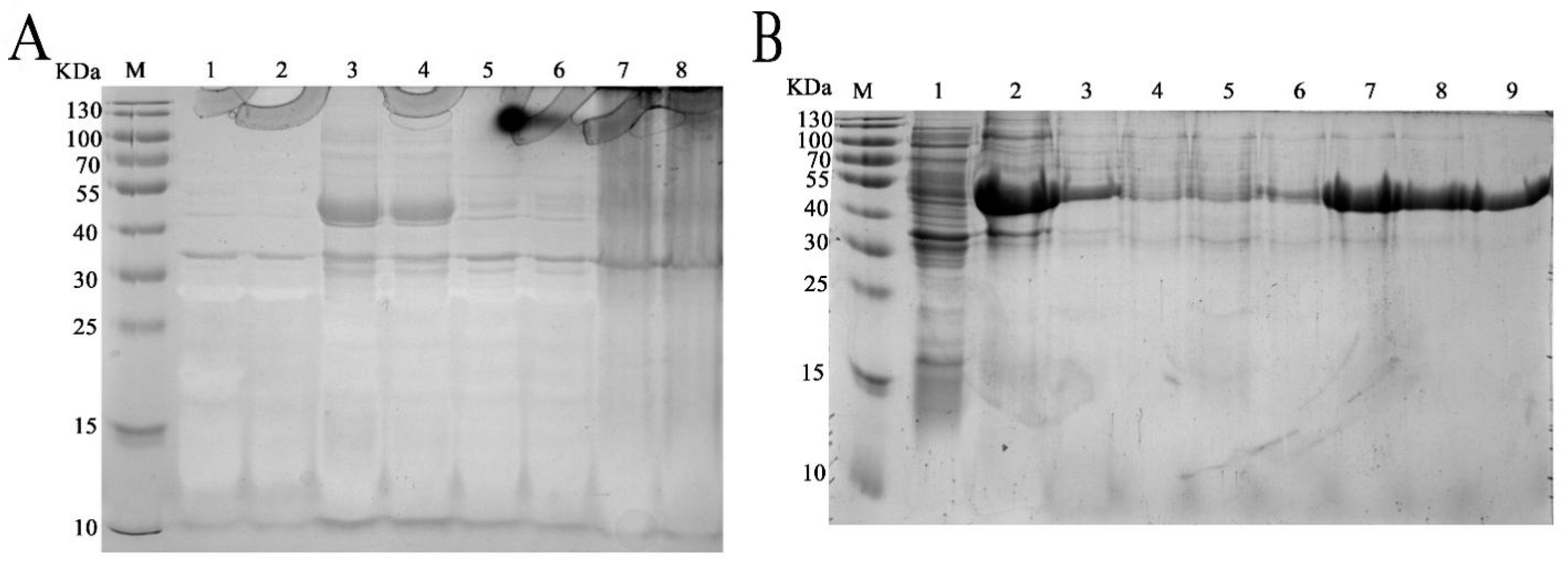
2.4. Heterologous Expression and Purification of rm-FliC
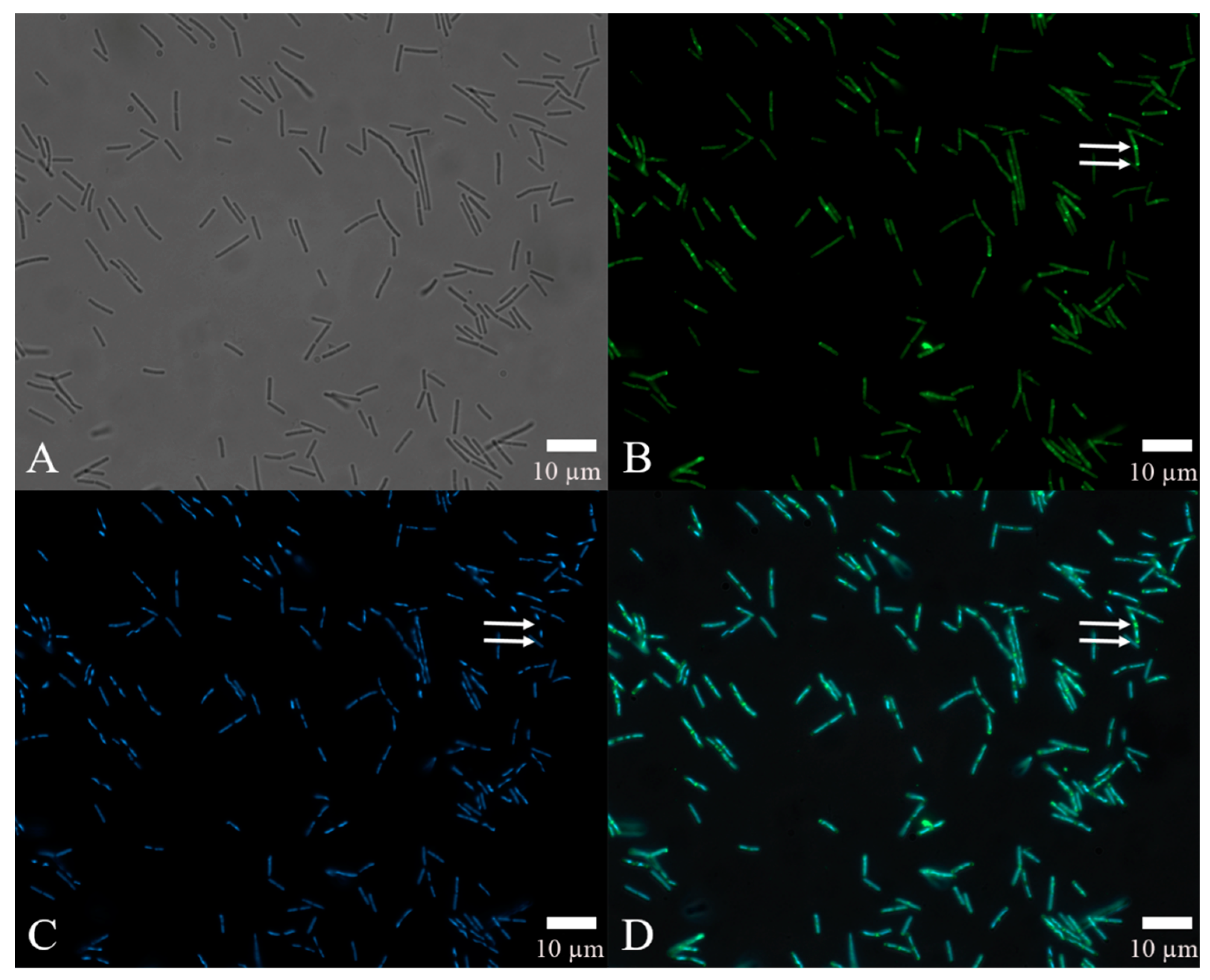
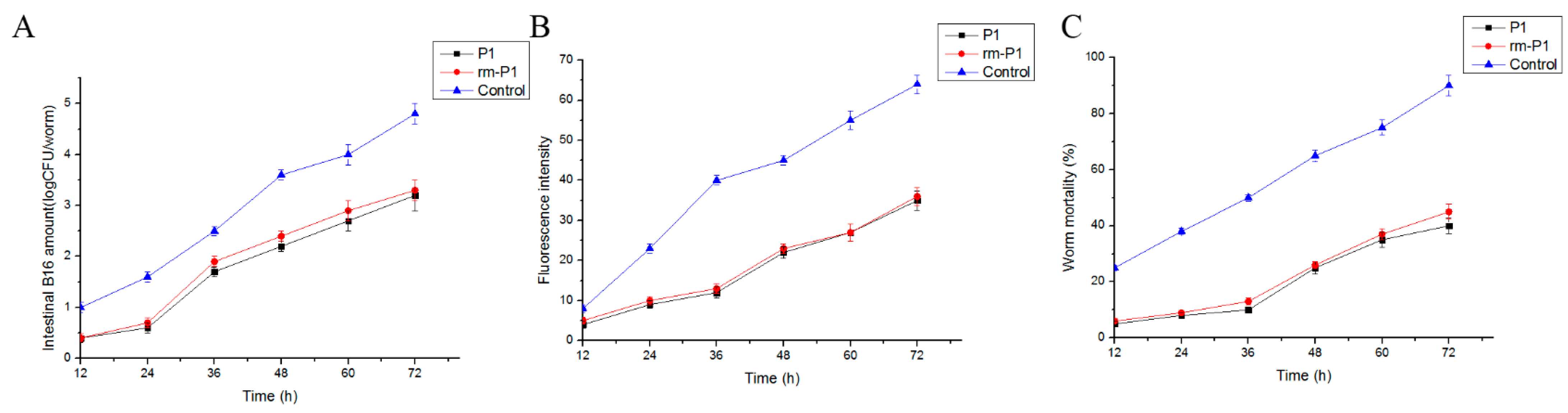
2.5. Bioassay and Localization Results Using rm-FliC Protein
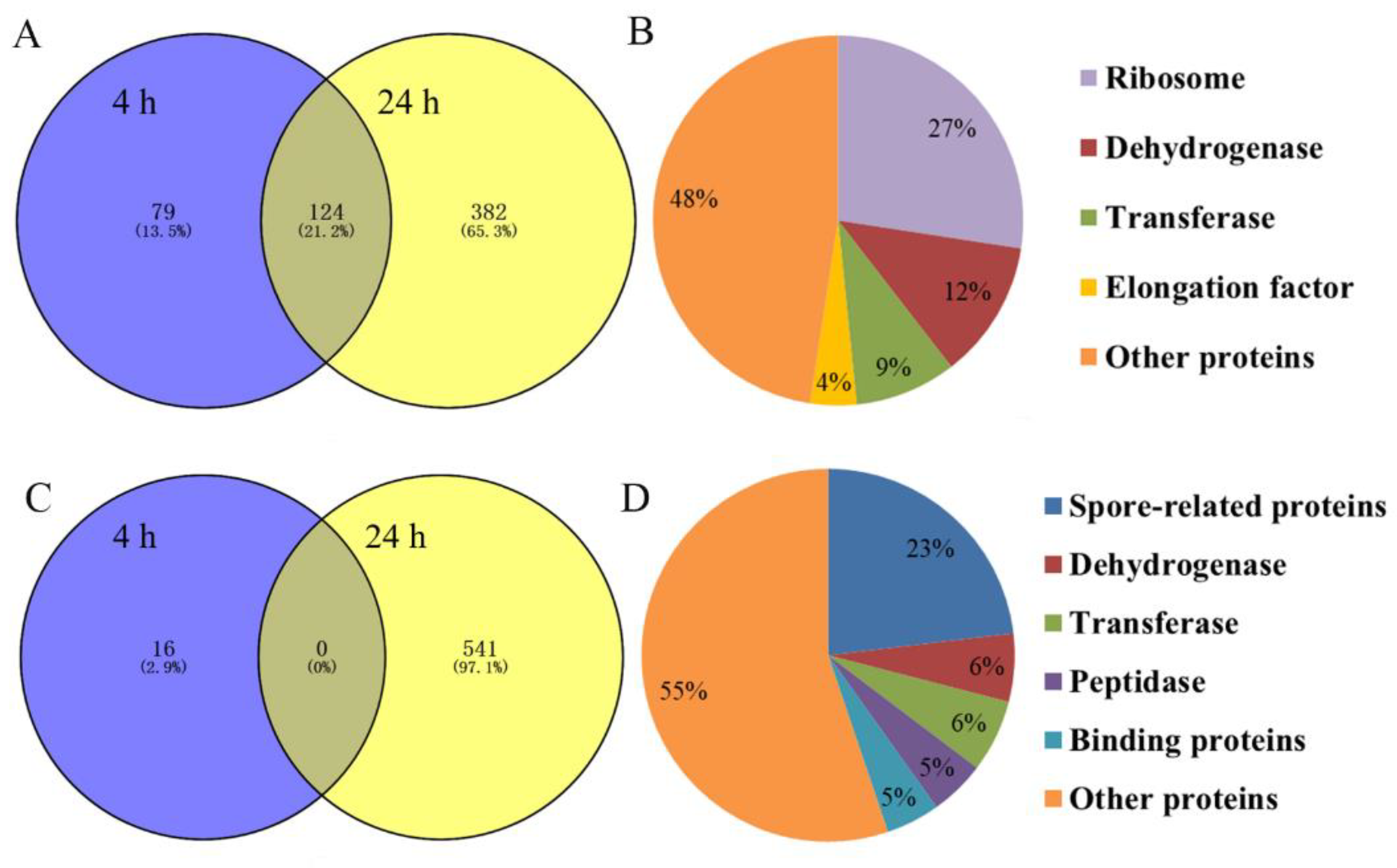
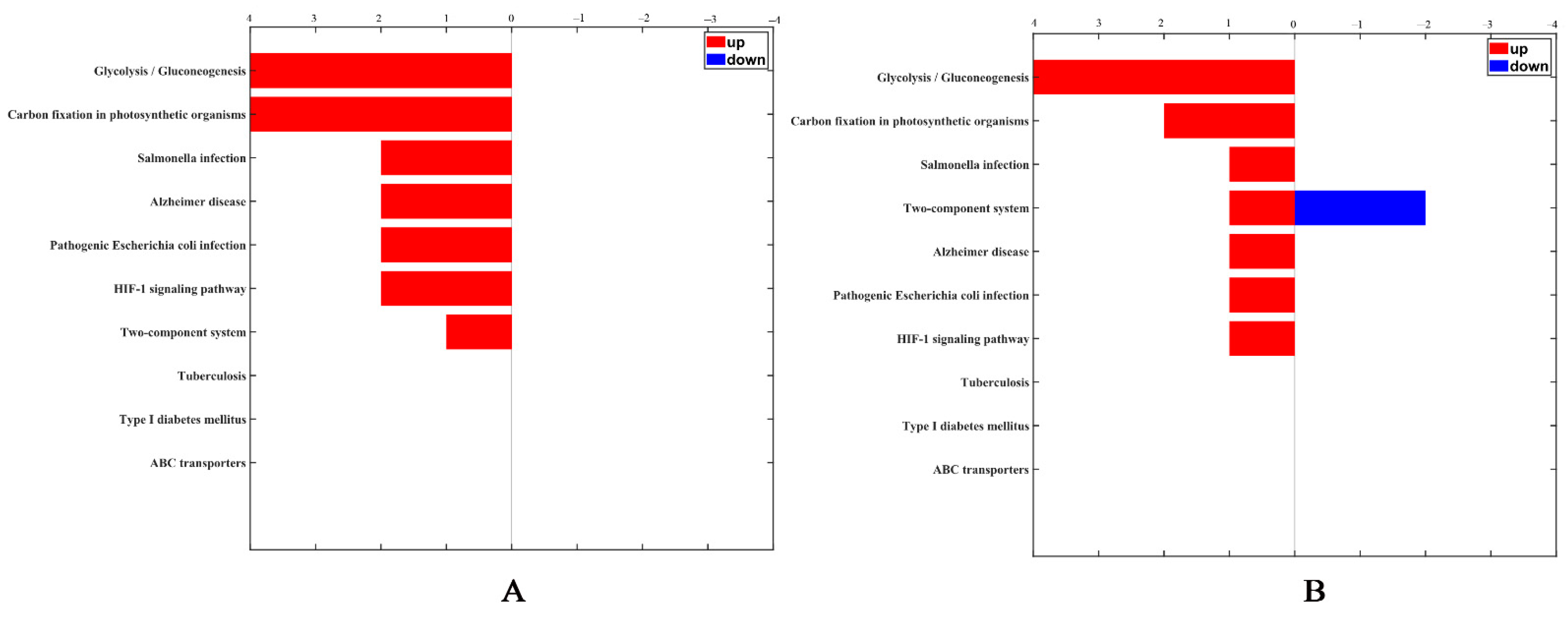
2.6. Identification and Quantitation of Differentially Expressed Proteins after rm-FliC Treatment In Vitro
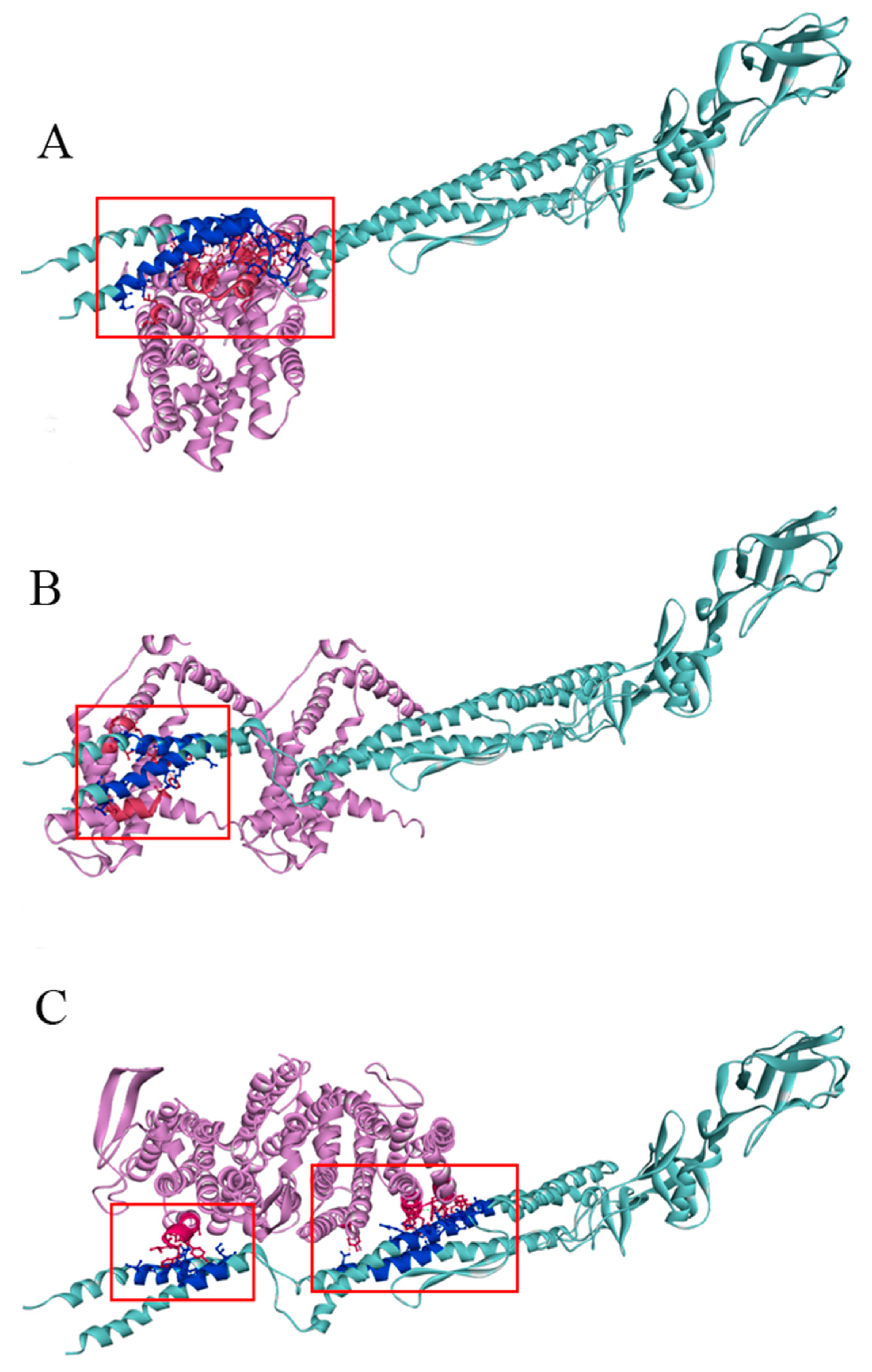
2.7. Molecular Docking Analysis
3. Discussion
4. Methods
4.1. Isolation, Screening and Culture Condition
4.2. Purification of the Inhibition Factor and Mass Spectrometry
4.3. Cloning of the Gene Encoding the Inhibition Factor
4.4. Heterologous Expression and Purification of FliC in P. diazotrophicus SCO41
4.5. Bioassays Using the FliC Protein
4.6. Localization of the FliC Protein in B. nematocida B16
4.7. Proteomic Analysis
4.8. Molecular Docking of FliC
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Schmidt, T.S.B.; Raes, J.; Bork, P. The Human gut microbiome: From association to modulation. Cell 2018, 172, 1198–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felix, M.A.; Duveau, F. Population dynamics and habitat sharing of natural populations of Caenorhabditis elegans and C. briggsae. BMC Biol. 2012, 10, 59. [Google Scholar] [CrossRef] [Green Version]
- Proenca, D.N.; Grass, G.; Morais, P.V. Understanding pine wilt disease: Roles of the pine endophytic bacteria and of the bacteria carried by the disease-causing pinewood nematode. MicrobiologyOpen 2017, 6, e00415. [Google Scholar] [CrossRef] [PubMed]
- Sajnaga, E.; Kazimierczak, W.; Skowronek, M.; Lis, M.; Skrzypek, T.; Wasko, A. Steinernema poinari (Nematoda: Steinernematidae): A new symbiotic host of entomopathogenic bacteria Xenorhabdus bovienii. Arch. Microbiol. 2018, 200, 1307–1316. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.R.; Liu, Q.Z. Evaluation and entomopathogenicity of gut bacteria associated with dauer juveniles of Oscheius chongmingensis (Nematoda: Rhabditidae). MicrobiologyOpen 2019, 8, e00823. [Google Scholar] [CrossRef] [Green Version]
- Mobasseri, M.; Hutchinson, M.C.; Afshar, F.J.; Pedram, M. New evidence of nematode-endosymbiont bacteria coevolution based on one new and one known dagger nematode species of Xiphinema americanum-group (Nematoda, Longidoridae). PLoS ONE 2019, 14, e0217506. [Google Scholar] [CrossRef] [Green Version]
- Cabreiro, F.; Gems, D. Worms need microbes too: Microbiota, health and aging in Caenorhabditis elegans. EMBO Mol. Med. 2013, 5, 1300–1310. [Google Scholar] [CrossRef]
- Montalvo-Katz, S.; Huang, H.; Appel, M.D.; Berg, M.; Shapira, M. Association with soil bacteria enhances p38-dependent infection resistance in Caenorhabditis elegans. Infect. Immun. 2013, 81, 514–520. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, T.; Yasui, C.; Hoshino, K.; Arikawa, K.; Nishikawa, Y. Influence of lactic acid bacteria on longevity of Caenorhabditis elegans and host defense against Salmonella enterica serovar enteritidis. Appl. Environ. Microbiol. 2007, 73, 6404–6409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacNeil, L.T.; Watson, E.; Arda, H.E.; Zhu, L.J.; Walhout, A.J. Diet-induced developmental acceleration independent of TOR and Insulin in C. elegans. Cell 2013, 153, 240–252. [Google Scholar] [CrossRef] [Green Version]
- Iatsenko, I.; Yim, J.J.; Schroeder, F.C.; Sommer, R.J. B. subtilis GS67 protects C. elegans from Gram-Positive pathogens via fengycin-mediated microbial antagonism. Curr. Biol. 2014, 24, 2720–2727. [Google Scholar]
- Oro, V.; Knezevic, M.; Dinic, Z.; Delic, D. Bacterial microbiota isolated from cysts of Globodera rostochiensis (Nematoda: Heteroderidae). Plants 2020, 9, 1146. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Yang, J.; Zhang, K.Q. Bacteria used in the biological control of plant-parasitic nematodes: Populations, mechanisms of action, and future prospects. FEMS Microbiol. Ecol. 2007, 61, 197–213. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zou, C.; Xu, J.; Ji, X.; Niu, X.; Yang, J.; Huang, X.; Zhang, K.Q. Molecular mechanisms of nematode-nematophagous microbe interactions: Basis for biological control of plant-parasitic nematodes. Annu. Rev. Phytopathol. 2015, 53, 67–95. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Wang, Y.; Deng, Y.; Zhang, Y.; Zhang, L.; Niu, Q. Stenotrophomonas strain CPCC 101271, an intestinal lifespan-prolonging bacterium for Caenorhabditis elegans that assists in host resistance to “Bacillus nematocida” colonization. Arch. Microbiol. 2021, 203, 4951–4960. [Google Scholar] [CrossRef] [PubMed]
- Buffie, C.G.; Pamer, E.G. Microbiota-mediated colonization resistance against intestinal pathogens. Nat. Rev. Immunol. 2013, 13, 790–801. [Google Scholar] [CrossRef] [Green Version]
- Suriyanarayanan, T.; Periasamy, S.; Lin, M.H.; Ishihama, Y.; Swarup, S. Flagellin FliC phosphorylation affects type 2 protease secretion and biofilm dispersal in Pseudomonas aeruginosa PAO1. PLoS ONE 2016, 11, e0164155. [Google Scholar] [CrossRef]
- Biedma, M.E.; Cayet, D.; Tabareau, J.; Rossi, A.H.; Ivicak-Kocjan, K.; Moreno, G.; Errea, A.; Soulard, D.; Parisi, G.; Jerala, R.; et al. Recombinant flagellins with deletions in domains D1, D2, and dD3: Characterization as novel immunoadjuvants. Vaccine 2019, 37, 652–663. [Google Scholar] [CrossRef]
- Bruxelle, J.F.; Mizrahi, A.; Hoys, S.; Collignon, A.; Janoir, C.; Pechine, S. Clostridium difficile flagellin FliC: Evaluation as adjuvant and use in a mucosal vaccine against Clostridium difficile. PLoS ONE 2017, 12, e0187212. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.K.; Gupta, A.; Batra, L.; Tuteja, U. Escherichia coli expressed flagellin C (FliC) of Salmonella Typhi improved the protective efficacy of YopE against plague infection. Vaccine 2019, 37, 19–24. [Google Scholar] [CrossRef]
- Zhao, Y.; Chang, J.; Zhang, B.; Tong, P.; Wang, C.; Ran, D.; Su, Y. TLR-5 agonist Salmonella abortus equi flagellin FliC enhances FliC-gD-based DNA vaccination against equine herpesvirus 1 infection. Arch. Virol. 2019, 164, 1371–1382. [Google Scholar] [CrossRef]
- Hajam, I.A.; Dar, P.A.; Shahnawaz, I.; Jaume, J.C.; Lee, J.H. Bacterial flagellin-a potent immunomodulatory agent. Exp. Mol. Med. 2017, 49, e373. [Google Scholar] [CrossRef]
- Dai, F.; Li, Y.; Shao, Y.; Li, C.; Zhang, W. FliC of Vibrio splendidus-related strain involved in adhesion to Apostichopus japonicus. Microb. Pathog. 2020, 149, 104503. [Google Scholar] [CrossRef]
- Bao, Z.; Wei, H.L.; Ma, X.; Swingle, B. Pseudomonas syringae AlgU downregulates flagellin gene expression, helping evade plant immunity. J. Bacteriol. 2020, 202, e00418–e00419. [Google Scholar] [CrossRef]
- Niu, Q.; Zhang, L.; Zhang, K.; Huang, X.; Hui, F.; Kan, Y.; Yao, L. Changes in intestinal microflora of Caenorhabditis elegans following Bacillus nematocida B16 infection. Sci. Rep. 2016, 6, 20178. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Huang, B.; Chen, J.; Li, W.; Yang, L.; Yao, L.; Niu, Q. Whole-genome analysis of the colonization-resistant bacterium Phytobacter sp. SCO41T isolated from Bacillus nematocida B16-fed adult Caenorhabditis elegans. Mol. Biol. Rep. 2019, 46, 1563–1575. [Google Scholar] [CrossRef]
- Ma, Y.; Yao, R.; Li, Y.; Wu, X.; Li, S.; An, Q. Proposal for unification of the genus Metakosakonia and the genus Phytobacter to a single genus Phytobacter and reclassification of Metakosakonia massiliensis as Phytobacter massiliensis comb. nov. Curr. Microbiol. 2020, 77, 1945–1954. [Google Scholar] [CrossRef]
- Pillonetto, M.; Arend, L.N.; Faoro, H.; D’Espindula, H.R.S.; Blom, J.; Smits, T.H.M.; Mira, M.T.; Rezzonico, F. Emended description of the genus Phytobacter, its type species Phytobacter diazotrophicus (zhang 2008) and description of Phytobacter ursingii sp. nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 176–184. [Google Scholar] [CrossRef]
- Zhang, G.X.; Peng, G.X.; Wang, E.T.; Yan, H.; Yuan, Q.H.; Zhang, W.; Lou, X.; Wu, H.; Tan, Z.Y. Diverse endophytic nitrogen-fixing bacteria isolated from wild rice Oryza rufipogon and description of Phytobacter diazotrophicus gen. nov. sp. nov. Arch. Microbiol. 2008, 189, 431–439. [Google Scholar] [CrossRef]
- Medina-Cordoba, L.K.; Chande, A.T.; Rishishwar, L.; Mayer, L.W.; Valderrama-Aguirre, L.C.; Valderrama-Aguirre, A.; Gaby, J.C.; Kostka, J.E.; Jordan, I.K. Genomic characterization and computational phenotyping of nitrogen-fixing bacteria isolated from Colombian sugarcane fields. Sci. Rep. 2021, 11, 9187. [Google Scholar] [CrossRef]
- Salha, Y.; Sudalaimuthuasari, N.; Kundu, B.; AlMaskari, R.S.; Alkaabi, A.S.; Hazzouri, K.M.; AbuQamar, S.F.; El-Tarabily, K.A.; Amiri, K.M.A. Complete Genome Sequence of Phytobacter diazotrophicus strain UAEU22, a plant growth-promoting bacterium isolated from the date palm rhizosphere. Microbiol. Resour. Announc. 2020, 9, e00499-20. [Google Scholar] [CrossRef]
- Benslama, O.; Boulahrouf, A. High-quality draft genome sequence of Enterobacter sp. Bisph2, a glyphosate-degrading bacterium isolated from a sandy soil of Biskra, Algeria. Genom. Data 2016, 8, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Sekizuka, T.; Matsui, M.; Takahashi, T.; Hayashi, M.; Suzuki, S.; Tokaji, A.; Kuroda, M. Complete genome sequence of bla IMP-6-positive Metakosakonia sp. MRY16-398 isolate from the ascites of a diverticulitis patient. Front. Microbiol. 2018, 9, 2853. [Google Scholar] [CrossRef] [Green Version]
- Tetz, G.; Tetz, V. Draft genome sequence of Kluyvera intestini strain GT-16 isolated from the stomach of a patient with gastric cancer. Genome Announc. 2016, 4, e01432-16. [Google Scholar] [CrossRef] [Green Version]
- Hughes, K.T.; Aldridge, P.D. Putting a lid on it. Nat. Struct. Biol. 2001, 8, 96–97. [Google Scholar] [CrossRef]
- Gomez-Gomez, L.; Boller, T. Flagellin perception: A paradigm for innate immunity. Trends Plant Sci. 2002, 7, 251–256. [Google Scholar] [CrossRef]
- Santos, P.G.; Santos, P.A.; Bello, A.R.; Freitas-Almeida, A.C. Association of Aeromonas caviae polar and lateral flagella with biofilm formation. Lett. Appl. Microbiol. 2011, 52, 49–55. [Google Scholar] [CrossRef]
- Shimoyama, T.; Kato, S.; Ishii, S.; Watanabe, K. Flagellum mediates symbiosis. Science 2009, 323, 1574. [Google Scholar] [CrossRef]
- Nezametdinova, V.Z.; Zakharevich, N.V.; Alekseeva, M.G.; Averina, O.V.; Mavletova, D.A.; Danilenko, V.N. Identification and characterization of the serine/threonine protein kinases in Bifidobacterium. Arch. Microbiol. 2014, 196, 125–136. [Google Scholar] [CrossRef]
- Kang, C.M.; Abbott, D.W.; Park, S.T.; Dascher, C.C.; Cantley, L.C.; Husson, R.N. The Mycobacterium tuberculosis serine/threonine kinases PknA and PknB: Substrate identification and regulation of cell shape. Genes Dev. 2005, 19, 1692–1704. [Google Scholar] [CrossRef] [Green Version]
- Jensen, H.M.; Eng, T.; Chubukov, V.; Herbert, R.A.; Mukhopadhyay, A. Improving membrane protein expression and function using genomic edits. Sci. Rep. 2017, 7, 13030. [Google Scholar] [CrossRef]
- Kumazaki, K.; Kishimoto, T.; Furukawa, A.; Mori, H.; Tanaka, Y.; Dohmae, N.; Ishitani, R.; Tsukazaki, T.; Nureki, O. Crystal structure of Escherichia coli YidC, a membrane protein chaperone and insertase. Sci. Rep. 2014, 4, 7299. [Google Scholar] [CrossRef] [Green Version]
- Borisova, V.B.; Gennis, R.B.; Hemp, J.; Verkhovskyc, M.I. The cytochrome bd respiratory oxygen reductases. Biochim. Biophys. Acta (BBA)-Bioenergy 2011, 1807, 1398–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endley, S.; McMurray, D.; Ficht, T.A. Interruption of the cydB locus in Brucella abortus attenuates intracellular survival and virulence in the mouse model of infection. J. Bacteriol. 2001, 183, 2454–2462. [Google Scholar] [CrossRef] [Green Version]
- Gray, N. Ecology of nematophagous fungi: Comparison of the soil sprinkling method with the Baerman funnel technique in the isolation of endoparasites. Soil Biol. Biochem. 1984, 16, 81–83. [Google Scholar] [CrossRef]
- Petersen, C.; Dirksen, P.; Prahl, S.; Strathmann, E.A.; Schulenburg, H. The prevalence of Caenorhabditis elegans across 1.5 years in selected North German locations: The importance of substrate type, abiotic parameters, and Caenorhabditis competitors. BMC Ecol. 2014, 14, 4. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.W.; Niu, Q.H.; Zhou, W.; Zhang, K.Q. Bacillus nematocida sp. nov., a novel bacterial strain with nematotoxic activity isolated from soil in Yunnan, China. Syst. Appl. Microbiol. 2005, 28, 323–327. [Google Scholar] [CrossRef]
- Niu, Q.H.; Huang, X.; Hui, F.; Huang, S.; Ke, T.; Zhang, K.Q.; Zhang, L. Colonization of Caenorhabditis elegans by Bacillus nematocida B16, a bacterial opportunistic pathogen. J. Mol. Microbiol. Biotechnol. 2012, 22, 258–267. [Google Scholar] [CrossRef]
- Niu, Q.; Zheng, H.; Zhang, L.; Qin, F.; Facemire, L.; Zhang, G.; Cao, F.; Zhang, K.Q.; Huang, X.; Yang, J.; et al. Knockout of the adp gene related with colonization in Bacillus nematocida B16 using customized transcription activator-like effectors nucleases. Microb. Biotechnol. 2015, 8, 681–692. [Google Scholar] [CrossRef]
- Aballay, A.; Yorgey, P.; Ausubel, F.M. Salmonella typhimurium proliferates and establishes a persistent infection in the intestine of Caenorhabditis elegans. Curr. Biol. 2000, 10, 1539–1542. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, Q.; Liu, S.; Yin, M.; Lei, S.; Rezzonico, F.; Zhang, L. Phytobacter diazotrophicus from Intestine of Caenorhabditis elegans Confers Colonization-Resistance against Bacillus nematocida Using Flagellin (FliC) as an Inhibition Factor. Pathogens 2022, 11, 82. https://doi.org/10.3390/pathogens11010082
Niu Q, Liu S, Yin M, Lei S, Rezzonico F, Zhang L. Phytobacter diazotrophicus from Intestine of Caenorhabditis elegans Confers Colonization-Resistance against Bacillus nematocida Using Flagellin (FliC) as an Inhibition Factor. Pathogens. 2022; 11(1):82. https://doi.org/10.3390/pathogens11010082
Chicago/Turabian StyleNiu, Qiuhong, Suyao Liu, Mingshen Yin, Shengwei Lei, Fabio Rezzonico, and Lin Zhang. 2022. "Phytobacter diazotrophicus from Intestine of Caenorhabditis elegans Confers Colonization-Resistance against Bacillus nematocida Using Flagellin (FliC) as an Inhibition Factor" Pathogens 11, no. 1: 82. https://doi.org/10.3390/pathogens11010082