
Non-Assembled ORF2 Capsid Protein of Porcine Circovirus 2b Does Not Confer Protective Immunity

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. No PCVAD Was Observed after Challenge and No Virological Protection Was Induced by the Experimental PCV2 Vaccine
2.2. The Recombinant Cap Vaccine Did Not Induce a Significant Antibody Response to PCV2
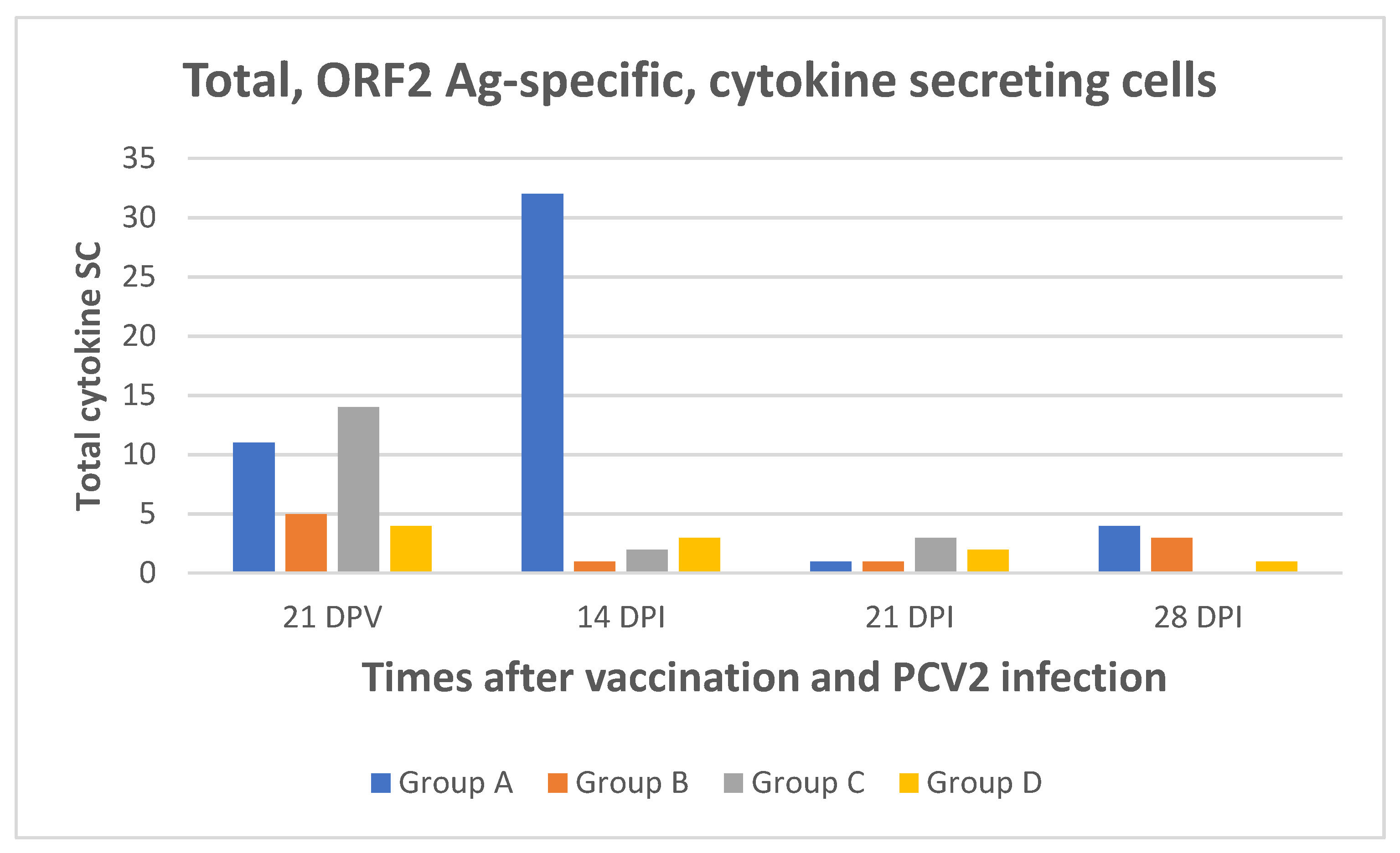
2.3. Poor Cell-Mediated Immune Responses to PCV2 Were Induced by the Cap Vaccine
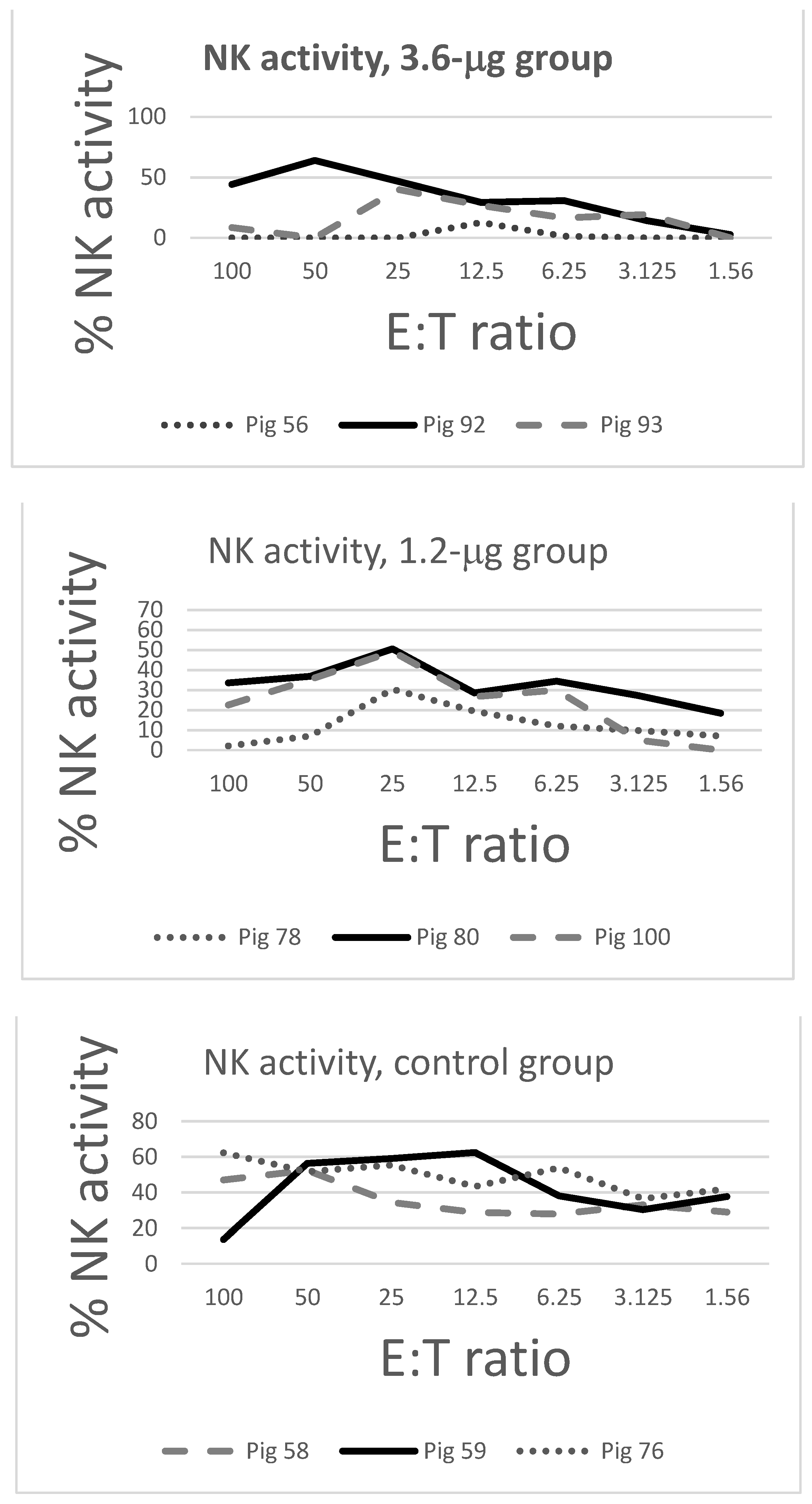
2.4. NK Assays
2.5. Histology
3. Discussion
4. Materials and Methods
4.1. Virus and Cells
4.2. Recombinant ORF2 Antigen and Monoclonal Antibodies
4.3. Vaccine Formulation
4.4. Experimental Design
4.5. Total and Neutralizing Anti-PCV2 Antibodies
4.6. PCV2 DNA in Serum
4.7. PCV2-Specific Interferon-γ Release Assay
4.8. Proliferation Assay with Swine PBMC
4.9. PCV2-Specific, IFN γ-Positive T Lymphocytes
4.10. ELISPOT Assays
4.11. Natural Killer (NK) Assay with K-562 Cells
4.12. Necropsy and Histopathology
4.13. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Segales, J. Porcine circovirus type 2 (PCV2) infections: Clinical signs, pathology and laboratory diagnosis. Virus Res. 2012, 164, 10–19. [Google Scholar] [CrossRef]
- Segales, J.; Olvera, A.; Grau-Roma, L.; Charreyre, C.; Nauwynck, H.; Larsen, L.; Dupont, K.; McCullough, K.; Ellis, J.; Krakowka, S.; et al. PCV-2 genotype definition and nomenclature. Vet. Rec. 2008, 162, 867–868. [Google Scholar] [CrossRef]
- Alarcon, P.; Velasova, M.; Mastin, A.; Nevel, A.; Stark, K.D.; Wieland, B. Farm level risk factors associated with severity of post-weaning multi-systemic wasting syndrome. Prev. Vet. Med. 2011, 101, 182–191. [Google Scholar] [CrossRef]
- Segales, J. Best practice and future challenges for vaccination against porcine circovirus type 2. Expert Rev. Vaccines 2015, 14, 473–487. [Google Scholar] [CrossRef]
- Harmon, K.M.; Gauger, P.C.; Zhang, J.; Pineyro, P.E.; Dunn, D.D.; Chriswell, A.J. Whole-Genome Sequences of Novel Porcine Circovirus Type 2 Viruses Detected in Swine from Mexico and the United States. Genome Announc. 2015, 3, e01315-15. [Google Scholar] [CrossRef] [Green Version]
- Park, K.H.; Oh, T.; Yang, S.; Cho, H.; Kang, I.; Chae, C. Evaluation of a porcine circovirus type 2a (PCV2a) vaccine efficacy against experimental PCV2a, PCV2b, and PCV2d challenge. Vet. Microbiol. 2019, 231, 87–92. [Google Scholar] [CrossRef]
- Franzo, G.; Segales, J. Porcine Circovirus 2 Genotypes, Immunity and Vaccines: Multiple Genotypes but One Single Serotype. Pathogens 2020, 9, 1049. [Google Scholar] [CrossRef]
- Zanotti, C.; Martinelli, N.; Lelli, D.; Amadori, M. Correlates of Protection Following Vaccination with Inactivated Porcine Circovirus 2 Vaccines. Viral. Immunol. 2015, 28, 600–608. [Google Scholar] [CrossRef]
- Guarneri, F.; Tresoldi, E.T.; Sarli, G.; Boniotti, M.B.; Lelli, D.; Barbieri, I.; Bacci, B.; D’Annunzio, G.; Amadori, M. Protective immunity in swine induced by Porcine Circovirus 2b inactivated vaccines with different antigen payload. Vet. Microbiol. 2021, 252, 108887. [Google Scholar] [CrossRef]
- Oh, Y.; Seo, H.W.; Han, K.; Park, C.; Chae, C. Protective effect of the maternally derived porcine circovirus type 2 (PCV2)-specific cellular immune response in piglets by dam vaccination against PCV2 challenge. J. Gen. Virol. 2012, 93, 1556–1562. [Google Scholar] [CrossRef] [Green Version]
- Salmon, H.; Berri, M.; Gerdts, V.; Meurens, F. Humoral and cellular factors of maternal immunity in swine. Dev. Comp. Immunol. 2009, 33, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Patterson, A.R.; Elsener, J.; Meng, X.J.; Halbur, P.G. Influence of maternal antibodies on efficacy of porcine circovirus type 2 (PCV2) vaccination to protect pigs from experimental infection with PCV2. Clin. Vaccine Immunol. 2008, 15, 397–401. [Google Scholar] [CrossRef] [Green Version]
- Sobrino, F.; Saiz, M.; Jimenez-Clavero, M.A.; Nunez, J.I.; Rosas, M.F.; Baranowski, E.; Ley, V. Foot-and-mouth disease virus: A long known virus, but a current threat. Vet. Res. 2001, 32, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Colbert, J.D.; Cruz, F.M.; Rock, K.L. Cross-presentation of exogenous antigens on MHC I molecules. Curr. Opin. Immunol. 2020, 64, 1–8. [Google Scholar] [CrossRef]
- Tsan, M.F.; Gao, B. Heat shock proteins and immune system. J. Leukoc. Biol. 2009, 85, 905–910. [Google Scholar] [CrossRef]
- Grgacic, E.V.; Anderson, D.A. Virus-like particles: Passport to immune recognition. Methods 2006, 40, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, C.; Wagner, R. Virus-like particles-universal molecular toolboxes. Curr. Opin. Biotechnol. 2007, 18, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Koinig, H.C.; Talker, S.C.; Stadler, M.; Ladinig, A.; Graage, R.; Ritzmann, M.; Hennig-Pauka, I.; Gerner, W.; Saalmuller, A. PCV2 vaccination induces IFN-gamma/TNF-alpha co-producing T cells with a potential role in protection. Vet. Res. 2015, 46, 20. [Google Scholar] [CrossRef] [Green Version]
- Opriessnig, T.; Halbur, P.G. Concurrent infections are important for expression of porcine circovirus associated disease. Virus Res. 2012, 164, 20–32. [Google Scholar] [CrossRef]
- Solis Worsfold, C.; Dardari, R.; Law, S.; Eschbaumer, M.; Nourozieh, N.; Marshall, F.; Czub, M. Assessment of neutralizing and non-neutralizing antibody responses against Porcine circovirus 2 in vaccinated and non-vaccinated farmed pigs. J. Gen. Virol 2015, 96, 2743–2748. [Google Scholar] [CrossRef]
- Fort, M.; Olvera, A.; Sibila, M.; Segales, J.; Mateu, E. Detection of neutralizing antibodies in postweaning multisystemic wasting syndrome (PMWS)-affected and non-PMWS-affected pigs. Vet. Microbiol. 2007, 125, 244–255. [Google Scholar] [CrossRef] [Green Version]
- Razzuoli, E.; Faggionato, E.; Dotti, S.; Villa, R.; Lombardo, T.; Boizza, L.; Ferrari, M.; Amadori, M. Isolation and culture of pig tonsil lymphocytes. Vet. Immunol. Immunopathol. 2012, 148, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Chambers, A.C.; Aksular, M.; Graves, L.P.; Irons, S.L.; Possee, R.D.; King, L.A. Overview of the Baculovirus Expression System. Curr. Protoc. Protein Sci. 2018, 91, 5.4.1–5.4.6. [Google Scholar] [CrossRef]
- Sala, G.; Rigola, S.; Alborali, G.L.; Brocchi, E.; Cordioli, P. Development of monoclonal antibodies-based ELISAs for the detection of antibodies against porcine circovirus type 1 and type 2. In Proceedings of the 5th International Congress of Veterinary Virology, Brescia, Italy, 27–30 August 2000; pp. 253–254. [Google Scholar]
- Olvera, A.; Sibila, M.; Calsamiglia, M.; Segales, J.; Domingo, M. Comparison of porcine circovirus type 2 load in serum quantified by a real time PCR in postweaning multisystemic wasting syndrome and porcine dermatitis and nephropathy syndrome naturally affected pigs. J. Virol. Methods 2004, 117, 75–80. [Google Scholar] [CrossRef]
- Steiner, E.; Balmelli, C.; Gerber, H.; Summerfield, A.; McCullough, K. Cellular adaptive immune response against porcine circovirus type 2 in subclinically infected pigs. BMC Vet. Res. 2009, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Guarneri, F.; Tresoldi, E.T.; Sarli, G.; Boniotti, M.B.; Lelli, D.; Barbieri, I.; Bacci, B.; D’Annunzio, G.; Amadori, M. Dataset of immune responses induced in swine by an inactivated Porcine Circovirus 2b vaccine. Data Brief. 2021, 35, 106906. [Google Scholar] [CrossRef] [PubMed]
- Boulet, S.; Ndongala, M.L.; Peretz, Y.; Boisvert, M.P.; Boulassel, M.R.; Tremblay, C.; Routy, J.P.; Sekaly, R.P.; Bernard, N.F. A dual color ELISPOT method for the simultaneous detection of IL-2 and IFN-gamma HIV-specific immune responses. J. Immunol. Methods 2007, 320, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Welter, A.; Sundararaman, S.; Li, R.; Zhang, T.; Karulin, A.Y.; Lehmann, A.; Naeem, V.; Roen, D.R.; Kuerten, S.; Lehmann, P.V. High-Throughput GLP-Capable Target Cell Visualization Assay for Measuring Cell-Mediated Cytotoxicity. Cells 2018, 7, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group D, Control PCV2 Genome Copies/mL | Group C, 1.2 μg ORF2/Dose PCV2 Genome Copies/mL | ||||||||||||
| Pig | DPV0 | DPV 21 | DPI 7 | DPI 14 | DPI 21 | DPI 28 | Pig | DPV 0 | DPV 21 | DPI 7 | DPI 14 | DPI 21 | DPI 28 |
| 057 * | NEG | - | - | - | - | - | 077 | NEG | NEG | NEG | 35.7 × 106 | 1.4 × 106 | 3.79 × 105 |
| 058 | NEG | NEG | NEG | 4.25 × 106 | 3.31 × 105 | 4.48 × 104 | 078 | NEG | NEG | NEG | 25.3 × 106 | 1.33 × 106 | 8.69 × 104 |
| 059 | NEG | NEG | NEG | NEG | 17.2 × 106 | 1.33 × 105 | 079 | NEG | NEG | NEG | 18.4 × 106 | 6.13 × 105 | 3.91 × 104 |
| 060 | NEG | NEG | NEG | 2.08 × 106 | 1.45 × 105 | 104 | 080 | NEG | NEG | NEG | 8.86 × 103 | 3.05 × 104 | 1.74 × 103 |
| 076 | NEG | NEG | NEG | NEG | 4.89 × 103 | NEG | 100 | NEG | NEG | NEG | 15.6 × 106 | 2.68 × 105 | 9.07 × 104 |
| Group B, 3.6 μg ORF2/Dose PCV2 Genome Copies/mL | Group A, 10.8 μg ORF2/Dose PCV2 Genome Copies/mL | ||||||||||||
| Pig | DPV 0 | DPV 21 | DPI 7 | DPI 14 | DPI 21 | DPI 28 | Pig | DPV 0 | DPV 21 | DPI 7 | DPI 14 | DPI 21 | DPI 28 |
| 056 | NEG | NEG | NEG | NEG | 1.52 × 104 | 3.74 × 104 | 091 | NEG | NEG | NEG | 4.89 × 105 | 8.00 × 105 | 3.53 × 104 |
| 092 | NEG | NEG | NEG | 2.23 × 105 | NEG | 1.28 × 104 | 095 | NEG | NEG | NEG | NEG | 5.06 × 104 | 9.92 × 104 |
| 093 | NEG | NEG | NEG | 3.31 × 105 | 1.34 × 104 | 2.5 × 103 | 097 | NEG | NEG | NEG | 2.08 × 104 | 2.04 × 104 | 4.29 × 103 |
| 094 | NEG | NEG | NEG | 1.51 × 105 | 8.69 × 104 | 3.17 × 103 | 098 | NEG | NEG | NEG | 2.49 × 105 | 2.76 × 105 | 2.17 × 104 |
| 096 | NEG | NEG | NEG | NEG | 2.02 × 105 | 8.18 × 104 | 099 | NEG | NEG | NEG | 3.45 × 106 | 4.43 × 105 | 4.69 × 104 |
| Control, PBS + Adjuvant | 1.2 μg ORF2/Dose | ||||||
| Group D | Group C | ||||||
| Number | Litter | Starting Titer | Titer DPV 21 | Number | Litter | Starting Titer | Titer DPV 21 |
| 057 * | White | 1/1000 | - | 080 | White | 1/1000 | 1/100 |
| 076 | White | 1/1000 | 1/100 | 100 | White | 1/100 | 1/100 |
| 059 | Green | 1/1000 | 1/100 | 077 | Green | 1/1000 | 1/100 |
| 060 | Green | 1/1000 | 1/100 | 078 | Green | 1/1000 | 1/100 |
| 058 | Red | 1/1000 | 1/100 | 079 | Red | 1/1000 | 1/1000 |
| 3.6 μg ORF2/Dose | 10.8 μg ORF2/Dose | ||||||
| Group B | Group A | ||||||
| Number | Litter | Starting Titer | Titer DPV 21 | Number | Litter | Starting Titer | Titer DPV 21 |
| 092 | White | 1/1000 | 1/100 | 097 | White | 1/1000 | 1/100 |
| 096 | White | 1/1000 | 1/1000 | 099 | Green | 1/1000 | 1/100 |
| 093 | Green | 1/1000 | 1/1000 | 091 | Red | 1/100 | 1/100 |
| 056 | Red | 1/100 | 1/100 | 095 | Red | 1/1000 | 1/1000 |
| 094 | Red | 1/1000 | 1/100 | 098 | Red | 1/1000 | 1/100 |
| Group D, Control IFN-γ Response (Ag and DmOD) | Group C, 1.2 μg ORF2 Ag/Dose IFN-γ Response (Ag and DmOD) | ||||||||||
| Pig | DPV 21 | DPI 7 | DPI 14 | DPI 21 | DPI 28 | Pig | DPV 21 | DPI 7 | DPI 14 | DPI 21 | DPI 28 |
| 057 * | - | - | - | - | - | 077 | ORF2:NEG PCV2:NEG | ORF2: 63 PCV2:NEG | ORF2: 13 PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG |
| 058 | ORF2: 58 PCV2: 25 | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2: NEG PCV2:NEG | ORF2:NEG PCV2:NEG | 078 | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2: 16 PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2: 46 PCV2:NEG |
| 059 | ORF2:NEG PCV2: 23 | ORF2:NEG PCV2: 17 | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | 079 | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG |
| 060 | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2: 10 PCV2:NEG | 080 | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG |
| 076 | ORF2:NEG PCV2:NEG | ORF2: 22 PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | 100 | ORF2:NEG PCV2: 20 | ND | ORF2:NEG PCV2:NEG | ORF2: 90 PCV2:NEG | ORF2:NEG PCV2:NEG |
| Group B, 3.6 μg ORF2 Ag/dose IFN-γ response (Ag and DmOD) | Group A, 10.8 μg ORF2 Ag/dose IFN-γ response (Ag and DmOD) | ||||||||||
| Pig | DPV 21 | DPI 7 | DPI 14 | DPI 21 | DPI 28 | Pig | DPV 21 | DPI 7 | DPI 14 | DPI 21 | DPI 28 |
| 056 | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2: 34 PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | 091 | ORF2: 29 PCV2:NEG | ORF2: 17 PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG |
| 092 | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2: 10 PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | 095 | ORF2:64 PCV2:NEG | ORF2: 56 PCV2:NEG | ORF2: 42 PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2: 16 PCV2:NEG |
| 093 | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | 097 | ORF2:NEG PCV2:NEG | ORF2: 11 PCV2: 30 | ORF2: 24 PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG |
| 094 | ORF2:NEG PCV2:NEG | ORF2: 36 PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | 098 | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2: 82 PCV2:NEG | ORF2: 15 PCV2:NEG |
| 096 | ORF2:NEG PCV2:NEG | ORF2: 12 PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG | 099 | ORF2: 32 PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2: 17 PCV2:NEG | ORF2:NEG PCV2:NEG | ORF2:NEG PCV2:NEG |
| Group A: 10.8 μg ORF2 Ag | Group B: 3.6 μg ORF2 Ag | ||||||||||
| Pig number | 21 DPV | 7 PI | 14 Pi | 21 PI | 28 PI | Pig number | 21 DPV | 7 PI | 14 Pi | 21 PI | 28 PI |
| 091 | NEG | NEG | NEG | NEG | NEG | 056 | NEG | 8 | NEG | NEG | NEG |
| 095 | NEG | NEG | NEG | NEG | 148 | 092 | NEG | NEG | NEG | NEG | NEG |
| 097 | NEG | NEG | NEG | NEG | NEG | 093 | 38 | NEG | NEG | NEG | NEG |
| 098 | 68 | NEG | NEG | NEG | NEG | 094 | 128 | NEG | NEG | NEG | 144 |
| 099 | 9 | NEG | NEG | NEG | 60 | 096 | NEG | NEG | 43 | NEG | 15 |
| Group C: 1.2μg ORF2 Ag | Group D: control | ||||||||||
| Pig number | 21 DPV | 7 PI | 14 Pi | 21 PI | 28 PI | Pig number | 21 DPV | 7 PI | 14 Pi | 21 PI | 28 PI |
| 077 | NEG | 31 | 45 | NEG | NEG | 057 * | ND | ND | ND | ND | ND |
| 078 | 1 | 52 | NEG | NEG | NEG | 058 | NEG | 72 | NEG | NEG | NEG |
| 079 | NEG | NEG | NEG | NEG | NEG | 059 | NEG | 33 | NEG | NEG | NEG |
| 080 | NEG | NEG | NEG | NEG | NEG | 060 | NEG | 54 | NEG | NEG | NEG |
| 100 | 43 | NEG | NEG | NEG | NEG | 076 | NEG | NEG | 40 | NEG | NEG |
| * Deceased |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guarneri, F.; Tonni, M.; Sarli, G.; Boniotti, M.B.; Lelli, D.; Barbieri, I.; D’Annunzio, G.; Alborali, G.L.; Bacci, B.; Amadori, M. Non-Assembled ORF2 Capsid Protein of Porcine Circovirus 2b Does Not Confer Protective Immunity. Pathogens 2021, 10, 1161. https://doi.org/10.3390/pathogens10091161
Guarneri F, Tonni M, Sarli G, Boniotti MB, Lelli D, Barbieri I, D’Annunzio G, Alborali GL, Bacci B, Amadori M. Non-Assembled ORF2 Capsid Protein of Porcine Circovirus 2b Does Not Confer Protective Immunity. Pathogens. 2021; 10(9):1161. https://doi.org/10.3390/pathogens10091161
Chicago/Turabian StyleGuarneri, Flavia, Matteo Tonni, Giuseppe Sarli, Maria Beatrice Boniotti, Davide Lelli, Ilaria Barbieri, Giulia D’Annunzio, Giovanni Loris Alborali, Barbara Bacci, and Massimo Amadori. 2021. "Non-Assembled ORF2 Capsid Protein of Porcine Circovirus 2b Does Not Confer Protective Immunity" Pathogens 10, no. 9: 1161. https://doi.org/10.3390/pathogens10091161