
A Rapid Method for Performing a Multivariate Optimization of Phage Production Using the RCCD Approach

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
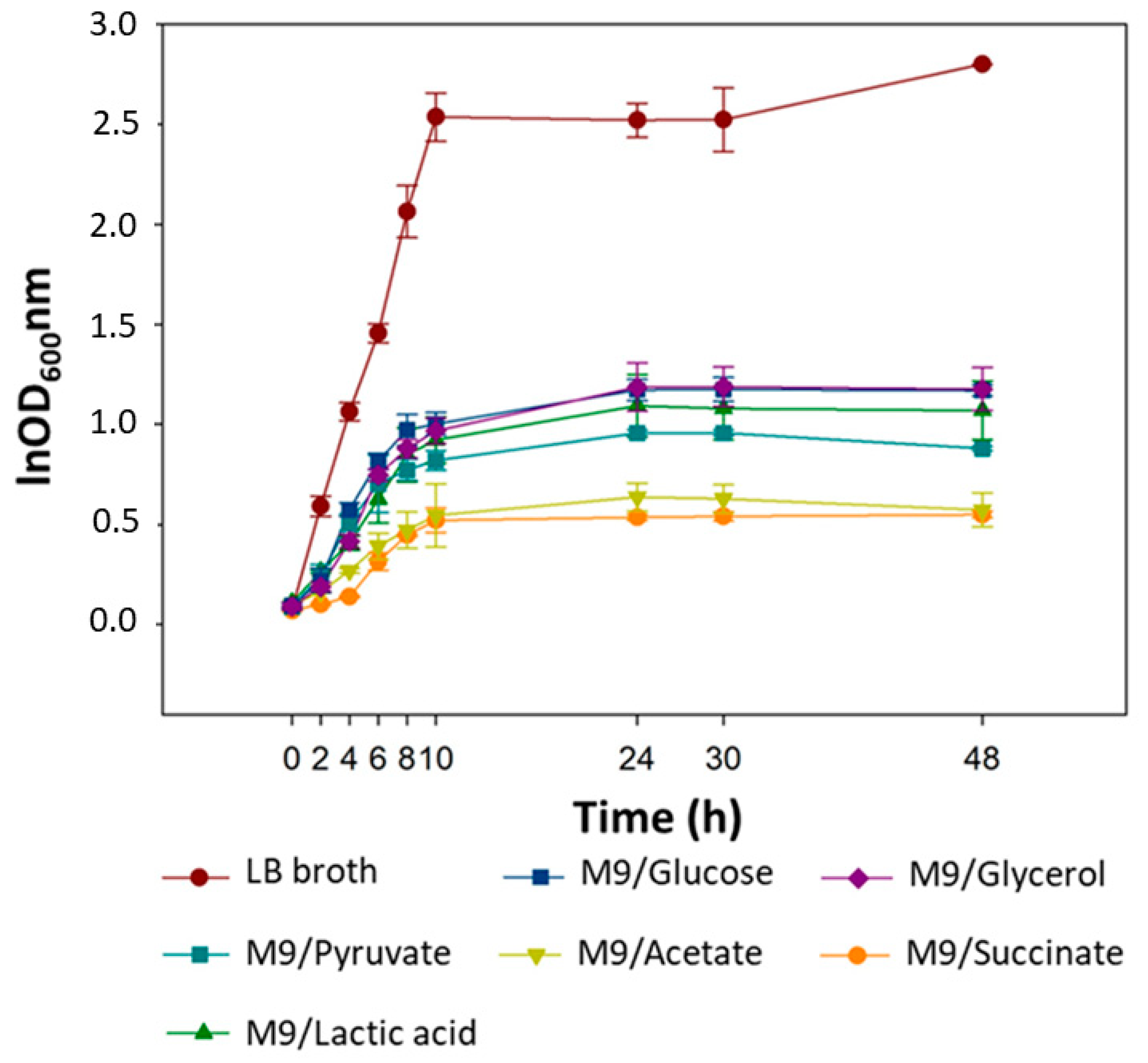
2.1. Bacterial Growth Kinetics
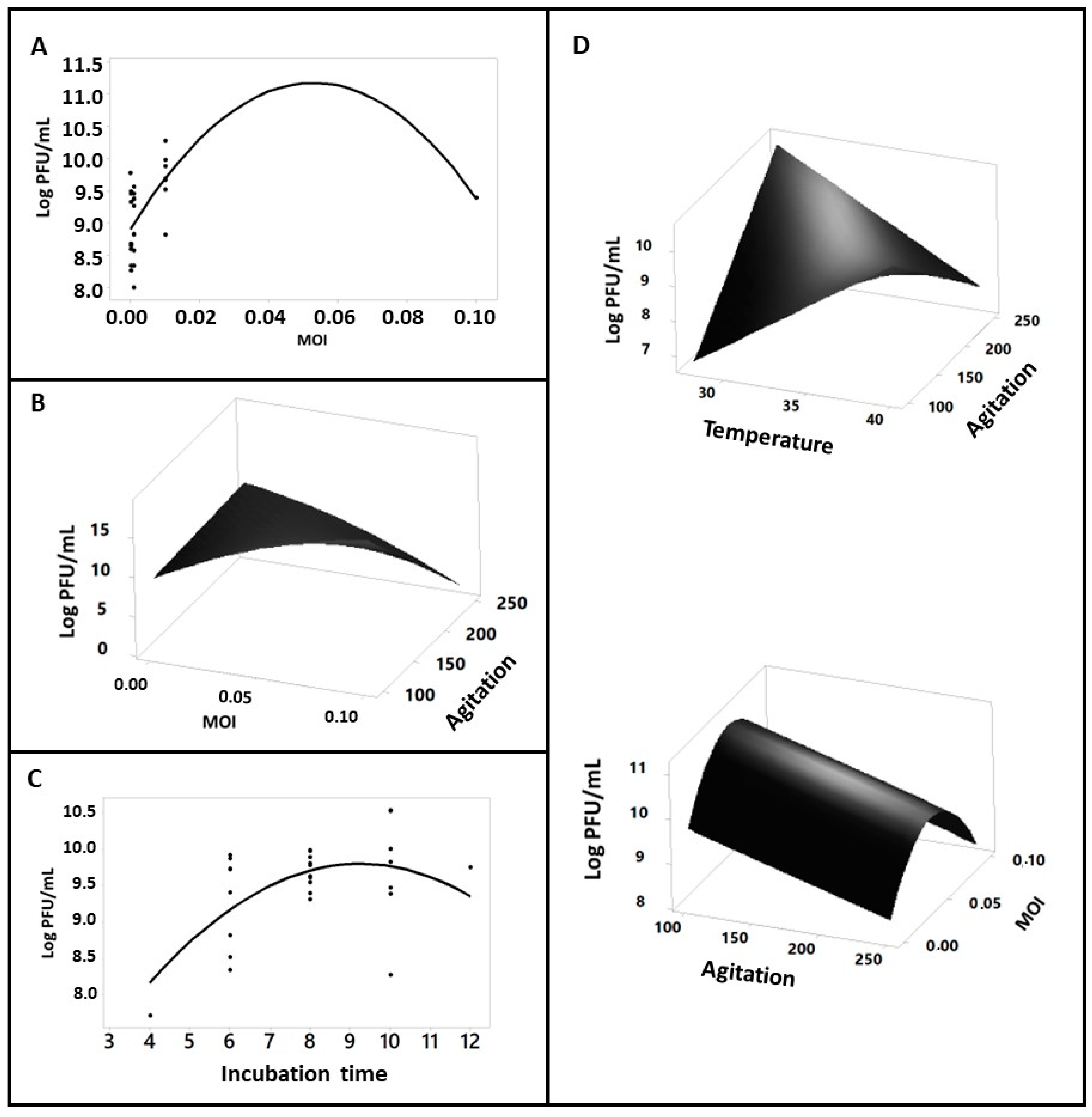
2.2. Impacts of Operational Cultivation Variables on Bacteriophage Production
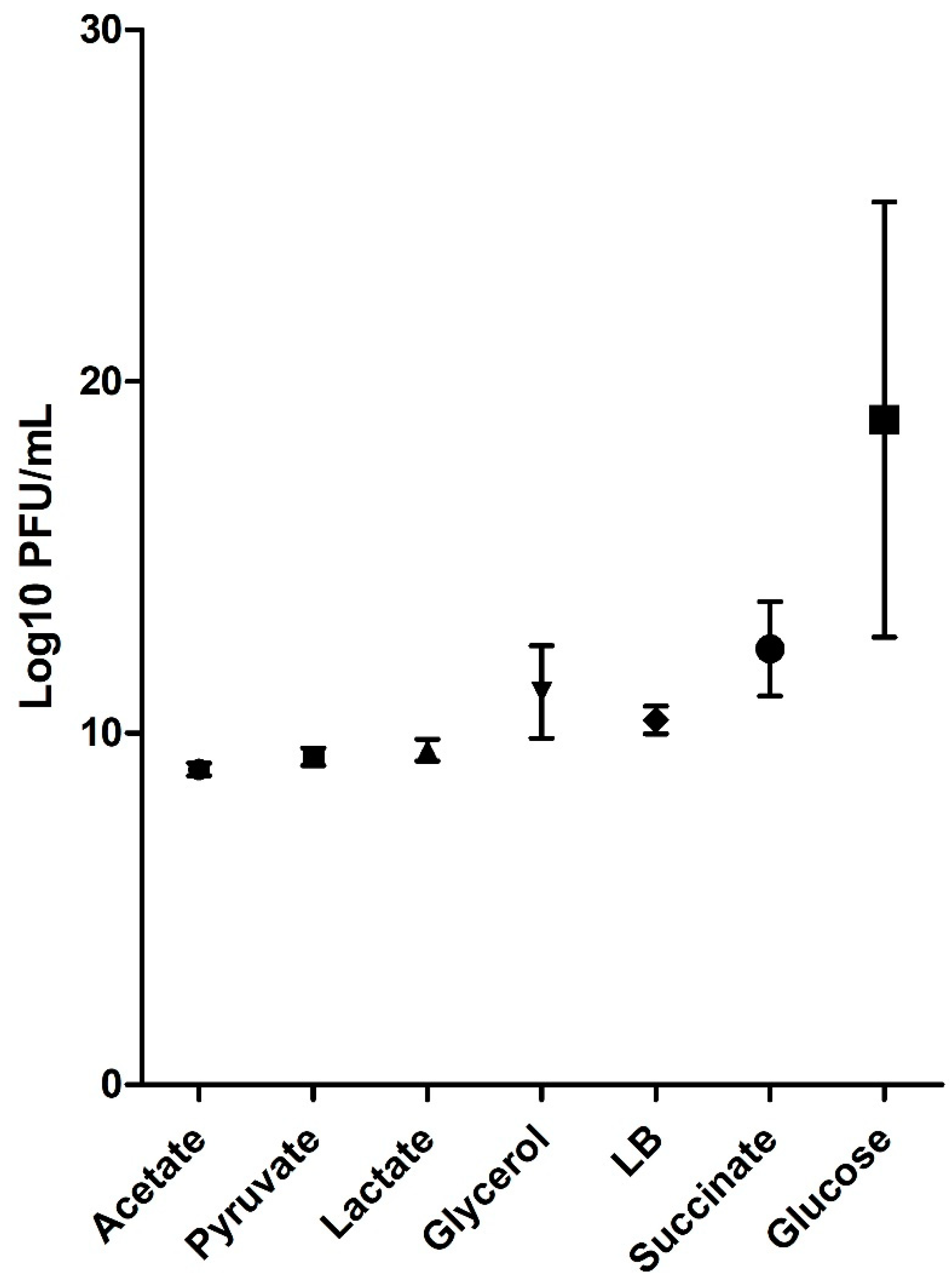
2.3. Prediction of Viral Particle Production in Different Culture Media
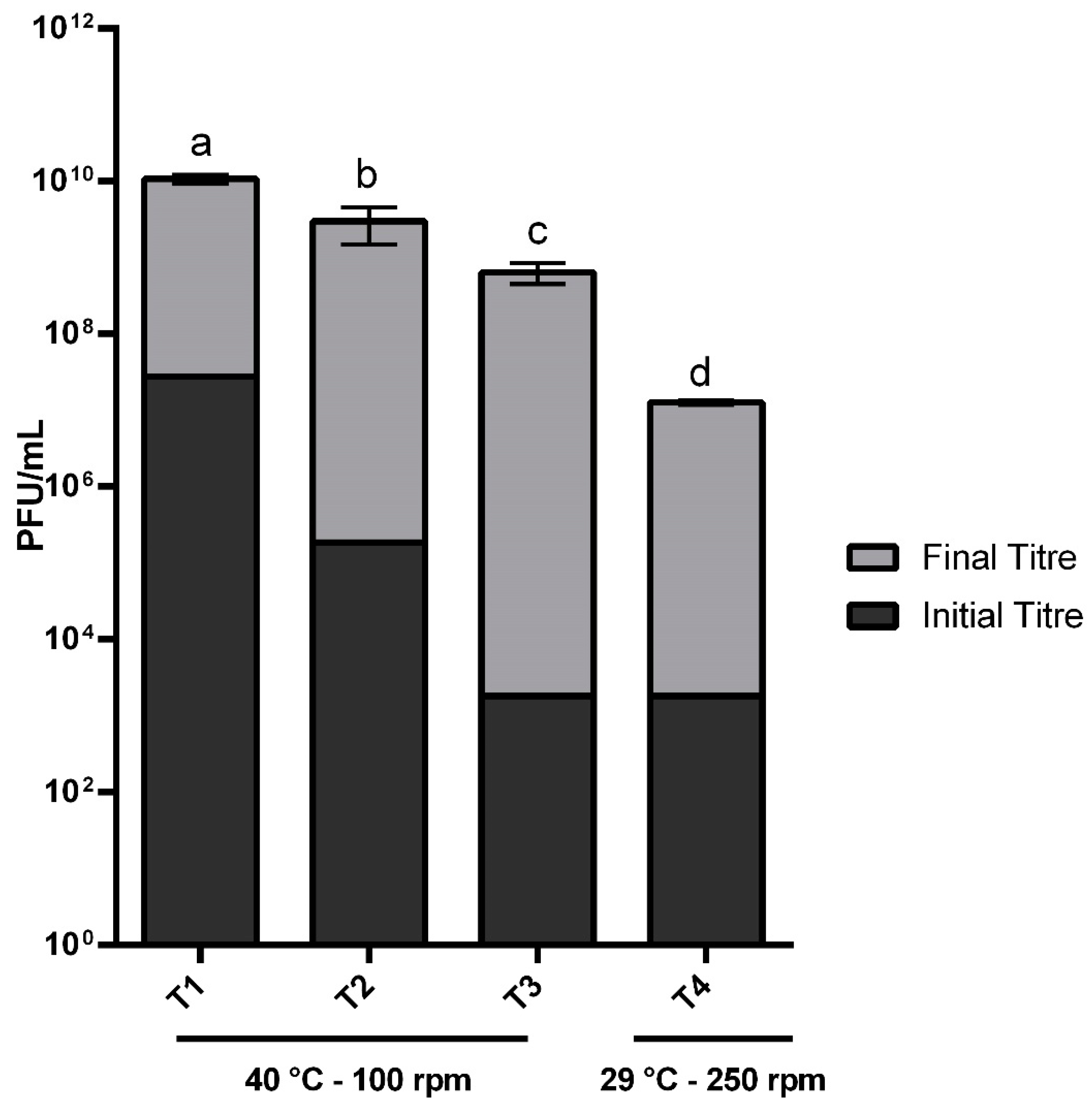
2.4. Validation of The Mathematical Model
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain and vB_EcoM-UFV09 Phage
4.2. M9 Minimal Medium
4.3. Bacterial Growth Kinetics
4.4. Rotatable Central Composite Design (RCCD) and the DOE (Design of Experiment) Optimization Process
4.5. Assembly of RCCD Experiments
4.6. Statistical Analysis and Elaboration of Surface Response Graphs
4.7. Prediction of Viral Particle Production in Different Culture Media
4.8. Model Validation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Golkar, Z.; Bagasra, O.; Gene Pace, D. Bacteriophage therapy: A potential solution for the antibiotic resistance crisis. J. Infect. Dev. Ctries. 2014, 8, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Keen, E.C. Phage therapy: Concept to cure. Front. Microbiol. 2012, 3, 238. [Google Scholar] [CrossRef] [Green Version]
- Pires, D.P.; Costa, A.R.; Pinto, G.; Meneses, L.; Azeredo, J. Current challenges and future opportunities of phage therapy. FEMS Microbiol. Rev. 2020, 44, 684–700. [Google Scholar] [CrossRef]
- Kingwell, K. Bacteriophage therapies re-enter clinical trials. Nat. Publ. Gr. 2015, 14, 515–516. [Google Scholar] [CrossRef]
- Aslam, S.; Lampley, E.; Wooten, D.; Karris, M.; Benson, C.; Strathdee, S.; Schooley, R.T. Lessons learned from the first 10 consecutive cases of intravenous bacteriophage therapy to treat multidrug-resistant bacterial infections at a single center in the United States. Open Forum Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Jault, P.; Leclerc, T.; Jennes, S.; Pirnay, J.P.; Que, Y.A.; Resch, G.; Rousseau, A.F.; Ravat, F.; Carsin, H.; Le Floch, R.; et al. Efficacy and tolerability of a cocktail of bacteriophages to treat burn wounds infected by Pseudomonas aeruginosa (PhagoBurn): A randomised, controlled, double-blind phase 1/2 trial. Lancet Infect. Dis. 2019, 19, 35–45. [Google Scholar] [CrossRef]
- Kutter, E.; De Vos, D.; Gvasalia, G.; Alavidze, Z.; Gogokhia, L.; Kuhl, S.; Abedon, S.T. Phage Therapy in Clinical Practice: Treatment of Human Infections. Curr. Pharm. Biotechnol. 2010, 11, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Fauconnier, A. Phage therapy regulation: From night to dawn. Viruses 2019, 11, 352. [Google Scholar] [CrossRef] [Green Version]
- Pelfrene, E.; Willebrand, E.; Cavaleiro Sanches, A.; Sebris, Z.; Cavaleri, M. Bacteriophage therapy: A regulatory perspective. J. Antimicrob. Chemother. 2016, 71, 2071–2074. [Google Scholar] [CrossRef] [PubMed]
- Clavijo, V.; Baquero, D.; Hernandez, S.; Farfan, J.C.; Arias, J.; Arévalo, A.; Donado-Godoy, P.; Vives-Flores, M. Phage cocktail SalmoFREE® reduces Salmonella on a commercial broiler farm. Poult. Sci. 2019, 98, 5054–5063. [Google Scholar] [CrossRef] [PubMed]
- Duarte, V.S.; Dias, R.S.; Kropinski, A.M.; da Silva Xavier, A.; Ferro, C.G.; Vidigal, P.M.P.; da Silva, C.C.; de Paula, S.O. A T4virus prevents biofilm formation by Trueperella pyogenes. Vet. Microbiol. 2018, 218, 45–51. [Google Scholar] [CrossRef]
- Callaway, T.R.; Edrington, T.S.; Brabban, A.; Kutter, B.; Karriker, L.; Stahl, C.; Wagstrom, E.; Anderson, R.; Poole, T.L.; Genovese, K.; et al. Evaluation of phage treatment as a strategy to reduce Salmonella populations in growing swine. Foodborne Pathog. Dis. 2011, 8, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Bicalho, M.L.S.; Machado, V.S.; Nydam, D.V.; Santos, T.M.A.; Bicalho, R.C. Evaluation of oral administration of bacteriophages to neonatal calves: Phage survival and impact on fecal Escherichia coli. Livest. Sci. 2012, 144, 294–299. [Google Scholar] [CrossRef]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Kumar, C.G.; Anand, S.K. Significance of microbial biofilms in food industry: A review. Int. J. Food Microbiol. 1998, 42, 9–27. [Google Scholar] [CrossRef]
- Costerton, J.W.; Cheng, K.J.; Geesey, G.G.; Ladd, T.I.; Nickel, J.C.; Dasgupta, M.; Marrie, T.J. Bacterial biofilms in nature and disease. Annu. Rev. Microbiol. 1987, 41, 435–464. [Google Scholar] [CrossRef] [PubMed]
- Belgini, D.R.B.; Dias, R.S.; Siqueira, V.M.; Valadares, L.A.B.; Albanese, J.M.; Souza, R.S.; Torres, A.P.R.; Sousa, M.P.; Silva, C.C.; De Paula, S.O.; et al. Culturable bacterial diversity from a feed water of a reverse osmosis system, evaluation of biofilm formation and biocontrol using phages. World J. Microbiol. Biotechnol. 2014, 30, 2689–2700. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.K.; Abedon, S.T. Bacteriophages and their Enzymes in Biofilm Control. Curr. Pharm. Des. 2015, 21, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Knecht, L.E.; Veljkovic, M.; Fieseler, L. Diversity and Function of Phage Encoded Depolymerases. Front. Microbiol. 2020, 10, 2949. [Google Scholar] [CrossRef] [PubMed]
- Beech, I.; Gaylarde, C. Recent advances in the study of biocorrosion - an overview. Rev. Microbiol. 1999, 30, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Koch, G. Cost of Corrosion; Elsevier Ltd.: Amsterdam, The Netherlands, 2017; ISBN 9780081012192. [Google Scholar]
- Koch, G.; Varney, J.; Thopson, N.; Moghissi, O.; Gould, M.; Payer, J. International Measures of Prevention, Application, and Economics of Corrosion Technologies Study; NACE International: Houston, TX, USA, 2016; 216p.
- Enning, D.; Venzlaff, H.; Garrelfs, J.; Dinh, H.T.; Meyer, V.; Mayrhofer, K.; Hassel, A.W.; Stratmann, M.; Widdel, F. Marine sulfate-reducing bacteria cause serious corrosion of iron under electroconductive biogenic. Environ. Microbiol. 2012, 14, 1772–1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gieg, L.M.; Jack, T.R.; Foght, J.M. Biological souring and mitigation in oil reservoirs. Appl. Microbiol. Biotechnol. 2011, 92, 263–282. [Google Scholar] [CrossRef] [PubMed]
- Rabus, R.; Venceslau, S.S.; Wöhlbrand, L.; Voordouw, G.; Wall, J.D.; Pereira, I.A.C. A Post-Genomic View of the Ecophysiology, Catabolism and Biotechnological Relevance of Sulphate-Reducing Prokaryotes. Adv. Microb. Physiol. 2015, 66, 55–321. [Google Scholar] [CrossRef] [PubMed]
- Gieg, L.M.; Davidova, I.A.; Duncan, K.E.; Suflita, J.M. Methanogenesis, sulfate reduction and crude oil biodegradation in hot Alaskan oilfields. Environ. Microbiol. 2010, 12, 3074–3086. [Google Scholar] [CrossRef]
- Grabowski, A.; Nercessian, O.; Fayolle, F.; Blanchet, D.; Jeanthon, C. Microbial diversity in production waters of a low-temperature biodegraded oil reservoir. FEMS Microbiol. Ecol. 2005, 54, 427–443. [Google Scholar] [CrossRef]
- Fraise, A.P. Susceptibility of antibiotic-resistant cocci to biocides. J. Appl. Micriobiology Symp. Suppl. 2002, 92, 158–162. [Google Scholar] [CrossRef]
- Raman, V.; Tamilselvi, S.; Rajendran, N. Evaluation of effective biocides for SRB to control microbiologically influenced corrosion. Mater. Corros. 2008, 044, 329–334. [Google Scholar] [CrossRef]
- Barton, L.L.; Fauque, G.D. Biochemistry, Physiology and Biotechnology of Sulfate-Reducing Bacteria. In Advances in Applied Microbiology; Elsevier Inc.: Amsterdam, The Netherlands, 2009; Volume 68, pp. 41–98. ISBN 9780123748034. [Google Scholar]
- Pires, D.P.; Oliveira, H.; Melo, L.D.R.; Sillankorva, S.; Azeredo, J. Bacteriophage-encoded depolymerases: Their diversity and biotechnological applications. Appl. Microbiol. Biotechnol. 2016, 100, 2141–2151. [Google Scholar] [CrossRef] [Green Version]
- Mohaghegh, A.; Ananda, M.; Bhattacharjee, S.; Goel, R. Biofilm control with natural and genetically-modified phages. World J. Microbiol. Biotechnol. 2016, 34, 67. [Google Scholar] [CrossRef]
- Agboluaje, M.; Sauvageau, D. Bacteriophage Production in Bioreactors. In Bacteriophage Therapy: From Lab to Clinical Practice; Azeredo, J., Sillankorva, S., Eds.; Human Press: Totowa, NJ, USA, 2018; Volume 1693, pp. 173–193. ISBN 9781493973958. [Google Scholar]
- Jurač, K.; Nabergoj, D.; Podgornik, A. Bacteriophage production processes. Appl. Microbiol. Biotechnol. 2019, 103, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Gonzáles-Menéndez, E.; Arroyo-López, F.; Martínez, B.; García, P.; Garrido-Fernandez, A.; Rodríguez, A. Optimizing Propagation of Staphylococcus aureus Infecting Bacteriophage vB _ SauM-phiIPLA-RODI on Surface Methodology. Viruses 2018, 10, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grieco, S.H.H.; Wong, A.Y.K.; Dunbar, W.S.; MacGillivray, R.T.A.; Curtis, S.B. Optimization of fermentation parameters in phage production using response surface methodology. J. Ind. Microbiol. Biotechnol. 2012, 39, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- Grieco, S.-H.H.; Seungil, L.; Dunbar, W.S.; Macgillivray, R.T.A.; Curtis, S.B. Maximizing filamentous phage yield during computer-controlled fermentation. Bioprocess Biosyst. Eng. 2009, 32, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Warner, C.M.; Barker, N.; Perkins, S.L.E.J. M13 bacteriophage production for large-scale applications. Bioprocess Biosyst Eng 2014, 37, 2067–2072. [Google Scholar] [CrossRef]
- Ward, J.M.; Branston, S.; Stanley, E.; Keshavarz-moore, E. Scale-Up and Bioprocessing of Phages. In Bacteriophages—Perspectives and Future; Savva, R., Ed.; IntechOpen: London, UK, 2019; pp. 133–157. [Google Scholar]
- Gutiérrez, D.; Martín-Platero, A.M.; Rodríguez, A.; Martínez-Bueno, M.; García, P.; Martínez, B. Typing of bacteriophages by randomly amplified polymorphic DNA (RAPD)-PCR to assess genetic diversity. FEMS Microbiol. Lett. 2011, 322, 90–97. [Google Scholar] [CrossRef] [Green Version]
- García, R.; Latz, S.; Romero, J.; Higuera, G.; García, K.; Bastías, R. Bacteriophage Production Models: An Overview. Front. Microbiol. 2019, 10, 1187. [Google Scholar] [CrossRef]
- Miller, E.; Kutter, E.; Mosig, G.; Arisaka, F.; Kunisawa, T.; Ruger, W. Bacteriophage T4 Genome. Microbiol. Mol. Biol. Rev. 2003, 67, 86–156. [Google Scholar] [CrossRef] [Green Version]
- Sezonov, G.; Joseleau-Petit, D.; D’Ari, R. Escherichia coli Physiology in Luria-Bertani Broth. J. Bacteriol. 2007, 189, 8746–8749. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Nimtz, M.; Rinas, U. The metabolic potential of Escherichia coli BL21 in defined and rich medium. Microb. Cell Fact. 2014, 13, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Paliy, O.; Gunasekera, T.S. Growth of E. coli BL21 in minimal media with different gluconeogenic carbon sources and salt contents. Appl. Microbiol. Biotechnol. 2007, 1169–1172. [Google Scholar] [CrossRef]
- Sillankorva, S.; Pires, D.; Oliveira, H. The Influence of P. fluorescens Cell Morphology on the Lytic Performance and Production of Phage / IBB-PF7A. Curr. Microbiol. 2011, 63, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Calonico, C.; Delfino, V.; Pesavento, G.; Mundo, M.; Nostro, A. Lo Microbiological Quality of Ready-to-eat Salads from Processing Plant to the Consumers. J. Food Nutr. Res. 2019, 7, 427–434. [Google Scholar] [CrossRef]
- Sillankorva, S.; Oliveira, R.; Vieira, M.J.; Sutherland, I.; Azeredo, J. Pseudomonas fluorescens infection by bacteriophage U S1: The influence of temperature, host growth phase and media. FEMS Microbiol. Lett. 2004, 241, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Padfield, D.; Castledine, M.; Buckling, A. Temperature-dependent changes to host–parasite interactions alter the thermal performance of a bacterial host. ISME J. 2020, 14, 389–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molpeceres, J.; Guzman, M.; Aberturas, M.R.; Chacon, M.; Berges, L. Application of central composite designs to the preparation of polycaprolactone nanoparticles by solvent displacement. J. Pharm. Sci. 1996, 85, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Kollipara, S.; Bende, G.; Movva, S.; Saha, R. Application of rotatable central composite design in the preparation and optimization of poly(lactic-co-glycolic acid) nanoparticles for controlled delivery of paclitaxel. Drug Dev. Ind. Pharm. 2010, 36, 1377–1387. [Google Scholar] [CrossRef] [PubMed]
- Hader, R.J.; Park, S.H. Slope-Rotatable Central Composite Designs. Technometrics 1978, 20, 413–417. [Google Scholar] [CrossRef]
- Marr, A.G. Growth rate of Escherichia coli. Microbiol. Rev. 1991, 55, 316–333. [Google Scholar] [CrossRef]
- Chao, Y.; Patnaik, R.; Roof, W.D.; Young, R.F. Control of Gluconeogenic Growth by pps and pck in Escherichia coli. J. Bacteriol. 1993, 175, 6939–6944. [Google Scholar] [CrossRef] [Green Version]
- Clark, D.W.; Meyer, H.P.; Leist, C.; Fiechter, A. Effects of growth medium on phage production and induction in Escherichia coli K-12 lambda lysogens. J. Biotechnol. 1986, 3, 271–280. [Google Scholar] [CrossRef]
- Reddy, P.; McKenney, K. Improved method for the production of M13 phage and single-stranded DNA for DNA sequencing. Biotechniques 1996, 20, 854–860. [Google Scholar] [CrossRef]
- Choi, C.; Kuatsjah, E.; Wu, E.; Yuan, S. The Effect of Cell Size on the Burst Size of T4 Bacteriophage Infections of Escherichia coli B23. J. Exp. Microbiol. Immunol. 2010, 14, 85–91. [Google Scholar]
- Hadas, H.; Einav, M.; Zaritsky, A. Bacteriophage T4 development depends on the physiology of its host Escherichia coli. J. Bacteriol. 1994, 143, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabinovitch, A.; Hadas, H.; Einav, M.; Melamed, Z.; Zaritsky, A. Model for bacteriophage T4 development in Escherichia coli. J. Bacteriol. 1999, 181, 1677–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger-Munro, M.; Cheung, K.; Fang, A.; Wang, L. T4 Bacteriophage Average Burst Size Varies with Escherichia coli B23 Cell Culture Age. J. Exp. Microbiol. Immunol. 2013, 17, 115–119. [Google Scholar]
- Storms, Z.J.; Brown, T.; Cooper, D.G.; Sauvageau, D.; Leask, R.L. Impact of the cell life-cycle on bacteriophage T4 infection. FEMS Microbiol. Lett. 2014, 353, 63–68. [Google Scholar] [CrossRef]
- Bryan, D.; El-shibiny, A.; Hobbs, Z.; Porter, J.; Kutter, E.M. Bacteriophage T4 Infection of Stationary Phase E. coli: Life after Log from a Phage Perspective. Front. Microbiol. Front. Microbiol. 2016, 7, 1391. [Google Scholar] [CrossRef] [Green Version]
- Golec, P.; Karczewska-Golec, J.; Loś, M.; Wegrzyn, G. Bacteriophage T4 can produce progeny virions in extremely slowly growing Escherichia coli host: Comparison of a mathematical model with the experimental data. FEMS Microbiol. Lett. 2014, 351, 156–161. [Google Scholar] [CrossRef] [Green Version]
- Abedon, S.T.; Hyman, P.; Thomas, C. Experimental Examination of Bacteriophage Latent-Period Evolution as a Response to Bacterial Availability. Appl. Environ. Microbiol. 2003, 69, 7499–7506. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Sun, Q.; Yang, B.; Pan, X.; He, Y.; Yang, H. Quorum sensing influences phage infection efficiency via affecting cell population and physiological state. J. Basic Microbiol. 2017, 57, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Müller-Merbach, M.; Kohler, K.; Hinrichs, J. Environmental factors for phage-induced fermentation problems: Replication and adsorption of the Lactococcus lactis phage P008 as influenced by temperature and pH. Food Microbiol. 2007, 24, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Mojica, K.D.A.; Brussaard, C.P.D. Factors affecting virus dynamics and microbial host-virus interactions in marine environments. FEMS Microbiol. Ecol. 2014, 89, 495–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabig, M.; Herman-Antosiewicz, A.; Kwiatkowska, M.; Los, M.; Thomas, M.S.; Wȩgrzyn, G. The cell surface protein Ag43 facilitates phage infection of Escherichia coli in the presence of bile salts and carborhydrates. Microbiology 2002, 148, 1533–1542. [Google Scholar] [CrossRef] [Green Version]
- Zaburlin, D.; Quiberoni, A.; Mercanti, D. Changes in Environmental Conditions Modify Infection Kinetics of Dairy Phages. Food Environ. Virol. 2017, 9, 270–276. [Google Scholar] [CrossRef]
- Doyle, M.P.; Schoeni, J.L. Survival and growth characteristics of Escherichia coli associated with hemorrhagic colitis. Appl. Environ. Microbiol. 1984, 48, 855–856. [Google Scholar] [CrossRef] [Green Version]
- Wittmann, C.; Becker, J. Oxygen supply in disposable shake-flasks: Prediction of oxygen transfer rate, oxygen saturation and maximum cell concentration during aerobic growth. Biotechnol. Lett. 2013, 35, 1223–1230. [Google Scholar] [CrossRef]
- Storms, Z.J.; Sauvageau, D. Modeling tailed bacteriophage adsorption: Insight into mechanisms. Virology 2015, 485, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Abedon, S.T.; Herschler, T.D.; Stopar, D. Bacteriophage Latent-Period Evolution as a Response to Resource Availability. Appl. Environ. Microbiol. 2001, 67, 4233–4241. [Google Scholar] [CrossRef] [Green Version]
- Boogerd, F.C.; Boe, L.; Michelsen, O.L.E.; Jensen, P.R. atp Mutants of Escherichia coli Fail To Grow on Succinate Due to a Transport Deficiency. J. Bacteriol. 1998, 180, 5855–5859. [Google Scholar] [CrossRef] [Green Version]
- Golais, F.; Hollý, J.; Vítkovská, J. Coevolution of bacteria and their viruses. Folia Microbiol. 2013, 58, 177–186. [Google Scholar] [CrossRef]
- Shapiro, O.H.; Kushmaro, A. Bacteriophage ecology in environmental biotechnology processes. Curr. Opin. Biotechnol. 2011, 22, 449–455. [Google Scholar] [CrossRef]
- Activated, I.; Bacteria, S. Ecology of Bacteriophages. Virology 1991, 57, 2147–2151. [Google Scholar]
- Jover, L.F.; Cortez, M.H.; Weitz, J.S. Mechanisms of multi-strain coexistence in host-phage systems with nested infection networks. J. Theor. Biol. 2013, 332, 65–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choua, M.; Heath, M.R.; Speirs, D.C.; Bonachela, J.A. The effect of viral plasticity on the persistence of host-virus systems. J. Theor. Biol. 2020, 498, 110263. [Google Scholar] [CrossRef] [PubMed]
- Weitz, J.S.; Dushoff, J. Alternative stable states in host - Phage dynamics. Theor. Ecol. 2008, 1, 13–19. [Google Scholar] [CrossRef]
- Hammes, F.; Vital, M.; Egli, T. Critical evaluation of the volumetric “bottle effect” on microbial batch growth. Appl. Environ. Microbiol. 2010, 76, 1278–1281. [Google Scholar] [CrossRef] [Green Version]
- Kram, K.E.; Finkel, S.E. Culture volume and vessel affect long-term survival, mutation frequency, and oxidative stress of Escherichia coli. Appl. Environ. Microbiol. 2014, 80, 1732–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancuso, F.; Shi, J.; Malik, D.J. High throughput manufacturing of bacteriophages using continuous stirred tank bioreactors connected in series to ensure optimum host bacteria physiology for phage production. Viruses 2018, 10, 537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, C.S.; Brown, M.E.; Sama, J.N.; Jackson, M.E.; Dayie, T.K. Growth of wildtype and mutant E. coli strains in minimal media for optimal production of nucleic acids for preparing labeled nucleotides. Appl. Microbiol. Biotechnol. 2010, 88, 771–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium | OD600 Mean (24 h) | µ |
|---|---|---|
| LB | 2.52 | 1.475 |
| M9/Glucose | 1.17 | 0.3737 |
| M9/Glycerol | 1.19 | 0.357 |
| M9/Pyruvate | 0.96 | 0.3485 |
| M9/Succinate | 0.54 | 0.2142 |
| M9/Lactic Acid | 1.09 | 0.1943 |
| M9/Acetate | 0.64 | 0.1763 |
| Coefficients | Acet. | Pyruv. | Gluc. | Glyc. | Succi. | LB |
|---|---|---|---|---|---|---|
| β0 | 9.247 | 11.95 | 11.136 | 11.191 | 10.673 | 10.725 |
| Linear | ||||||
| β1 (Te) | −0.28 | −2.19 | −1.64 | 0.05 | −1.04 | 0.25 |
| β2 (IT) | 0.48 | −0.55 | 1.03 | −0.00 | −0.64 | 1.93 |
| β3 (Ag) | 1.990 | −4.88 | −4.22 * | −3.37 | 0.83 | −3.33 |
| β4 (MOI) | 0.125 | −0.091 | 0.036 | 0.245 | −0.22 | −0.022 |
| Quadratic | ||||||
| β11 (Te2) | −0.797 | −1.254 | −0.039 | −0.158 | −0.268 | −0.129 |
| β22 (IT2) | −0.257 | −0.691 | −0.632 | 0.142 | −0.410 | −1.056 * |
| β33 (Ag2) | 0.209 | −0.392 | −0.042 | 0.040 | −0.360 | −0.135 |
| β44 (MOI2) | −0.293 | −1.358 | −1.347 * | −2.048 * | −1.675 * | −1.075 |
| Interactions | ||||||
| β12 (Te*IT) | 0.687 | 0.473 | −0.090 | 0.577 | 0.173 | −0.481 |
| β13 (Te*Ag) | 0.050 | −1.426 | −0.473 | 0.539 | −1.720 * | −0.099 |
| β14 (Te*MOI) | 0.30 | −2.48 | −2.16 | 0.12 | −1.25 | −0.35 |
| β23 (IT*Ag) | −0.340 | 0.098 | −0.684 | −0.480 | 0.268 | −0.130 |
| β24 (IT*MOI) | 0.25 | −0.96 | 1.10 | 0.00 | −1.15 | 1.44 |
| β34 (Ag*MOI) | 2.00 | −5.75 | −4.52 * | −3.85 | 0.69 | −3.30 |
| R2 adj. (%) | 21.31 | 12.60 | 32.31 | 3.42 | 31.29 | 34.23 |
| Medium | Equations |
|---|---|
| Acetate | |
| Lactate | |
| Pyruvate | |
| Glucose ab | |
| Glycerol a | |
| Succinate ab | |
| LB a |
| Medium | Temperature (Te) | Incubation Time (It) | Agitation (Ag) | Multiplicity of Infection (MOI) |
|---|---|---|---|---|
| Acetate | - | - | - | - |
| Lactate | - | - | - | - |
| Pyruvate | - | - | - | - |
| Glycerol | - | - | - | X |
| Succinate | X | - | X | X |
| Glucose | - | - | X | X |
| LB | - | X | - | - |
| Carbon Source | Price (US$) | Price/Liter * [0.4%] (US$) | Total Price, M9 Medium + Carbon Source |
|---|---|---|---|
| LB Broth, Miller | 169.00/Kg | 4.23 ** | 4.23 ** |
| M9 medium without carbon source | 0.09/L | 0.09 | - |
| Glucose | 7.08/Kg | 0.027 | 0.177 |
| Glycerol | 9.28/L | 0.036 | 0.186 |
| Lactic Acid | 12.57/L | 0.048 | 0.198 |
| Sodium Acetate | 9.66/Kg | 0.038 | 0.188 |
| Sodium Pyruvate | 400/Kg | 1.59 | 1.74 |
| Sodium Succinate | 340.20/Kg | 1.36 | 1.51 |
| Variables | Levels | ||||
|---|---|---|---|---|---|
| −α | −1 | 0 | +1 | +α | |
| Temperature (°C/X1) | 28 | 31 | 34 | 37 | 40 |
| Incubation Time (h/X2) | 4 | 6 | 8 | 10 | 12 |
| Agitation (rpm/X3) | 100 | 138 | 175 | 213 | 250 |
| MOI (PFU/mL/X4) | 0.00001 | 0.0001 | 0.001 | 0.01 | 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, J.; Dias, R.; Junior, J.I.; Marcelino, M.; Silva, M.; Carmo, A.; Sousa, M.; Silva, C.; de Paula, S. A Rapid Method for Performing a Multivariate Optimization of Phage Production Using the RCCD Approach. Pathogens 2021, 10, 1100. https://doi.org/10.3390/pathogens10091100
Silva J, Dias R, Junior JI, Marcelino M, Silva M, Carmo A, Sousa M, Silva C, de Paula S. A Rapid Method for Performing a Multivariate Optimization of Phage Production Using the RCCD Approach. Pathogens. 2021; 10(9):1100. https://doi.org/10.3390/pathogens10091100
Chicago/Turabian StyleSilva, Jessica, Roberto Dias, José Ivo Junior, Maraísa Marcelino, Mirelly Silva, Adriele Carmo, Maira Sousa, Cynthia Silva, and Sergio de Paula. 2021. "A Rapid Method for Performing a Multivariate Optimization of Phage Production Using the RCCD Approach" Pathogens 10, no. 9: 1100. https://doi.org/10.3390/pathogens10091100