
Current Understanding of the Pathogenesis of Porcine Circovirus 3


Abstract
:1. Introduction
2. Pathogenesis of Certain PCV3 Associated Diseases
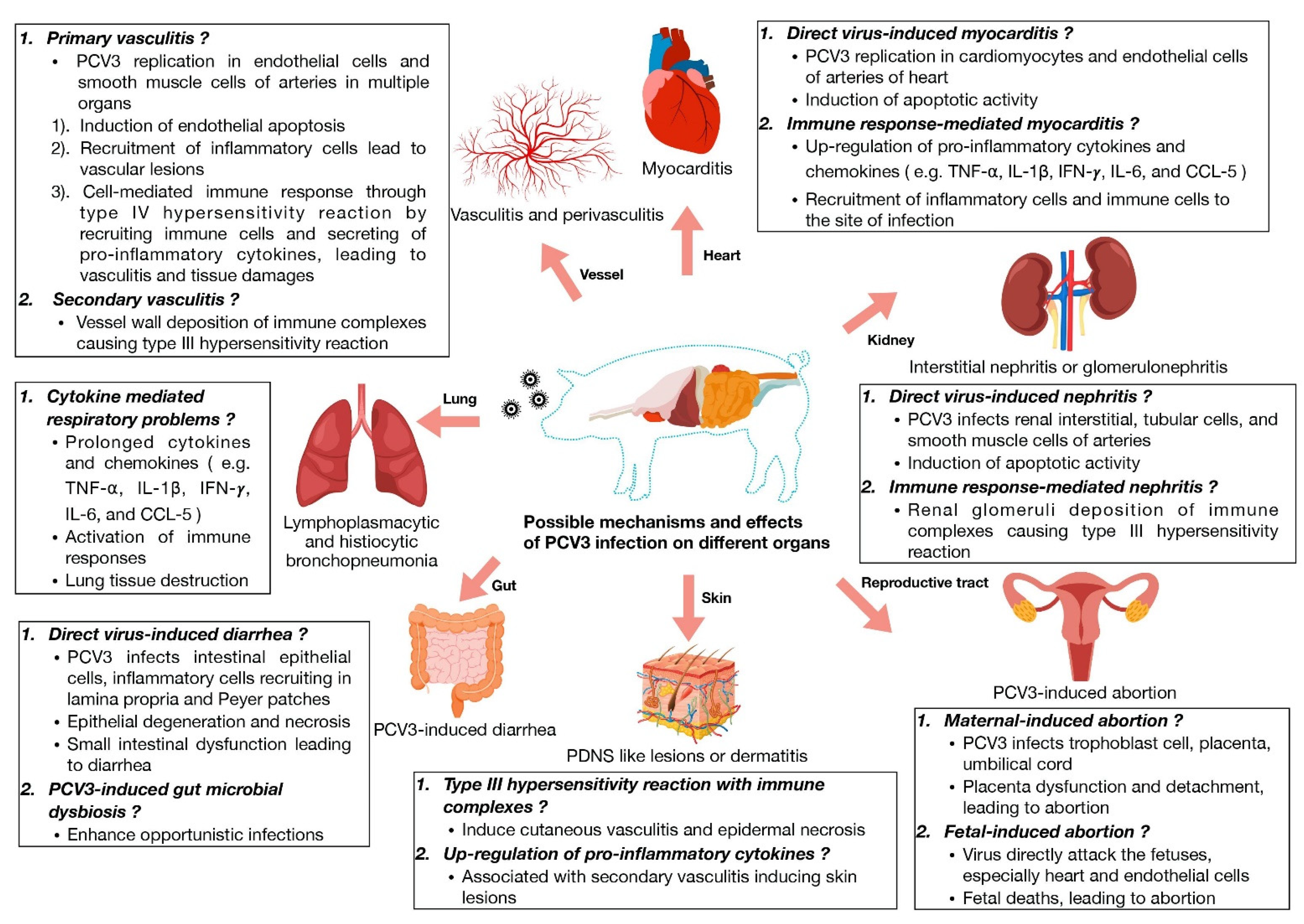
2.1. Myocarditis, Nephritis and Vasculitis
2.2. Porcine Dermatitis and Nephropathy Syndrome (PDNS)
2.3. Reproductive Failure
2.4. Respiratory Disease and Diarrhea
3. What Have We Learnt from the Recent Experimental Infection Studies?
3.1. Clinical Signs and Pathological Changes from the Experimental Infection Studies
3.2. PCV3 Infection Kinetics and Dynamics in the Experimental Infection Studies
3.3. Antibody Responses following PCV3 Infection in the Experimental Infection Studies
4. Possible Factors Contributing to the Varied Outcomes between the Experimental Infection Studies
4.1. Virus Strain
4.2. Virus Infectivity
4.3. Coinfection
4.4. Route of Infection
4.5. Immune Stimulation
4.6. Pig Breed
4.7. Microbial Colonization
5. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harding, J.C. The clinical expression and emergence of porcine circovirus 2. Vet. Microbiol. 2003, 98, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Harding, J.C. Postweaning multisystemic wasting syndrome: Epidemiology and clinical presentation. J. Swine Health Prod. 1998, 6, 249–254. [Google Scholar]
- West, K.H.; Bystrom, J.M.; Wojnarowicz, C.; Shantz, N.; Jacobson, M.; Allan, G.M.; Haines, D.M.; Clark, E.G.; Krakowka, S.; McNeilly, F.; et al. Myocarditis and abortion associated with intrauterine infection of sows with porcine circovirus 2. J. Vet. Diagn. Investig. 1999, 11, 530–532. [Google Scholar] [CrossRef] [Green Version]
- Rosell, C.; Segalés, J.; Ramos-Vara, J.A.; Folch, J.M.; Rodriguez-Arrioja, G.M.; Duran, C.O.; Balasch, M.; Plana-Durán, J.; Domingo, M. Identification of porcine circovirus in tissues of pigs with porcine dermatitis and nephropathy syndrome. Vet. Rec. 2000, 146, 40–43. [Google Scholar] [CrossRef]
- Kim, J.; Chung, H.-K.; Chae, C. Association of porcine circovirus 2 with porcine respiratory disease complex. Vet. J. 2003, 166, 251–256. [Google Scholar] [CrossRef]
- Phan, T.; Giannitti, F.; Rossow, S.; Marthaler, D.; Knutson, T.P.; Li, L.; Deng, X.; Resende, T.; Vannucci, F.; Delwart, E. Detection of a novel circovirus PCV3 in pigs with cardiac and multi-systemic inflammation. Virol. J. 2016, 13, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palinski, R.; Piñeyro, P.; Shang, P.; Yuan, F.; Guo, R.; Fang, Y.; Byers, E.; Hause, B.M. A Novel Porcine Circovirus Distantly Related to Known Circoviruses Is Associated with Porcine Dermatitis and Nephropathy Syndrome and Reproductive Failure. J. Virol. 2016, 91, e01879-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.H.; Hu, W.Q.; Li, J.Y.; Liu, T.N.; Zhou, J.Y.; Opriessnig, T.; Xiao, C.T. Novel circovirus species identified in farmed pigs designated as Porcine circovirus 4, Hunan province, China. Transbound. Emerg. Dis. 2019, 67, 1057–1061. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.; Do, H.; Huynh, T.; Park, Y.; Park, B.; Chung, H. Molecular-based detection, genetic characterization and phylogenetic analysis of porcine circovirus 4 from Korean domestic swine farms. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef]
- Zhai, S.-L.; Zhou, X.; Zhang, H.; Hause, B.M.; Lin, T.; Liu, R.; Chen, Q.-L.; Wei, W.-K.; Lv, D.-H.; Wen, X.-H.; et al. Comparative epidemiology of porcine circovirus type 3 in pigs with different clinical presentations. Virol. J. 2017, 14, 222. [Google Scholar] [CrossRef] [Green Version]
- Kedkovid, R.; Woonwong, Y.; Arunorat, J.; Sirisereewan, C.; Sangpratum, N.; Lumyai, M.; Kesdangsakonwut, S.; Teankum, K.; Jittimanee, S.; Thanawongnuwech, R. Porcine circovirus type 3 (PCV3) infection in grower pigs from a Thai farm suffering from porcine respiratory disease complex (PRDC). Vet. Microbiol. 2018, 215, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Park, J.-Y.; Jung, J.-Y.; Kim, H.-Y.; Park, Y.-R.; Lee, K.-K.; Lyoo, Y.S.; Yeo, S.-G.; Park, C.-K. Detection and genetic characterization of porcine circovirus 3 from aborted fetuses and pigs with respiratory disease in Korea. J. Vet. Sci. 2018, 19, 721–724. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Yuan, W.; Li, Z.; Zhang, Y.; Zeng, X.; Zhao, M.; Ye, Y.; Ding, Z.; He, H.; Wu, Q.; et al. Porcine Circovirus Type 3 in Pig Farms Experiencing Diarrhea in Jiangxi, China: Prevalence, Genome Sequence and Pathogenicity. Animals 2020, 10, 2324. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.H.; Mai, K.J.; Zhou, L.; Wu, R.T.; Tang, X.Y.; Wu, J.L.; He, L.L.; Lan, T.; Xie, Q.M.; Sun, Y.; et al. Detection and genome sequencing of porcine circovirus 3 in neonatal pigs with congenital tremors in South China. Transbound. Emerg. Dis. 2017, 64, 1650–1654. [Google Scholar] [CrossRef] [PubMed]
- Arruda, B.; Piñeyro, P.; Derscheid, R.; Hause, B.; Byers, E.; Dion, K.; Long, D.; Sievers, C.; Tangen, J.; Williams, T.; et al. PCV3-associated disease in the United States swine herd. Emerg. Microbes Infect. 2019, 8, 684–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, T.; Chae, C. First isolation and genetic characterization of porcine circovirus type 3 using primary porcine kidney cells. Vet. Microbiol. 2020, 241, 108576. [Google Scholar] [CrossRef] [PubMed]
- Mora-Díaz, J.; Piñeyro, P.; Shen, H.; Schwartz, K.; Vannucci, F.; Li, G.; Arruda, B.; Giménez-Lirola, L. Isolation of PCV3 from Perinatal and Reproductive Cases of PCV3-Associated Disease and In Vivo Characterization of PCV3 Replication in CD/CD Growing Pigs. Viruses 2020, 12, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Wang, D.; Wang, J.; Zhu, S.; She, R.; Ren, X.; Tian, J.; Quan, R.; Hou, L.; Li, Z.; et al. Induction of Porcine Dermatitis and Nephropathy Syndrome in Piglets by Infection with Porcine Circovirus Type 3. J. Virol. 2019, 93, e02045-18. [Google Scholar] [CrossRef] [Green Version]
- Temeeyasen, G.; Lierman, S.; Arruda, B.L.; Main, R.; Vannucci, F.; Gimenez-Lirola, L.G.; Piñeyro, P.E. Pathogenicity and immune response against porcine circovirus type 3 infection in caesarean-derived, colostrum-deprived pigs. J. Gen. Virol. 2020, 102, jgv001502. [Google Scholar] [CrossRef]
- Bera, B.C.; Choudhary, M.; Anand, T.; Virmani, N.; Sundaram, K.; Choudhary, B.; Tripathi, B.N. Detection and genetic characterization of porcine circovirus 3 (PCV3) in pigs in India. Transbound. Emerg. Dis. 2020, 67, 1062–1067. [Google Scholar] [CrossRef]
- Jiang, H.; Wei, L.; Wang, D.; Wang, J.; Zhu, S.; She, R.; Liu, T.; Tian, J.; Quan, R.; Hou, L.; et al. ITRAQ-based quantitative proteomics reveals the first proteome profiles of piglets infected with porcine circovirus type 3. J. Proteom. 2020, 212, 103598. [Google Scholar] [CrossRef] [PubMed]
- Marks, F.S.; Almeida, L.L.; Driemeier, D.; Canal, C.; de Barcellos, D.E.S.N.; Guimarães, J.A.; Reck, J. Porcine circovirus 2 (PCV2) increases the expression of endothelial adhesion/junction molecules. Braz. J. Microbiol. 2016, 47, 870–875. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, X.; Cheng, A.; Wang, M.; Yin, Z.; Huang, J.; Jia, R. Apoptosis Triggered by ORF3 Proteins of the Circoviridae Family. Front. Cell. Infect. Microbiol. 2021, 10, 609071. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Villamor, E.; Quezada, M.; Bautista, M.; Romanini, S.; Carrasco, L.; Salguero, F.J.; Gómez-Villamandos, J. Classical Swine Fever: Pathogenesis of Glomerular Damage and Immunocharacterization of Immunocomplex Deposits. J. Comp. Pathol. 2001, 124, 246–254. [Google Scholar] [CrossRef]
- Wellenberg, G.; Stockhofe-Zurwieden, N.; de Jong, M.; Boersma, W.; Elbers, A. Excessive porcine circovirus type 2 antibody titres may trigger the development of porcine dermatitis and nephropathy syndrome: A case-control study. Vet. Microbiol. 2004, 99, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Thibault, S.; Drolet, R.; Germain, M.-C.; D’Allaire, S.; LaRochelle, R.; Magar, R. Cutaneous and Systemic Necrotizing Vasculitis in Swine. Vet. Pathol. 1998, 35, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Meng, X.-J.; Halbur, P.G. Porcine Circovirus Type 2–Associated Disease: Update on Current Terminology, Clinical Manifestations, Pathogenesis, Diagnosis, and Intervention Strategies. J. Vet. Diagn. Investig. 2007, 19, 591–615. [Google Scholar] [CrossRef]
- Sipos, W.; Duvigneau, J.; Pietschmann, P.; Schilcher, F.; Hofbauer, G.; Hartl, R.; Schmoll, F. Porcine dermatitis and nephropathy syndrome (PDNS) is associated with a systemic cytokine expression profile indicative of proinflammation and a Th1 bias. Vet. Immunol. Immunopathol. 2005, 107, 303–313. [Google Scholar] [CrossRef]
- Handzel, Z.T.; Busse, W.W.; Sedgwick, J.B.; Vrtis, R.; Lee, W.M.; Kelly, E.; E Gern, J. Eosinophils bind rhinovirus and activate virus-specific T cells. J. Immunol. 1998, 160, 1279–1284. [Google Scholar]
- Samarasinghe, A.E.; Melo, R.C.N.; Duan, S.; LeMessurier, K.; Liedmann, S.; Surman, S.L.; Lee, J.J.; Hurwitz, J.L.; Thomas, P.G.; McCullers, J.A. Eosinophils Promote Antiviral Immunity in Mice Infected with Influenza A Virus. J. Immunol. 2017, 198, 3214–3226. [Google Scholar] [CrossRef]
- Shi, H.-Z. Eosinophils function as antigen-presenting cells. J. Leukoc. Biol. 2004, 76, 520–527. [Google Scholar] [CrossRef]
- Flores-Torres, A.S.; Salinas-Carmona, M.C.; Salinas, E.; Rosas-Taraco, A.G. Eosinophils and Respiratory Viruses. Viral Immunol. 2019, 32, 198–207. [Google Scholar] [CrossRef]
- Zou, Y.; Zhang, N.; Zhang, J.; Zhang, S.; Jiang, Y.; Wang, D.; Tan, Q.; Yang, Y.; Wang, N. Molecular detection and sequence analysis of porcine circovirus type 3 in sow sera from farms with prolonged histories of reproductive problems in Hunan, China. Arch. Virol. 2018, 163, 2841–2847. [Google Scholar] [CrossRef]
- Deim, Z.; Dencső, L.; Erdélyi, I.; Valappil, S.K.; Varga, C.; Pósa, A.; Makrai, L.; Rákhely, G. Porcine circovirus type 3 detection in a Hungarian pig farm experiencing reproductive failures. Vet. Rec. 2019, 185, 84. [Google Scholar] [CrossRef] [PubMed]
- Saporiti, V.; Valls, L.; Maldonado, J.; Perez, M.; Correa-Fiz, F.; Segalés, J.; Sibila, M. Porcine Circovirus 3 Detection in Aborted Fetuses and Stillborn Piglets from Swine Reproductive Failure Cases. Viruses 2021, 13, 264. [Google Scholar] [CrossRef] [PubMed]
- Serena, M.S.; Cappuccio, J.A.; Barrales, H.; Metz, G.E.; Aspitia, C.G.; Lozada, I.; Perfumo, C.J.; Quiroga, M.A.; Piñeyro, P.; Echeverría, M.G. First detection and genetic characterization of porcine circovirus type 3 (PCV3) in Argentina and its association with reproductive failure. Transbound. Emerg. Dis. 2020, 68, 1761–1766. [Google Scholar] [CrossRef]
- Brunborg, I.M.; Jonassen, C.M.; Moldal, T.; Bratberg, B.; Lium, B.; Koenen, F.; Schönheit, J. Association of Myocarditis with High Viral Load of Porcine Circovirus Type 2 in Several Tissues in Cases of Fetal Death and High Mortality in Piglets. A Case Study. J. Vet. Diagn. Investig. 2007, 19, 368–375. [Google Scholar] [CrossRef]
- Faccini, S.; Barbieri, I.; Gilioli, A.; Sala, G.; Gibelli, L.R.; Moreno, A.; Sacchi, C.; Rosignoli, C.; Franzini, G.; Nigrelli, A. Detection and genetic characterization of Porcine circovirus type 3 in Italy. Transbound. Emerg. Dis. 2017, 64, 1661–1664. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Wu, X.; Zhang, L.; Xin, C.; Liu, Y.; Shi, J.; Peng, Z.; Xu, S.; Fu, F.; Yu, J.; et al. The occurrence of porcine circovirus 3 without clinical infection signs in Shandong Province. Transbound. Emerg. Dis. 2017, 64, 1337–1341. [Google Scholar] [CrossRef]
- Giudici, S.D.; Franzoni, G.; Bonelli, P.; Angioi, P.P.; Zinellu, S.; Deriu, V.; Carta, T.; Sechi, A.M.; Salis, F.; Balzano, F.; et al. Genetic Characterization of Porcine Circovirus 3 Strains Circulating in Sardinian Pigs and Wild Boars. Pathogens 2020, 9, 344. [Google Scholar] [CrossRef] [PubMed]
- Sarli, G.; Morandi, F.; Panarese, S.; Bacci, B.; Ferrara, D.; Bianco, C.; Fusaro, L.; Bacci, M.L.; Galeati, G.; Dottori, M.; et al. Reproduction in porcine circovirus type 2 (PCV2) seropositive gilts inseminated with PCV2b spiked semen. Acta Vet. Scand. 2012, 54, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.; Zhang, M.; Liu, Q.; Cao, Y.; Zhang, W.; Liang, Y.; Song, X.; Ji, K.; Shao, Y.; Qi, K.; et al. Epidemiology and Evolution of Emerging Porcine Circovirus-like Viruses in Pigs with Hemorrhagic Dysentery and Diarrhea Symptoms in Central China from 2018 to 2021. Viruses 2021, 13, 2282. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Wang, J.; Zhang, W.; Quan, R.; Wang, D.; Zhu, S.; Jiang, H.; Wei, L.; Liu, J. Dynamic Alterations of Gut Microbiota in Porcine Circovirus Type 3-Infected Piglets. Front. Microbiol. 2020, 11, 1360. [Google Scholar] [CrossRef]
- Woźniak, A.; Miłek, D.; Stadejek, T. Wide Range of the Prevalence and Viral Loads of Porcine Circovirus Type 3 (PCV3) in Different Clinical Materials from 21 Polish Pig Farms. Pathogens 2020, 9, 411. [Google Scholar] [CrossRef]
- Tochetto, C.; de Lima, D.A.; Varela, A.P.M.; Ortiz, L.C.; Loiko, M.R.; Scheffer, C.M.; Paim, W.P.; Cibulski, S.P.; Cerva, C.; Herpich, J.; et al. Investigation on porcine circovirus type 3 in serum of farrowing sows with stillbirths. Microb. Pathog. 2020, 149, 104316. [Google Scholar] [CrossRef]
- Opriessnig, T.; McKeown, N.E.; Zhou, E.-M.; Meng, X.-J.; Halbur, P.G. Genetic and experimental comparison of porcine circovirus type 2 (PCV2) isolates from cases with and without PCV2-associated lesions provides evidence for differences in virulence. J. Gen. Virol. 2006, 87, 2923–2932. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Prickett, J.R.; Madson, D.M.; Shen, H.-G.; Juhan, N.M.; Pogranichniy, R.M.; Meng, X.; Halbur, P.G. Porcine circovirus type 2 (PCV2)-infection and re-inoculation with homologous or heterologous strains: Virological, serological, pathological and clinical effects in growing pigs. Vet. Res. 2010, 41, 31. [Google Scholar] [CrossRef] [Green Version]
- Fux, R.; Söckler, C.; Link, E.K.; Renken, C.; Krejci, R.; Sutter, G.; Ritzmann, M.; Eddicks, M. Full genome characterization of porcine circovirus type 3 isolates reveals the existence of two distinct groups of virus strains. Virol. J. 2018, 15, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Wu, J.; Jiang, M.; Xie, Y.; Bu, W.; Liu, C.; Zhang, G.; Luo, M. A Novel Technique for Constructing Infectious Cloning of Type 3 Porcine Circovirus. Front. Microbiol. 2020, 11, 1067. [Google Scholar] [CrossRef]
- Ellis, J.; Krakowka, S.; Lairmore, M.; Haines, D.; Bratanich, A.; Clark, E.; Allan, G.; Konoby, C.; Hassard, L.; Meehan, B.; et al. Reproduction of Lesions of Postweaning Multisystemic Wasting Syndrome in Gnotobiotic Piglets. J. Vet. Diagn. Investig. 1999, 11, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, T.; Zhang, X.; Liu, X.; Ren, L. Co-Infection of Swine with Porcine Circovirus Type 2 and Other Swine Viruses. Viruses 2019, 11, 185. [Google Scholar] [CrossRef] [Green Version]
- Opriessnig, T.; Fenaux, M.; Thomas, P.; Hoogland, M.J.; Rothschild, M.F.; Meng, X.J.; Halbur, P.G. Evidence of Breed-dependent Differences in Susceptibility to Porcine Circovirus Type-2-associated Disease and Lesions. Vet. Pathol. 2006, 43, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Patterson, A.R.; Madson, D.M.; Pal, N.; Rothschild, M.; Kuhar, D.; Lunney, J.K.; Juhan, N.M.; Meng, X.J.; Halbur, P.G. Difference in severity of porcine circovirus type two-induced pathological lesions between Landrace and Pietrain pigs1. J. Anim. Sci. 2009, 87, 1582–1590. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, H.; Wang, P.; Wang, L.; Sun, Y.; Liu, G.; Zhang, P.; Kang, L.; Jiang, S.; Jiang, Y. RNA-Seq Analysis Reveals Genes Underlying Different Disease Responses to Porcine Circovirus Type 2 in Pigs. PLoS ONE 2016, 11, e0155502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baghbani, T.; Nikzad, H.; Azadbakht, J.; Izadpanah, F.; Kashani, H.H. Dual and mutual interaction between microbiota and viral infections: A possible treat for COVID-19. Microb. Cell Fact. 2020, 19, 1–25. [Google Scholar] [CrossRef]
- Dominguez-Diaz, C.; Garcia-Orozco, A.; Riera-Leal, A.; Padilla-Arellano, J.R.; Fafutis-Morris, M. Microbiota and Its Role on Viral Evasion: Is It With Us or Against Us? Front. Cell. Infect. Microbiol. 2019, 9, 256. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Ma, W.-T.; Pang, M.; Fan, Q.-L.; Hua, J.-L. The Commensal Microbiota and Viral Infection: A Comprehensive Review. Front. Immunol. 2019, 10, 1551. [Google Scholar] [CrossRef]
- Harms, P.A.; Sorden, S.D.; Halbur, P.G.; Bolin, S.R.; Lager, K.M.; Morozov, I.; Paul, P.S. Experimental Reproduction of Severe Disease in CD/CD Pigs Concurrently Infected with Type 2 Porcine Circovirus and Porcine Reproductive and Respiratory Syndrome Virus. Vet. Pathol. 2001, 38, 528–539. [Google Scholar] [CrossRef]
- Lin, C.-M.; Hou, Y.; Marthaler, D.; Gao, X.; Liu, X.; Zheng, L.; Saif, L.J.; Wang, Q. Attenuation of an original US porcine epidemic diarrhea virus strain PC22A via serial cell culture passage. Vet. Microbiol. 2017, 201, 62–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolin, S.R.; Stoffregen, W.C.; Nayar, G.P.S.; Hamel, A.L. Postweaning Multisystemic Wasting Syndrome Induced after Experimental Inoculation of Cesarean-Derived, Colostrum-Deprived Piglets with Type 2 Porcine Circovirus. J. Vet. Diagn. Investig. 2001, 13, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Okuda, Y.; Ono, M.; Yazawa, S.; Shibata, I. Experimental Reproduction of Postweaning Multisystemic Wasting Syndrome in Cesarean-Derived, Colostrum-Deprived Piglets Inoculated with Porcine Circovirus Type 2 (PCV2): Investigation of Quantitative PCV2 Distribution and Antibody Responses. J. Vet. Diagn. Investig. 2003, 15, 107–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Radlowski, E.C.; Monaco, M.H.; Fahey, J.G.C.; Gaskins, H.R.; Donovan, S.M. Mode of Delivery and Early Nutrition Modulate Microbial Colonization and Fermentation Products in Neonatal Piglets. J. Nutr. 2013, 143, 795–803. [Google Scholar] [CrossRef] [Green Version]
- Cortes, L.C.P.; Leveque, R.M.; Funk, J.; Marsh, T.L.; Mulks, M.H. Development of the tonsillar microbiome in pigs from newborn through weaning. BMC Microbiol. 2018, 18, 35. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Giménez-Lirola, L.G.; Halbur, P.G. Polymicrobial respiratory disease in pigs. Anim. Health Res. Rev. 2011, 12, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.; Pors, S.; Jensen, H.; Bille-Hansen, V.; Bisgaard, M.; Flachs, E.; Nielsen, O. An Investigation of the Pathology and Pathogens Associated with Porcine Respiratory Disease Complex in Denmark. J. Comp. Pathol. 2010, 143, 120–131. [Google Scholar] [CrossRef]
- García, N.; Fernández-Garayzábal, J.F.; Goyache, J.; Domínguez, L.; Vela, A.I. Associations between biovar and virulence factor genes in Pasteurella multocida isolates from pigs in Spain. Vet. Rec. 2011, 169, 362. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Hensley, C.; Mahsoub, H.M.; Ramesh, A.K.; Zhou, P. Microbiota in viral infection and disease in humans and farm animals. Prog. Mol. Biol. Transl. Sci. 2020, 171, 15–60. [Google Scholar] [CrossRef]


{kind=link}
| Parameters | Study A | Study B | Study C | Study D |
|---|---|---|---|---|
| Virus | ||||
| Name a | LY (MF318451) | ISU27734 (MK058528) | N/A | JX (MK656956) |
| Origin | China, 2015 | USA, 2018 | N/A | China, 2018 |
| Farms of origin | ||||
| Clinical signs/lesions | PDNS-like lesions | Lymphocytic myocarditis | N/A | Diarrhea |
| Affected pigs | Piglet | Piglet (8 days) | Fetus and suckling piglet | Suckling and weaned pigs |
| Inocula | ||||
| Infectious material | Virus isolate (infective clone) | Virus isolate | Tissue homogenate | Intestinal content |
| Route and titer b | IN: 2 × 106.53 TCID50 | IN: 6.6 × 1010 gc IM: 6.6 × 1010 gc | IN: 2.04 × 1011 gc IM: 6.38 × 1012 gc (twice, 7 days apart) | Oral: 3.0 × 106.5 gc |
| Inoculated pigs | ||||
| Breed | Duroc × Large White | N/A | N/A | N/A |
| Age | 4 and 8 weeks old | 6 weeks old | 5 weeks old | 3 weeks old |
| Farrowing status | Conventional | CD/CD | CD/CD | N/A |
| PCV3 detection | ||||
| Virus titer | qPCR (ORF2) | qPCR (ORF2) | qPCR (ORF2) | None |
| Tropism | IHC (PCV3 antigen) | ISH (ORF1 mRNA) | ISH (ORF1 mRNA) | None |
| Antibody detection | ||||
| Target | Capsid | Capsid | Capsid | None |
| Isotype | N/A | IgM and IgG | IgM and IgG | None |
| Duration of studies | 28 days | 28 days | 42 days | 7 days |
| Parameter | Study A | Study B | Study C |
|---|---|---|---|
| Disease severity | +++ | + | + |
| Number of affected tissues | +++ | + | ++ |
| Degree of tissue tropism a | +++ | + | ++ |
| Peak viremia titer | +++ | + | ++ |
| Onset of anti-PCV3 antibody b | +++ | −/+ | + |
| Organ | Study A | Study B | Study C |
|---|---|---|---|
| Heart | IHC: + Myocarditis Myolysis and epicardial necrosis Hemorrhage Eosinophil infiltration | ISH: + Lymphoplasmacytic myocarditis Periarteritis | ISH: + Lymphoplasmacytic myocarditis and periarteritis |
| Kidney | IHC: + Interstitial glomerulonephritis | ISH: + Lymphoplasmacytic interstitial nephritis Periarteritis | ISH: + Lymphoplasmacytic interstitial nephritis and periarteritis |
| Intestine | IHC: + Lymphocyte infiltration Epithelial degeneration/necrosis Eosinophil infiltration | ISH: − Lymphoplasmacytic periarteritis Arteritis of the serosa | ISH: + Lymphoplasmacytic periarteritis and arteritis of the serosa |
| Spleen | IHC: + Lymphoid depletion/necrosis Hemorrhage | Absent (ISH and H&E) | ISH: + Lymphoplasmacytic periarteritis and arteritis |
| Liver | IHC: + Congestion Hepatocyte atrophy | Absent (ISH and H&E) | ISH: + Lymphoplasmacytic hepatitis |
| Lung | IHC: + Lymphoplasmacytic and histiocytic bronchointersitial pneumonia | Absent (ISH and H&E) | Absent (ISH and H&E) |
| TLN | IHC: + Lymphocytic necrosis Congestion and hemorrhage Eosinophil infiltration | Absent (ISH and H&E) | Absent (ISH and H&E) |
| MLN | IHC: + Lymphoid depletion/necrosis Eosinophil infiltration | Absent (ISH and H&E) | Absent (ISH and H&E) |
| ILN | IHC: + Lymphoid depletion Hyperplasia of epithelial-like cells Eosinophil infiltration | Absent (ISH and H&E) | Absent (ISH and H&E) |
| Brain | N/A (IHC and H&E) | Absent (ISH and H&E) | ISH: − Lymphoplasmacytic encephalitis with perivasculitis |
| Amino Acid Positions of Each ORF | Study A LY (MF318451 a) | Study B ISU27734 (MK058528 a) | Study D JX (MK656956 a) |
|---|---|---|---|
| ORF1 | |||
| aa 122 | S | S | A |
| aa 278 | C | F | C |
| ORF2 | |||
| aa 24 | V | A | V |
| aa 27 | K | R | K |
| aa 95 | F | S | S |
| aa 150 | L | I | I |
| ORF3231 | |||
| aa 1 | F | S | F |
| aa 4 | D | G | D |
| aa 12 | S | A | S |
| aa 32 | V | L | L |
| aa 227 b | V | V | G |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sirisereewan, C.; Thanawongnuwech, R.; Kedkovid, R. Current Understanding of the Pathogenesis of Porcine Circovirus 3. Pathogens 2022, 11, 64. https://doi.org/10.3390/pathogens11010064
Sirisereewan C, Thanawongnuwech R, Kedkovid R. Current Understanding of the Pathogenesis of Porcine Circovirus 3. Pathogens. 2022; 11(1):64. https://doi.org/10.3390/pathogens11010064
Chicago/Turabian StyleSirisereewan, Chaitawat, Roongroje Thanawongnuwech, and Roongtham Kedkovid. 2022. "Current Understanding of the Pathogenesis of Porcine Circovirus 3" Pathogens 11, no. 1: 64. https://doi.org/10.3390/pathogens11010064