
A Comparison of Virulence of Three Porcine Circovirus Type 2 (PCV2) Genotypes (a, b, and d) in Pigs Singularly Inoculated with PCV2 and Dually Inoculated with PCV2 and Porcine Reproductive and Respiratory Syndrome Virus


Abstract
:1. Introduction
2. Results
2.1. Clinical Signs
2.2. Growth Performance
2.3. Enzyme-Linked Immunosorbent Assay
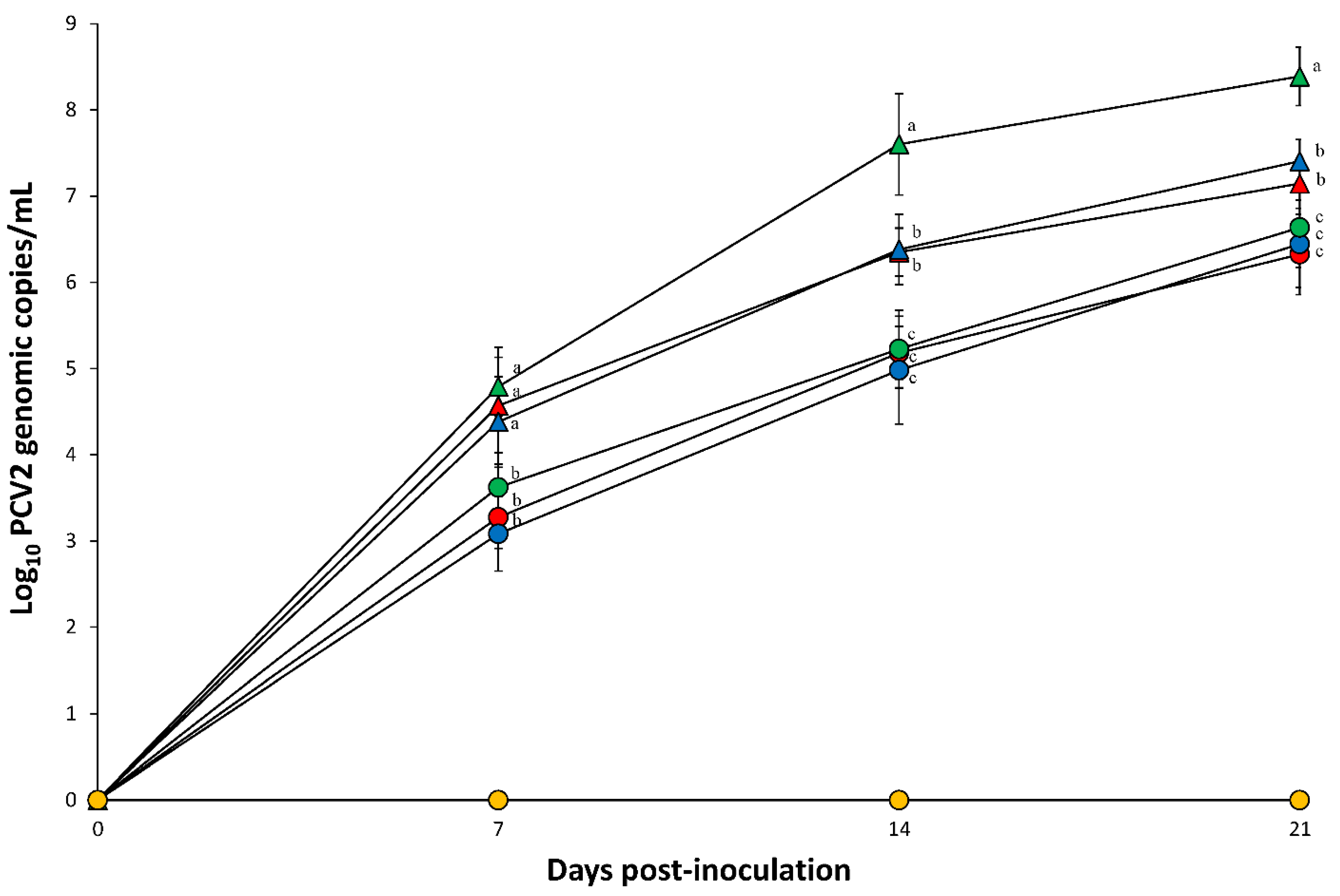
2.4. Quantification of PCV2 DNA in the Blood
2.5. Quantification of PRRSV cDNA in the Blood
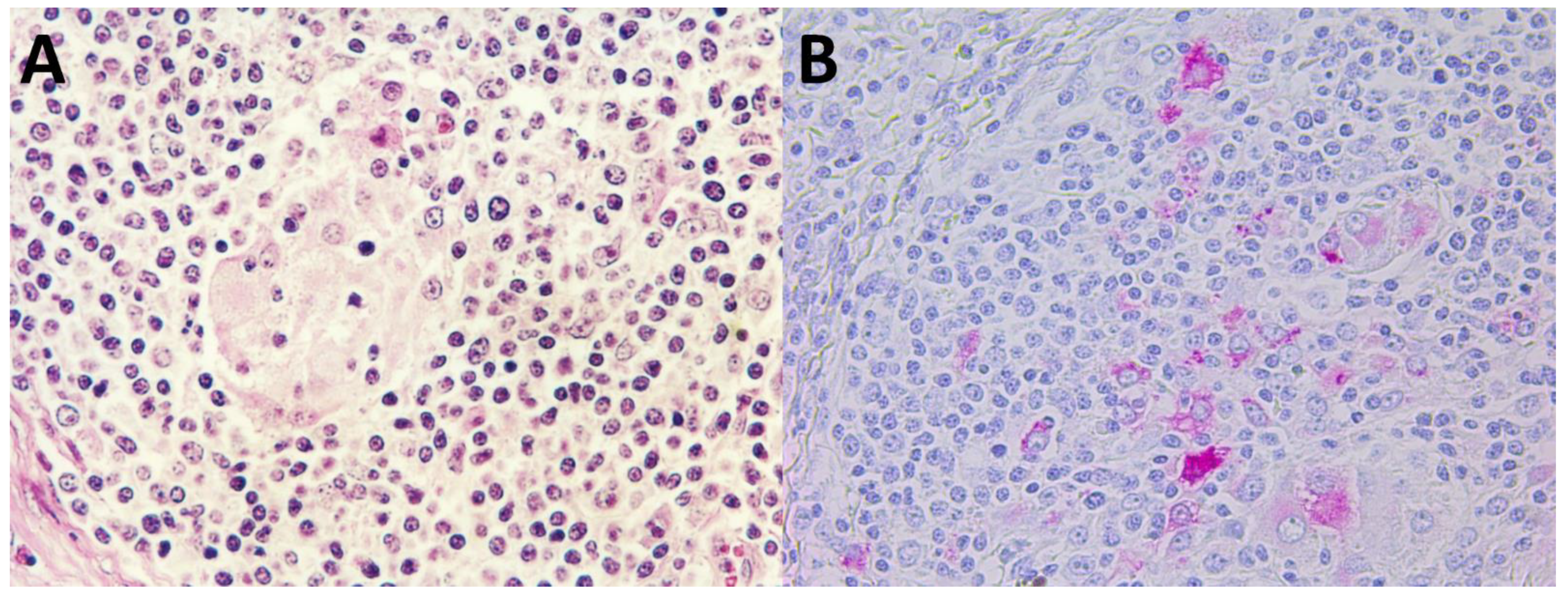
2.6. Histopathology
2.7. Immunohistochemistry
2.8. Correlation between PCV2 Viremia and Lymphoid Lesions
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design
4.3. Clinical Observation
4.4. Growth Performance
4.5. Quantification of PCV2 DNA in the Blood
4.6. Quantification of PRRSV cDNA in the Blood
4.7. Serology
4.8. Histopathology
4.9. Immunohistochemistry
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rosario, K.; Breitbart, M.; Harrach, B.; Segales, J.; Delwar, E.; Biagini, P.; Varsani, A. Revisiting the taxonomy of the family Circoviridae: Establishment of the genus Cyclovirus and removal of the genus Gyrovirus. Arch. Virol. 2017, 162, 1447–1463. [Google Scholar] [CrossRef] [Green Version]
- Franzo, G.; Segalés, J. Porcine circovirus 2 (PCV-2) genotype update and proposal of a new genotyping methodology. PLoS ONE 2018, 13, e0208585. [Google Scholar] [CrossRef] [Green Version]
- Chae, C. A review of porcine circovirus 2-associated syndromes and diseases. Vet. J. 2005, 169, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Ratification vote on taxonomic proposals to the international committee on taxonomy of viruses. Arch. Virol. 2016, 161, 2921–2924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmerman, J.J.; Benfield, D.A.; Dee, S.A.; Murtaugh, M.P.; Stadejek, T.; Stevenson, G.W.; Torremorell, M. Porcine Reproductive and Respiratory Syndrome Virus (porcine arterivirus). In Diseases of Swine, 10th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Eds.; Wiley-Blackwell: Ames, IA, USA, 2012; pp. 461–486. [Google Scholar]
- Xiao, C.T.; Harmon, K.M.; Halbur, P.G.; Opriessnig, T. PCV2d-2 is the predominant type of PCV2 DNA in pig samples collected in the U.S. during 2014-2016. Vet. Microbiol. 2016, 197, 72–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, C.T.; Halbur, P.G.; Opriessnig, T. Global molecular genetic analysis of porcine circovirus type 2 (PCV2) sequences confirms the presence of four main PCV2 genotypes and reveals a rapid increase of PCV2d. J. Gen. Virol. 2015, 96, 1830–1841. [Google Scholar] [CrossRef]
- Kwon, T.; Lee, D.-U.; Yoo, S.J.; Je, S.H.; Shin, J.Y.; Lyoo, Y.S. Genotypic diversity of porcine circovirus type 2 (PCV2) and genotype shift to PCV2d in Korean pig population. Virus Res. 2017, 228, 24–29. [Google Scholar] [CrossRef]
- Cho, H.; Kang, I.; Oh, T.; Yang, S.; Park, K.H.; Min, K.-D.; Ham, H.J.; Chae, C. Comparative study of the virulence of 3 major Korean porcine circovirus type 2 genotypes (a, b, and d). Can. J. Vet. Res. 2020, 44, 235–240. [Google Scholar]
- Guo, L.; Fu, Y.; Wang, Y.; Lu, Y.; Wei, Y.; Tang, Q.; Fan, P.; Liu, J.; Zhang, L.; Zhang, F.; et al. A porcine circovirus type 2 (PCV2) mutant with 234 amino acids in capsid protein showed more virulence in vivo, compared with classical PCV2a /b strain. PLoS ONE 2012, 7, e41463. [Google Scholar] [CrossRef] [Green Version]
- Opriessnig, T.; Xiao, C.T.; Gerber, P.F.; Halbur, P.G.; Matzinger, S.R.; Meng, X.J. Mutant USA strain of porcine circovirus type 2 (mPCV2) exhibits similar virulence to the classical PCV2a and PCV2b strains in caesarean-derived, colostrum-deprived pigs. J. Gen. Virol. 2014, 95, 2495–2503. [Google Scholar] [CrossRef] [Green Version]
- Drolet, R.; Larochelle, R.; Morin, M.; Delisle, B.; Magar, R. Detection rates of porcine reproductive and respiratory syndrome virus, porcine circovirus type 2, and swine influenza virus in porcine proliferative and necrotizing pneumonia. Vet. Pathol. 2003, 40, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Harms, P.A.; Sorden, S.D.; Halbur, P.G.; Bolin, S.R.; Lager, K.M.; Morozov, I.; Paul, P.S. Experimental reproduction of severe disease in CD/CD pigs concurrently infected with type 2 porcine circovirus and porcine reproductive and respiratory syndrome virus. Vet. Pathol. 2001, 38, 528–539. [Google Scholar] [CrossRef]
- Opriessnig, T.; Madson, D.M.; Prickett, J.R.; Kuhar, D.; Lunney, J.K.; Elsener, J.; Halbur, P.G. Effect of porcine circovirus type 2 (PCV2) vaccination on porcine reproductive and respiratory syndrome virus (PRRSV) and PCV2 coinfection. Vet. Microbiol. 2008, 131, 103–114. [Google Scholar] [CrossRef]
- Rovira, A.; Balasch, M.; Segalés, J.; Garcia, L.; Plana-Dúran, J.; Rosell, C.; Ellerbrok, H.; Mankertz, A.; Domingo, M. Experimental inoculation of conventional pigs with porcine reproductive and respiratory syndrome virus and porcine circovirus 2. J. Virol. 2002, 76, 3232–3239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.; Liu, M.; Liu, Z.; Meng, F.; Wang, H.; Cao, L.; Jiao, Q.; Han, Z.; Liu, S. Epidemiological investigation of porcine circovirus type 2 and its coinfection rate in Shandong province in China from 2015 to 2018. BMC Vet. Res. 2021, 17, 17. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, T.; Zhang, X.; Liu, X.; Ren, L. Co-infection of swine with porcine circovirus type 2 and other swine viruses. Viruses 2019, 11, 185. [Google Scholar] [CrossRef] [Green Version]
- Opriessnig, T.; Gauger, P.C.; Faaberg, K.S.; Shen, H.; Beach, N.M.; Meng, X.-J.; Wang, C.; Halbur, P.G. Effect of porcine circovirus type 2a or 2b on infection kinetics and pathogenicity of two genetically divergent strains of porcine reproductive and respiratory syndrome virus in the conventional pig model. Vet. Microbiol. 2012, 158, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Park, C.; Seo, H.W.; Park, S.-J.; Han, K.; Chae, C. Comparison of porcine corcovirus type 2 (PCV2)-associated lesions produced by co-infection between two genotypes of PCV2 and two genotypes of porcine reproductive and respiratory syndrome virus. J. Gen. Virol. 2014, 95, 2486–2494. [Google Scholar] [CrossRef] [Green Version]
- Meerts, P.; Misinzo, G.; Nauwynck, H.J. Enhancement of porcine circovirus type 2 replication in porcine cell lines by IFN-γ before and after treatment and by IFN-α after treatment. J. Interferon Cytokine Res. 2005, 25, 684–693. [Google Scholar] [CrossRef]
- Weesendorp, E.; Morgan, S.; Stockhofe-Zurwieden, N.; Popma-De Graaf, D.J.; Graham, S.P.; Rebel, J.M.J. Comparative analysis of immune responses following experimental infection of pigs with European porcine reproductive and respiratory syndrome virus strains of different virulence. Vet. Microbiol. 2013, 163, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.; Cho, W.-S.; Kim, B.; Chae, C. Expression of interferon-gamma and tumour necrosis factor-alpha in pigs experimentally infected with porcine reproductive and respiratory syndrome virus (PRRSV). J. Comp. Pathol. 2001, 127, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, S.; Huang, F.F.; Huang, Y.M.; Meng, X.J. Interferon-mediated enhancement of in vitro replication of porcine circovirus type 2 is influenced by an interferon stimulated response element in the PCV2 genome. Virus Res. 2009, 145, 236–243. [Google Scholar] [CrossRef]
- Ramamoorthy, S.; Opriessnig, T.; Pal, N.; Huang, F.F.; Meng, X.J. Effect of an interferon-stimulated response element (ISRE) mutant of porcine circovirus type 2 (PCV2) on PCV2-induced pathological lesions in a porcine reproductive and respiratory syndrome virus (PRRSV) co-infection model. Vet. Microbiol. 2011, 147, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Franzo, G.; Cortey, M.; Segalés, J.; Hughes, J.; Drigo, M. Phylodynamic analysis of porcine circovirus type 2 reveals global waves of emerging genotypes and the circulation of recombinant forms. Mol. Phylogenet. Evol. 2016, 100, 269–280. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Zhao, J.; Xing, G.; Li, G.; Wang, R.; Wang, Z.; Zhang, C.; Franzo, G.; Su, S.; Zhou, J. Genetic analysis and evolutionary changes of Porcine circovirus 2. Mol. Phylogenet. Evol. 2019, 139, 106520. [Google Scholar] [CrossRef]
- Rose, N.; Larour, G.; Le Diguerher, G.; Eveno, E.; Jolly, J.P.; Blanchard, P.; Oger, A.; Le Dimna, M.; Jestin, A.; Medec, F. Risk factors for porcine post-weaning multisystemic wasting syndrome (PMWS) in 149 French farrow-to-finish herds. Prev. Vet. Med. 2003, 61, 209–225. [Google Scholar] [CrossRef] [PubMed]
- Fraile, L.; Calsamiglia, M.; Mateu, E.; Espinal, A.; Cuxart, A.; Seminati, C.; Martin, M.; Domingo, M.; Segales, J. Prevalence of infection with porcine circovirus-2 (PCV2) and porcine reproductive and respiratory syndrome virus (PRRSV) in an integrated swine production system experiencing postweaning multisystemic wasting syndrome. Can. J. Vet. Res. 2009, 73, 308–312. [Google Scholar] [PubMed]
- Milek, D.; Wozniak, A.; Podgorska, K.; Stadejek, S. Do porcine parvoviruses 1 through 7 (PPV1-PPV7) have an impact on porcine circovirus type 2 (PCV2) viremia in pigs? Vet. Microbiol. 2020, 242, 108613. [Google Scholar] [CrossRef]
- Han, K.; Seo, H.W.; Oh, Y.; Kang, I.; Park, C.; Chae, C. Comparison of the virulence of European and North American genotypes of porcine reproductive and respiratory syndrome virus in experimentally infected pigs. Vet. J. 2013, 195, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Beaver, B.V.; Reed, W.; Leary, S.; McKiernan, B.; Bain, F.; Schultz, R.; Bennet, B.T.; Pascoe, P.; Shull, E.; Cork, L.C.; et al. 2000 Report of the AVMA panel on euthanasia. J. Am. Vet. Med. Assoc. 2001, 218, 669–696. [Google Scholar]
- Halbur, P.G.; Paul, P.S.; Frey, M.L.; Landgraf, J.; Eernisse, K.; Meng, X.-J.; Lum, M.A.; Andrews, J.J.; Rathje, J.A. Comparison of the pathogenicity of two US porcine reproductive and respiratory syndrome virus isolates with that of the Lelystad virus. Vet. Pathol. 1995, 32, 648–660. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Park, C.; Choi, K.; Chae, C. Comparison of three commercial one-dose porcine circovirus type 2 (PCV2) vaccines in a herd with concurrent circulation of PCV2b and mutant PCV2b. Vet. Microbiol. 2015, 177, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, C.A.; Del Castillo, J.R.E.; Music, N.; Fontaine, G.; Harel, J.; Tremblay, D. Development and use of a multiplex real-time quantitative polymerase chain reaction assay for detection and differentiation of Porcine circovirus-2 genotypes 2a and 2b in an epidemiological survey. J. Vet. Diagn. Investig. 2008, 20, 545–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasilk, A.; Callahan, J.D.; Christopher-Hennings, J.; Gay, T.A.; Fang, Y.; Dammen, M.; Reos, M.E.; Torremorell, M.; Polson, D.; Mellencamp, M.; et al. Detection of U.S., Lelystad, and European-like porcine reproductive and respiratory syndrome viruses and relative quantitation in boar semen and serum samples by real-time PCR. J. Clin. Microbiol. 2004, 42, 4453–4461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Chae, C. Expression of monocyte chemoattractant protein-1 and macrophage inflammatory protein-1 in porcine circovirus 2-induced granulomatous inflammation. J. Comp. Pathol. 2004, 131, 121–126. [Google Scholar] [CrossRef]
- Kim, D.; Kim, C.H.; Han, K.; Seo, H.W.; Oh, Y.; Park, C.; Kang, I.; Chae, C. Comparative efficacy of commercial Mycoplasma hyopneumoniae and porcine circovirus 2 (PCV2) vaccines in pigs experimentally infected with M. hyopneumoniae and PCV2. Vaccine 2011, 29, 3206–3212. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Weight (kg) | ADWG | ||
|---|---|---|---|
| Groups | 42 Days Old (0 dpi) | 63 Days Old (21 dpi) | Between 42 and 63 Days Old |
| PCV2a/PRRSV-2 | 8.47 ± 0.26 | 13.17 ± 0.71 a | 228.81 ± 34.45 a |
| PCV2b/PRRSV-2 | 8.55 ± 0.28 | 13.53 ± 0.87 a | 237.30 ± 41.45 a |
| PCV2d/PRRSV-2 | 8.32 ± 0.46 | 12.77 ± 0.68 a | 211.90 ± 27.56 a |
| PCV2a | 8.35 ± 0.10 | 16.03 ± 0.53 b | 365.87 ± 28.48 b |
| PCV2b | 8.42 ± 0.24 | 16.07 ± 0.73 b | 364.29 ± 33.37 b |
| PCV2d | 8.50 ± 0.23 | 16.22 ± 0.63 b | 367.46 ± 26.33 b |
| Negative control | 8.42 ± 0.26 | 16.17 ± 0.46 b | 369.05 ± 17.24 b |
| Groups | Microscopic Lung Lesion Scores | Microscopic Lymphoid Lesion Scores | PCV2-Antigen Positive Cells within Lymphoid Lesion |
|---|---|---|---|
| PCV2a/PRRSV-2 | 2.73 ± 0.52 a | 3.07 ± 0.39 a | 30.06 ± 4.57 a |
| PCV2b/PRRSV-2 | 2.80 ± 0.47 a | 3.13 ± 0.27 a | 30.94 ± 4.50 a |
| PCV2d/PRRSV-2 | 3.47 ± 0.43 b | 3.63 ± 0.23 b | 38.50 ± 4.37 b |
| PCV2a | 1.27 ± 0.24 c | 1.40 ± 0.18 c | 18.44 ± 3.27 c |
| PCV2b | 1.37 ± 0.20 c | 1.40 ± 0.22 c | 19.39 ± 4.68 c |
| PCV2d | 1.33 ± 0.30 c | 1.37 ± 0.23 c | 18.83 ± 4.49 c |
| Negative control | 0 ± 0 | 0 ± 0 | 0 ± 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suh, J.; Oh, T.; Park, K.; Yang, S.; Cho, H.; Chae, C. A Comparison of Virulence of Three Porcine Circovirus Type 2 (PCV2) Genotypes (a, b, and d) in Pigs Singularly Inoculated with PCV2 and Dually Inoculated with PCV2 and Porcine Reproductive and Respiratory Syndrome Virus. Pathogens 2021, 10, 891. https://doi.org/10.3390/pathogens10070891
Suh J, Oh T, Park K, Yang S, Cho H, Chae C. A Comparison of Virulence of Three Porcine Circovirus Type 2 (PCV2) Genotypes (a, b, and d) in Pigs Singularly Inoculated with PCV2 and Dually Inoculated with PCV2 and Porcine Reproductive and Respiratory Syndrome Virus. Pathogens. 2021; 10(7):891. https://doi.org/10.3390/pathogens10070891
Chicago/Turabian StyleSuh, Jeongmin, Taehwan Oh, Keehwan Park, Siyeon Yang, Hyejean Cho, and Chanhee Chae. 2021. "A Comparison of Virulence of Three Porcine Circovirus Type 2 (PCV2) Genotypes (a, b, and d) in Pigs Singularly Inoculated with PCV2 and Dually Inoculated with PCV2 and Porcine Reproductive and Respiratory Syndrome Virus" Pathogens 10, no. 7: 891. https://doi.org/10.3390/pathogens10070891