
Anti-Inflammatory Properties of Plasma from Children with Short Bowel Syndrome


Abstract
:1. Introduction
2. Results
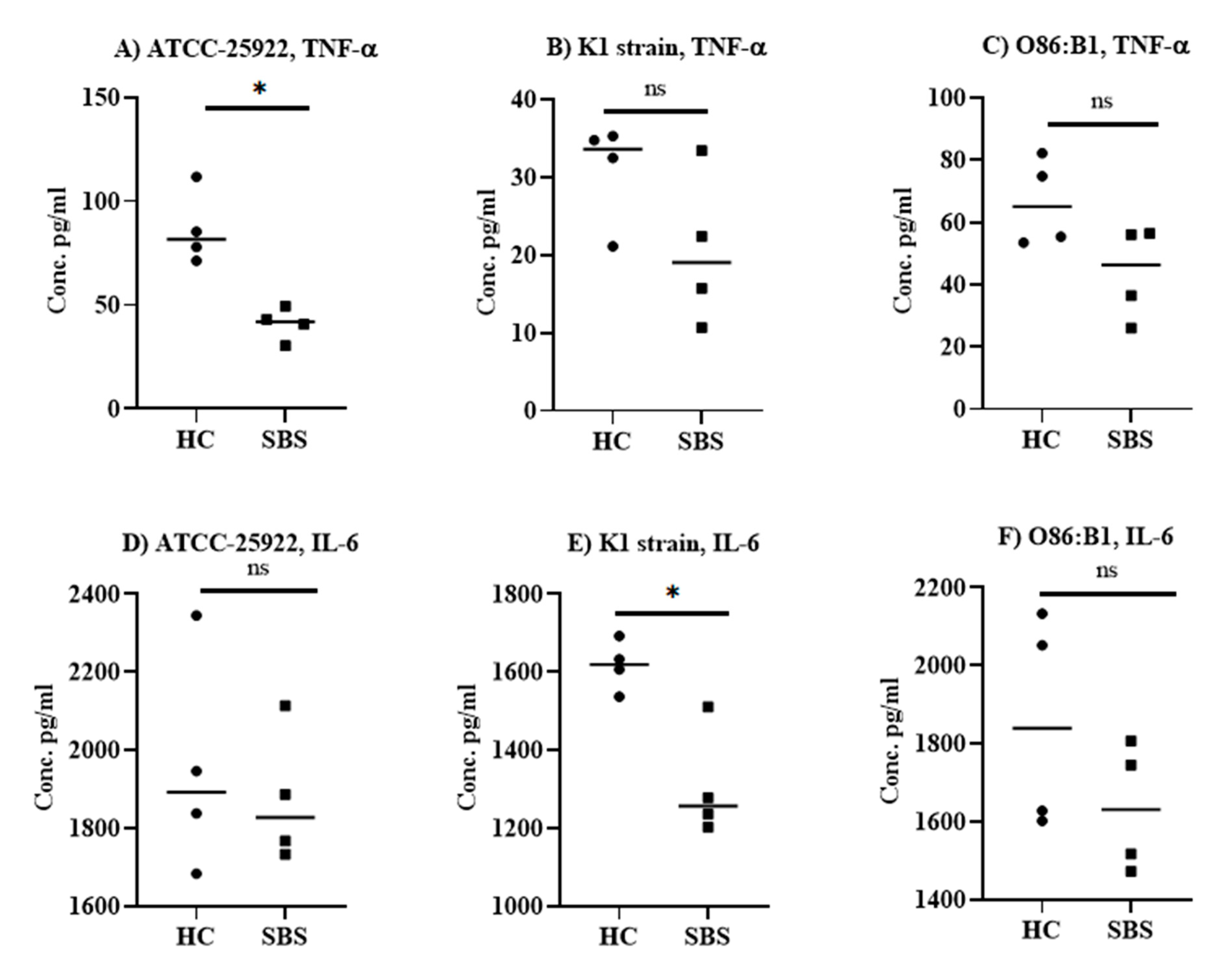
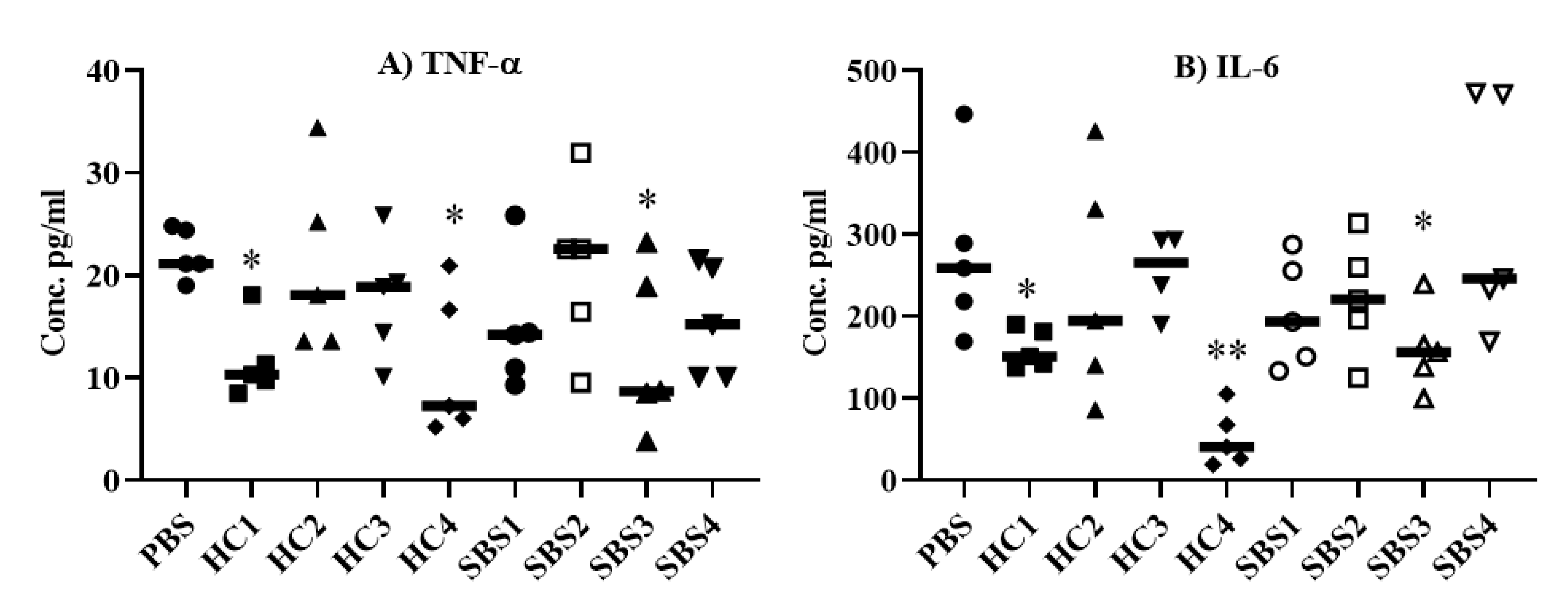
2.1. Anti-Inflammatory Effect of Plasma from SBS and Healthy Subjects In Vitro
2.2. Effect of Plasma from SBS and HC Subjects on Pro-Inflammatory Cytokine Levels in an E. coli Sepsis Murine Model
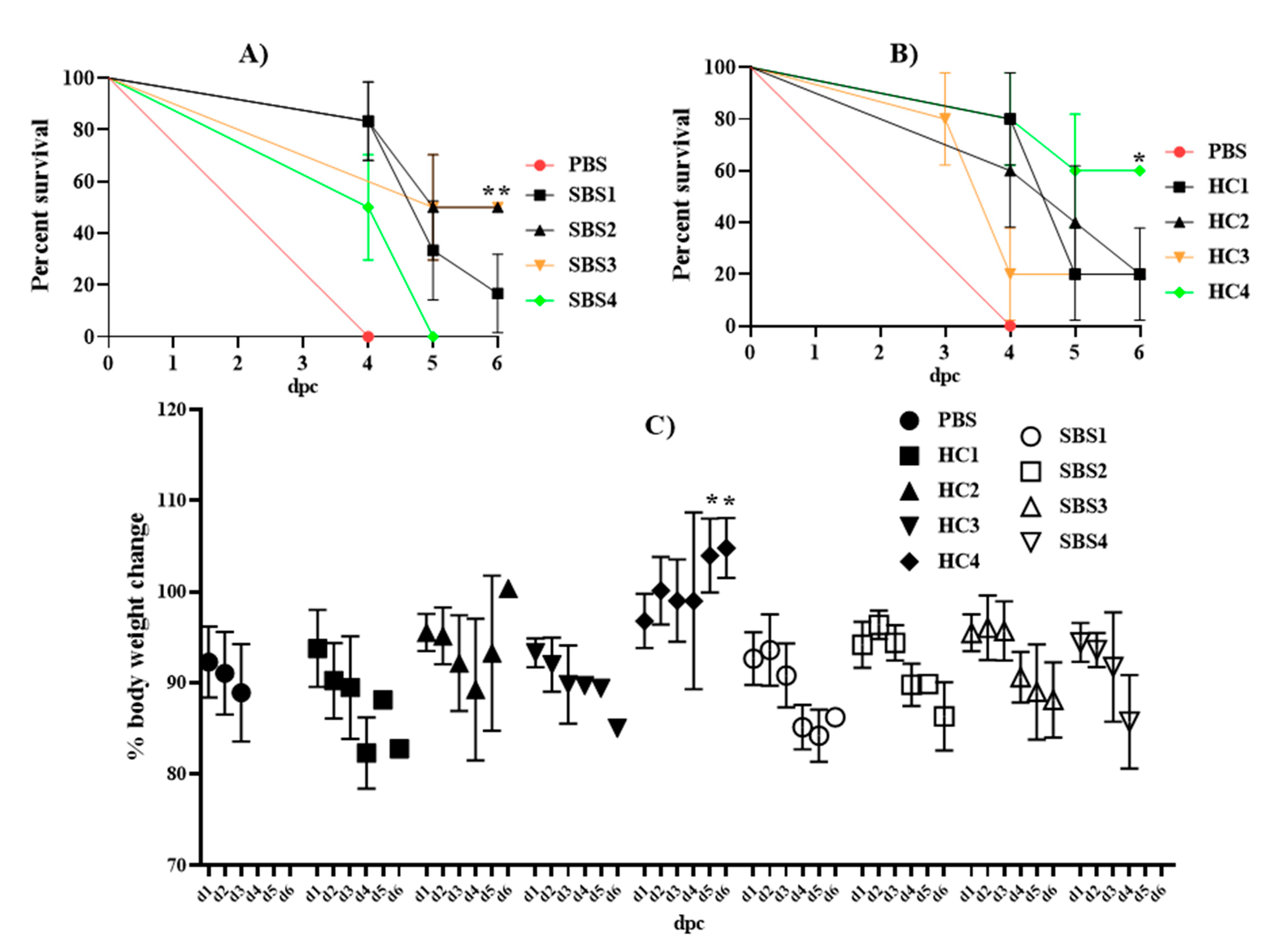
2.3. Effect of Plasma from SBS and HC Subjects on Mortality Induced by E. coli in a Murine Sepsis Model
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Bacterial Strains and Culture Conditions
5.2. In Vitro Studies with Plasma from SBS Subjects
5.3. In Vivo Efficacy of Plasma from SBS Subjects on Sepsis-Induced Mortality
5.4. Cytokine ELISA
5.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vanderhoof, J.A.; Langnas, A.N.; Pinch, L.W.; Thompson, J.S.; Kaufman, S.S. Short bowel syndrome. J. Pediatr. Gastroenterol. Nutr. 1992, 14, 359–370. [Google Scholar] [CrossRef]
- Wales, P.W.; De Silva, N.; Kim, J.H.; Lecce, L.; Sandhu, A.; Moore, A.M. Neonatal short bowel syndrome: A cohort study. J. Pediatr. Surg. 2005, 40, 755–762. [Google Scholar] [CrossRef]
- Goulet, O.; Ruemmele, F. Causes and management of intestinal failure in children. Gastroenterology 2006, 130, S16–S28. [Google Scholar] [CrossRef]
- Vlug, L.E.; Nagelkerke, S.C.J.; Jonkers-Schuitema, C.F.; Rings, E.H.H.M.; Tabbers, M.M. The role of a nutrition support team in the management of intestinal failure patients. Nutrients 2020, 12, 172. [Google Scholar] [CrossRef] [Green Version]
- Wilmore, D.W.; Groff, D.B.; Bishop, H.C.; Dudrick, S.J. Total parenteral nutrition in infants with catastrophic gastrointestinal anomalies. J. Pediatr. Surg. 1969, 4, 181–189. [Google Scholar] [CrossRef]
- Buchman, A.L.; Moukarzel, A.A.; Bhuta, S.; Belle, M.; Ament, M.E.; Eckhert, C.D.; Hollander, D.; Gornbein, J.; Kopple, J.D.; Vijayaroghavan, S.R. Parenteral nutrition is associated with intestinal morphologic and functional changes in humans. J. Parenter. Enter. Nutr. 1995, 19, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Duran, B. The effects of long-term total parenteral nutrition on gut mucosal immunity in children with short bowel syndrome: A systematic review. BMC Nurs. 2005, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, A.; Chiu, C.; Quiros-Tejeira, R.E.; Reyen, L.; Ament, M.; Atkinson, J.B. Fonkalsrud EW. Morbidity and mortality of short-bowel syndrome in infants with abdominal wall defects. Am. Surg. 2002, 68, 75–79. [Google Scholar] [PubMed]
- Zeichner ID, S.L.; Mongodin, E.F.; Hittle, L.; Huang, S.-H.; Torres, C. The bacterial communities of the small intestine and stool in children with short bowel syndrome. PLoS ONE 2019, 14, e0215351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alverdy, J.C.; Aoys, E.; Moss, G.S. Total parenteral nutrition promotes bacterial translocation from the gut. Surgery 1988, 104, 185–190. [Google Scholar] [PubMed]
- O’keefe, S.J.D.; Burnes, J.U.; Thompson, R.L. Recurrent Sepsis in Home Parenteral Nutrition Patients: An Analysis of Risk Factors. J. Parenter. Enter. Nutr. 1994, 18, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Alverdy, J.; Chi, H.S.; Sheldon, G.F. The effect of parenteral nutrition on gastrointestinal immunity. The importance of enteral stimulation. Ann. Surg. 1985, 202, 681–684. [Google Scholar] [CrossRef]
- D’Antiga, L.; Dhawan, A.; Davenport, M.; Mieli-Vergani, G.; Bjarnason, I. Intestinal absorption and permeability in paediatric short-bowel syndrome: A pilot study. J. Pediatr. Gastroenterol. Nutr. 1999, 29, 588–593. [Google Scholar] [CrossRef] [PubMed]
- McAndrew, H.F.; Lloyd, D.A.; Rintala, R.; Van Saene, H.K.F. The effects of intravenous epidermal growth factor on bacterial translocation and central venons catheter infection in the rat total parenteral nutrition model. Pediatr. Surg. Int. 2000, 16, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Bruewer, M.; Luegering, A.; Kucharzik, T.; Parkos, C.A.; Madara, J.L.; Hopkins, A.M.; Nusrat, A. Proinflammatory Cytokines Disrupt Epithelial Barrier Function by Apoptosis-Independent Mechanisms. J. Immunol. 2003, 171, 6164–6172. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, T.R.; Evans, M.E.; Fernández-Estívariz, C.; Jones, D.P. Trophic and cytoprotective nutrition for intestinal adaptation, mucosal repair, and barrier function. Annu. Rev. Nutr. 2003, 23, 229–261. [Google Scholar] [CrossRef]
- Briet, F.; Flourié, B.; Achour, L.; Maurel, M.; Rambaud, J.C.; Messing, B. Bacterial adaptation in patients with short bowel and colon in continuity. Gastroenterology 1995, 109, 1446–1453. [Google Scholar] [CrossRef]
- Buchman, A.L.; Moukarzel, A.; Goodson, B.; Herzog, F.; Pollack, P.; Reyen, L.; Alvarez, M.; Ament, M.E.; Gornbein, J. Catheter-related infections associated with home parenteral nutrition and predictive factors for the need for catheter removal in their treatment. J. Parenter. Enter. Nutr. 1994, 18, 297–302. [Google Scholar] [CrossRef]
- Ziegler, T.R.; Smith, R.J.; O’dwyer, S.T.; Demling, R.H.; Wilmore, D. Increased Intestinal Permeability Associated With Infection in Burn Patients. Arch. Surg. 1988, 123, 1313–1319. [Google Scholar] [CrossRef]
- O’Boyle, C.J.; MacFie, J.; Mitchell, C.J.; Johnstone, D.; Sagar, P.M.; Sedman, P.C. Microbiology of bacterial translocation in humans. Gut 1998, 42, 29–35. [Google Scholar] [CrossRef]
- Ziegler, T.R.; Luo, M.; Estívariz, C.F.; Moore, D.A.; Sitaraman, S.V.; Hao, L.; Bazargan, N.; Klapproth, J.M.; Tian, J.; Galloway, J.R.; et al. Detectable serum flagellin and lipopolysaccharide and upregulated anti-flagellin and lipopolysaccharide immunoglobulins in human short bowel syndrome. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R402. [Google Scholar] [CrossRef]
- Opal, S.M. Endotoxins and other sepsis triggers. Contrib. Nephrol. 2010, 167, 14–24. [Google Scholar] [CrossRef]
- Angus, D.C.; van der Poll, T. Severe Sepsis and Septic Shock. N. Engl. J. Med. 2013, 369, 840–851. [Google Scholar] [CrossRef]
- Thamphiwatana, S.; Angsantikul, P.; Escajadillo, T.; Zhang, Q.; Olson, J.; Luk, B.T.; Zhang, S.; Fang, R.H.; Gao, W.; Nizet, V.; et al. Macrophage-like nanoparticles concurrently absorbing endotoxins and proinflammatory cytokines for sepsis management. Proc. Natl. Acad. Sci. USA 2017, 114, 11488–11493. [Google Scholar] [CrossRef] [Green Version]
- Cirioni, O.; Giacometti, A.; Ghiselli, R.; Bergnach, C.; Orlando, F.; Silvestri, C.; Mocchegiani, F.; Licci, A.; Skerlavaj, B.; Rocchi, M.; et al. LL-37 protects rats against lethal sepsis caused by gram-negative bacteria. Antimicrob. Agents Chemother. 2006, 50, 1672–1679. [Google Scholar] [CrossRef] [Green Version]
- Mayeux, P.R. Pathobiology of lipopolysaccharide. J. Toxicol. Environ. Health 1997, 51, 415–435. [Google Scholar] [CrossRef] [PubMed]
- Cole, C.R.; Frem, J.C.; Schmotzer, B.; Gewirtz, A.T.; Meddings, J.B.; Gold, B.D.; Ziegler, T.R. The Rate of Bloodstream Infection Is High in Infants with Short Bowel Syndrome: Relationship with Small Bowel Bacterial Overgrowth, Enteral Feeding, and Inflammatory and Immune Responses. J. Pediatr. 2010, 156, 941–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahunja, K.M.; Ahmed, T.; Hossain, M.I.; Islam, M.M.; Monjory, M.B.; Shahid, A.S.M.S.B.; Faruque, A.S.G.; Chisti, M.J. Clinical and laboratory characteristics of children under five hospitalized with diarrhea and bacteremia. PLoS ONE 2020, 15, e0243128. [Google Scholar] [CrossRef]
- Chen, S.; Liu, S.; Yuan, X.; Mai, H.; Lin, J.; Wen, F. Etiology, drug sensitivity profiles and clinical outcome of bloodstream infections: A retrospective study of 784 pediatric patients with hematological and neoplastic diseases. Pediatric Hematol. Oncol. 2019, 36, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Galloway, D.P.; Troutt, M.L.; Kocoshis, S.A.; Gewirtz, A.T.; Ziegler, T.R.; Cole, C.R. Increased Anti-Flagellin and Anti-Lipopolysaccharide Immunoglobulins in Pediatric Intestinal Failure: Associations With Fever and Central Line-Associated Bloodstream Infections. JPEN. J. Parenter. Enteral Nutr. 2015, 39, 562–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kono, K.; Sekikawa, T.; Iizuka, H.; Ichihara, F.; Amemiya, H.; Ishikawa, T.; Matsumoto, Y. T-Cell Dysfunction in a Patient with Short Bowel Syndrome: Report of a Case. Surg. Today. 1999, 29, 1253–1256. [Google Scholar] [CrossRef]
- Kasten, K.R.; Tschöp, J.; Adediran, S.G.; Hildeman, D.A.; Caldwell, C.C. T cells are potent early mediators of the host response to sepsis. Shock 2010, 34, 327–336. [Google Scholar] [CrossRef]
- Steinhagen, F.; Schmidt, S.V.; Schewe, J.C.; Peukert, K.; Klinman, D.M.; Bode, C. Immunotherapy in sepsis—Brake or accelerate? Pharmacol. Ther. 2020, 208, 107476. [Google Scholar] [CrossRef] [PubMed]
- Shankar-Hari, M.; Spencer, J.; Sewell, W.A.; Rowan, K.M.; Singer, M. Bench-to-bedside review: Immunoglobulin therapy for sepsis—Biological plausibility from a critical care perspective. Crit. Care 2011, 16, 206. [Google Scholar] [CrossRef] [Green Version]
- Di Rosa, R.; Pietrosanti, M.; Luzi, G.; Salemi, S.; D’Amelio, R. Polyclonal intravenous immunoglobulin: An important additional strategy in sepsis? Eur. J. Intern. Med. 2014, 25, 511–516. [Google Scholar] [CrossRef]
- Alejandria, M.M.; Lansang, M.A.D.; Dans, L.F.; Mantaring, J.B. Intravenous immunoglobulin for treating sepsis, severe sepsis and septic shock. Cochrane Database Syst. Rev. 2013, 2013, CD001090. [Google Scholar] [CrossRef]
- Almansa, R.; Tamayo, E.; Andaluz-Ojeda, D.; Nogales, L.; Blanco, J.; Eiros, M.M.; Gomez-Herreras, J.I.; Bermejo-Martin, J.F. The original sins of clinical trials with intravenous immunoglobulins in sepsis. Crit. Care 2015, 19, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, M.Y.; Cisalpino, D.; Varadarajan, S.; Hellman, J.; Warren, H.S.; Cascalho, M.; Inohara, N.; Núñez, G. Gut Microbiota-Induced Immunoglobulin G Controls Systemic Infection by Symbiotic Bacteria and Pathogens. Immunity 2016, 44, 647–658. [Google Scholar] [CrossRef] [Green Version]
- Speer, E.M.; Diago-Navarro, E.; Ozog, L.S.; Raheel, M.; Levy, O.; Fries, B.C. A Neonatal Murine Escherichia coli Sepsis Model Demonstrates That Adjunctive Pentoxifylline Enhances the Ratio of Anti- vs. Pro-inflammatory Cytokines in Blood and Organ Tissues. Front. Immunol. 2020, 11, 2249. [Google Scholar] [CrossRef] [PubMed]
- Maric, S.; Flüchter, P.; Guglielmetti, L.C.; Staerkle, R.F.; Sasse, T.; Restin, T.; Schneider, C.; Holland-Cunz, S.G.; Crenn, P.; Vuille-Dit-Bille, R.N. Plasma citrulline correlates with basolateral amino acid transporter LAT4 expression in human small intestine. Clin. Nutr. 2021, 40, 2244–2251. [Google Scholar] [CrossRef] [PubMed]
- Pabisek-Miernik, J.; Kościelniak-Merak, B.; Sporadyk, M.; Tomasik, P.J. Ghrelin, visfatin and irisin in children with short bowel syndrome. Folia Med. Cracov. 2019, 59, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Turato, W.M.; Sales-Campos, H.; Braga, C.B.M.; Cunha, S.F.C.; Silvah, J.H.; da Silva, J.S.; Marchini, J.S.; de Barros Cardoso, C.R. The impact of intestinal resection on the immune function of short bowel syndrome patients. Hum. Immunol. 2016, 77, 1202–1208. [Google Scholar] [CrossRef] [PubMed]
- Metin, M.; Altun, A.; Köylüoğlu, G. The effect of probiotics on ıntestinal motility in an experimental short bowel model. Acta Cir. Bras. 2020, 35, 1–14. [Google Scholar] [CrossRef]
- Santacroce, L.; Man, A.; Charitos, I.A.; Haxhirexha, K.; Topi, S. Current knowledge about the connection between health status and gut microbiota from birth to elderly. A narrative review. Front. Biosci. 2021, 26, 135. [Google Scholar] [CrossRef]
- Ballini, A.; Gnoni, A.; De Vito, D.; Dipalma, G.; Cantore, S.; Gargiulo Isacco, C.; Saini, R.; Santacroce, L.; Topi, S.; Scarano, A.; et al. Effect of probiotics on the occurrence of nutrition absorption capacities in healthy children: A randomized double-blinded placebo-controlled pilot study. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8645–8657. [Google Scholar] [CrossRef] [PubMed]
- Asha, M.Z.; Khalil, S.F.H. Efficacy and Safety of Probiotics, Prebiotics and Synbiotics in the Treatment of Irritable Bowel Syndrome: A systematic review and meta-analysis. Sultan Qaboos Univ. Med. J. 2020, 20, e13–e24. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Subject 1 | Subject 2 | Subject 3 | Subject 4 |
|---|---|---|---|---|
| Age (years) | 5 | 9 | 14 | 9 |
| Sex | F | M | F | F |
| Diagnosis | Malrotation, Midgut volvulus, Jejunal atresia | Necrotizing enterocolitis, Jejunal perforations | Large and small bowel atresia, Persistent omphalovitelline duct | Gastroschisis |
| Remaining Small Bowel Length | Unknown. Noted to have resection of 3 cm jejunum, 6 cm ileum | 63.6 cm | 41 cm | 45 cm |
| PN a dependence | 9% | 14% | 72% | 100% |
| Current Central Line (Duration) | Broviac (59 months) | Port-a-Cath (12 months) | Broviac (13 months) | Broviac (39 months) |
| Number of Line Infections b | 0 | 2 | 1 | 21 |
| Time Since Last CLABSI, past organisms Isolated | NA | 8 years Lactobacillus | 7 years corynebacterium spp | 1 month E. coli, K.oxytoca, S.epidermidis, S.haemolyticus, S. hominis, S.gallolyticus, E. faecalis |
| Antibiotic Therapy for SIBO c or Prokinetic Effects | Yes | Yes | Yes | No |
| Days from Last Antibiotic Exposure | 7 | 14 | 60 | 35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hajam, I.A.; Ali, F.; Young, J.; Garcia, M.A.; Cannavino, C.; Ramchandar, N.; Liu, G.Y. Anti-Inflammatory Properties of Plasma from Children with Short Bowel Syndrome. Pathogens 2021, 10, 1021. https://doi.org/10.3390/pathogens10081021
Hajam IA, Ali F, Young J, Garcia MA, Cannavino C, Ramchandar N, Liu GY. Anti-Inflammatory Properties of Plasma from Children with Short Bowel Syndrome. Pathogens. 2021; 10(8):1021. https://doi.org/10.3390/pathogens10081021
Chicago/Turabian StyleHajam, Irshad Ahmed, Farhana Ali, Jocelyn Young, Mary Abigail Garcia, Christopher Cannavino, Nanda Ramchandar, and George Y. Liu. 2021. "Anti-Inflammatory Properties of Plasma from Children with Short Bowel Syndrome" Pathogens 10, no. 8: 1021. https://doi.org/10.3390/pathogens10081021