
Borrelia burgdorferi BmpA-BBK32 and BmpA-BBA64: New Recombinant Chimeric Proteins with Potential Diagnostic Value


Abstract
:1. Introduction
2. Results
2.1. Amino Acid Sequence Analysis
2.2. Expression and Purification of Chimeric Proteins BmpA-BBA64 and BmpA-BBK32
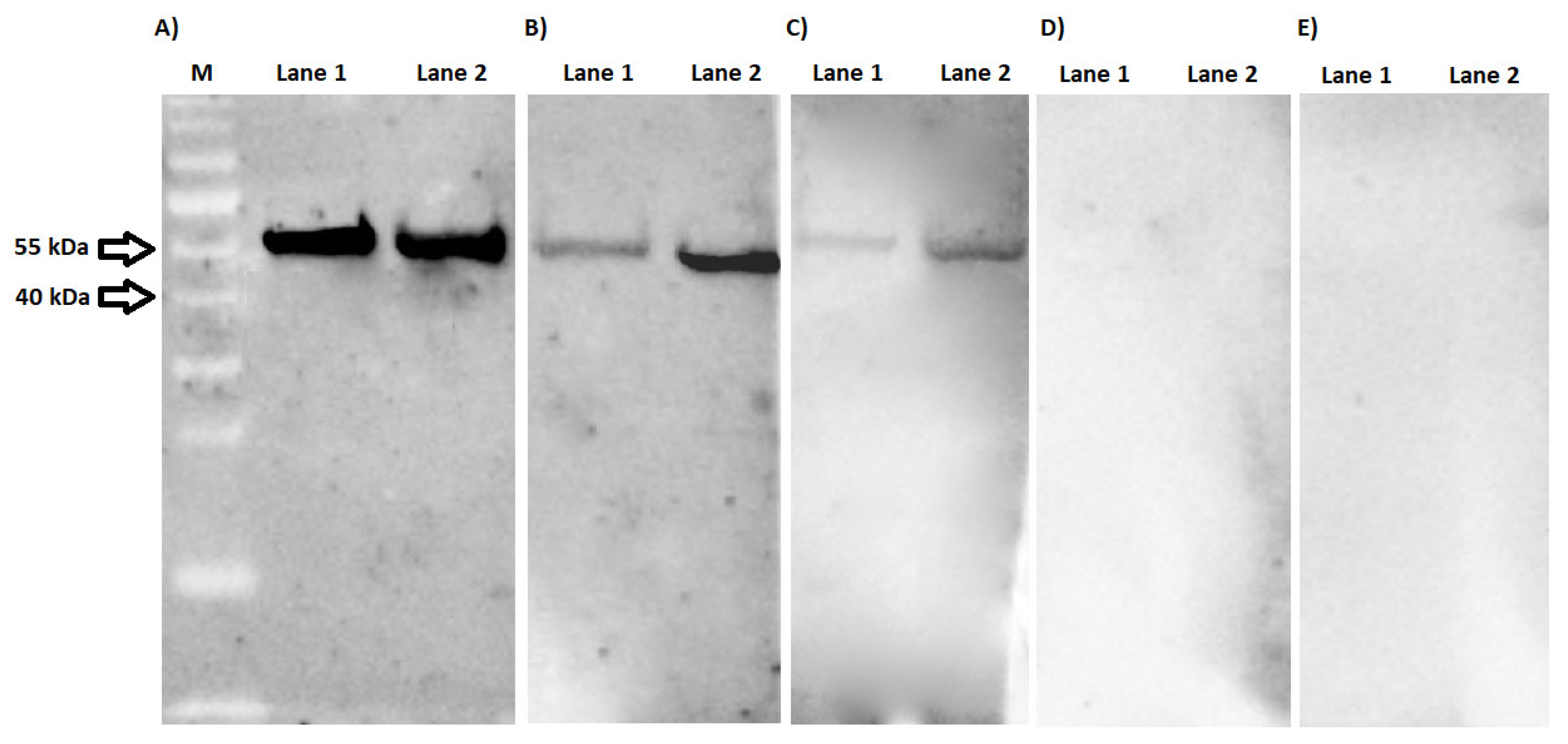
2.3. Western Blotting
3. Discussion
4. Materials and Methods
4.1. Serum Samples
4.2. Amino Acid Sequence Analysis
4.3. Construction of Recombinant Plasmids
4.4. Expression and Purification of Chimeric Proteins
4.5. Western Blot
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lohr, B.; Fingerle, V.; Norris, D.E.; Hunfeld, K.-P. Laboratory diagnosis of Lyme borreliosis: Current state of the art and future perspectives. Crit. Rev. Clin. Lab. Sci. 2018, 55, 219–245. [Google Scholar] [CrossRef] [PubMed]
- Stanek, G.; Reiter, M. The expanding Lyme Borrelia complex-clinical significance of genomic species? Clin. Microbiol. Infect. 2011, 17, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Pritt, B.S.; Respicio-Kingry, L.B.; Sloan, L.M.; Schriefer, M.E.; Replogle, A.J.; Bjork, J.; Liu, G.; Kingry, L.C.; Mead, P.S.; Neitzel, D.F.; et al. Borrelia mayonii sp. nov., a member of the Borrelia burgdorferi sensu lato complex, detected in patients and ticks in the upper midwestern United States. Int. J. Syst. Evol. Microbiol. 2016, 123, 4757–4763. [Google Scholar] [CrossRef] [PubMed]
- Golovchenko, M.; Vancová, M.; Clark, K.; Oliver, J.H.; Grubhoffer, L.; Rudenko, N. A divergent spirochete strain isolated from a resident of the southeastern United States was identified by multilocus sequence typing as Borrelia bissettii. Parasites Vectors 2016, 9, 4–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casjens, S.; Palmer, N.; Van Vugt, R.; Huang, W.M.; Stevenson, B.; Rosa, P.; Lathigra, R.; Sutton, G.; Peterson, J.; Dodson, R.J.; et al. A bacterial genome in flux: The twelve linear and nine circular extrachromosomal DNAs in an infectious isolate of the Lyme disease spirochete Borrelia burgdorferi. Mol. Microbiol. 2000, 35, 490–516. [Google Scholar] [CrossRef] [PubMed]
- Casjens, S.R.; Mongodin, E.F.; Qiu, W.G.; Luft, B.J.; Schutzer, S.E.; Gilcrease, E.B.; Huang, W.M.; Vujadinovic, M.; Aron, J.K.; Vargas, L.C.; et al. Genome stability of lyme disease spirochetes: Comparative genomics of Borrelia burgdorferi plasmids. PLoS ONE 2012, 7, e33280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glöckner, G.; Lehmann, R.; Romualdi, A.; Pradella, S.; Schulte-Spechtel, U.; Schilhabel, M.; Wilske, B.; Sühnel, J.; Platzer, M. Comparative analysis of the Borrelia garinii genome. Nucleic Acids Res. 2004, 32, 6038–6046. [Google Scholar] [CrossRef]
- Casjens, S.R.; Gilcrease, E.B.; Vujadinovic, M.; Mongodin, E.F.; Luft, B.J.; Schutzer, S.E.; Fraser, C.M.; Qiu, W.G. Plasmid diversity and phylogenetic consistency in the Lyme disease agent Borrelia burgdorferi. BMC Genom. 2017, 18, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Purser, J.E.; Norris, S.J. Correlation between plasmid content and infectivity in Borrelia burgdorferi. Proc. Natl. Acad. Sci. USA 2000, 97, 13865–13870. [Google Scholar] [CrossRef] [Green Version]
- Schwan, T.G.; Piesman, J.; Golde, W.T.; Dolan, M.C.; Rosa, P.A. Induction of an outer surface protein on Borrelia burgdorferi during tick feeding. Proc. Natl. Acad. Sci. USA 1995, 92, 2909–2913. [Google Scholar] [CrossRef] [Green Version]
- Ojaimi, C.; Brooks, C.; Casjens, S.; Rosa, P.; Elias, A.; Barbour, A.; Jasinskas, A.; Benach, J.; Katona, L.; Radolf, J.; et al. Profiling of temperature-induced changes in Borrelia burgdorferi gene expression by using whole genome arrays. Infect. Immun. 2003, 71, 1689–1705. [Google Scholar] [CrossRef] [Green Version]
- Kenedy, M.R.; Lenhart, T.R.; Akins, D.R. The role of Borrelia burgdorferi outer surface proteins. FEMS Immunol. Med. Microbiol. 2012, 66, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Schulte-Spechtel, U.; Fingerle, V.; Goettner, G.; Rogge, S.; Wilske, B. Molecular analysis of decorin-binding protein A (DbpA) reveals five major groups among European Borrelia burgdorferi sensu lato strains with impact for the development of serological assays and indicates lateral gene transfer of the dbpA gene. Int. J. Med. Microbiol. 2006, 296, 250–266. [Google Scholar] [CrossRef]
- Ivanova, L.; Christova, I.; Neves, V.; Aroso, M.; Meirelles, L.; Brisson, D.; Gomes-Solecki, M. Comprehensive seroprofiling of sixteen B. burgdorferi OspC: Implications for Lyme disease diagnostics design. Clin. Immunol. 2011, 35, 909–915. [Google Scholar] [CrossRef] [Green Version]
- Fraser, C.M.; Casjens, S.; Huang, W.M.; Sutton, G.G.; Clayton, R.; Lathigra, R.; White, O.; Ketchum, K.A.; Dodson, R.; Hickey, E.K.; et al. Genomic sequence of a Lyme disease spirochaete, Borrelia burgdorferi. Nature 1997, 390, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, R.D.; Howison, R.R.; Dietrich, G.; Patton, T.G.; Clifton, D.R.; Carroll, J.A. The bba64 gene of Borrelia burgdorferi, the Lyme disease agent, is critical for mammalian infection via tick bite transmission. Proc. Natl. Acad. Sci. USA 2010, 107, 7515–7520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilly, K.; Krum, J.G.; Bestor, A.; Jewett, M.W.; Grimm, D.; Bueschel, D.; Byram, R.; Dorward, D.; VanRaden, M.J.; Stewart, P.; et al. Borrelia burgdorferi OspC protein required exclusively in a crucial early stage of mammalian infection. Infect. Immun. 2006, 74, 3554–3564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ružić-Sabljić, E.; Cerar, T. Progress in the molecular diagnosis of Lyme disease. Expert Rev. Mol. Diagn. 2017, 17, 19–30. [Google Scholar] [CrossRef]
- Aguero-Rosenfeld, M.; Wang, G.; Scwartz, I.; Wormser, G. Diagnosis of Lyme Borreliosis. Clin. Microbiol. Rev. 2005, 18, 484–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguero-Rosenfeld, M.E.; Nowakowski, J.; McKenna, D.F.; Carbonaro, C.A.; Wormser, G.P. Serodiagnosis in early Lyme disease. J. Clin. Microbiol. 1993, 31, 3090–3095. [Google Scholar] [CrossRef] [Green Version]
- Engstrom, S.M.; Shoop, E.; Johnson, R.C. Immunoblot interpretation criteria for serodiagnosis of early Lyme disease. J. Clin. Microbiol. 1995, 33, 419–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, H.; Fingerle, V.; Hunfeld, K.P.; Huppertz, H.I.; Krause, A.; Rauer, S.; Ruf, B. Cutaneous Lyme borreliosis: Guideline of the German Dermatology Society. Ger. Med. Sci. 2017, 15, 1–31. [Google Scholar] [CrossRef]
- Theel, E.S. The Past, Present, and (Possible) Future of Serologic Testing for Lyme Disease. J. Clin. Microbiol. 2016, 54, 1191–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goettner, G.; Schulte-Spechtel, U.; Hillermann, R.; Liegl, G.; Wilske, B.; Fingerle, V. Improvement of Lyme Borreliosis serodiagnosis by a newly developed recombinant immunoglobulin G (IgG) and IgM line immunoblot assay and addition of VlsE and DbpA homologues. J. Clin. Microbiol. 2005, 43, 3602–3609. [Google Scholar] [CrossRef] [Green Version]
- Rawlins, M.L.; Gerstner, C.; Hill, H.R.; Litwin, C.M. Evaluation of a Western blot method for the detection of Yersinia antibodies: Evidence of serological cross-reactivity between Yersinia outer membrane proteins and Borrelia burgdorferi. Clin. Vaccine Immunol. 2005, 12, 1269–1274. [Google Scholar] [CrossRef] [Green Version]
- Magnarelli, L.A.; Fikrig, E.; Padula, S.J.; Anderson, J.F.; Flavell, R.A. Use of recombinant antigens of Borrelia burgdorferi in serologic tests for diagnosis of Lyme borreliosis. J. Clin. Microbiol. 1996, 34, 237–240. [Google Scholar] [CrossRef] [Green Version]
- Bruckbauer, H.R.; Preac-Mursic, V.; Fuchs, R.; Wilske, B. Cross-reactive proteins of Borrelia burgdorferi. Eur. J. Clin. Microbiol. Infect. Dis. 1992, 11, 224–232. [Google Scholar] [CrossRef]
- Roberts, W.C.; Mullikin, B.A.; Lathigra, R.; Hanson, M.S. Molecular analysis of sequence heterogeneity among genes encoding decorin binding proteins A and B of Borrelia burgdorferi sensu lato. Infect. Immun. 1998, 66, 5275–5285. [Google Scholar] [CrossRef] [Green Version]
- Theisen, M.; Frederiksen, B.; Lebech, A.M.; Vuust, J.; Hansen, K. Polymorphism in ospC gene of Borrelia burgdorferi and immunoreactivity of OspC protein: Implications for taxonomy and for use of OspC protein as a diagnostic antigen. J. Clin. Microbiol. 1993, 31, 2570–2576. [Google Scholar] [CrossRef] [Green Version]
- Gilmore, R.D.; Mbow, M.L.; Stevenson, B. Analysis of Borrelia burgdorferi gene expression during life cycle phases of the tick vector Ixodes scapularis. Microbes Infect. 2001, 3, 799–808. [Google Scholar] [CrossRef]
- Oksi, J.; Uksila, J.; Marjamaki, M.; Nikoskelainen, J.; Viljanen, M.K. Antibodies against whole sonicated Borrelia burgdorferi spirochetes, 41- kilodalton flagellin, and P39 protein in patients with PCR- or culture- proven late Lyme borreliosis. J. Clin. Microbiol. 1995, 33, 2260–2264. [Google Scholar] [CrossRef] [Green Version]
- Heikkilä, T.; Seppälä, I.; Saxén, H.; Panelius, J.; Peltomaa, M.; Julin, T.; Carlsson, S.A.; Lahdenne, P. Recombinant BBK32 protein in serodiagnosis of early and late Lyme borreliosis. J. Clin. Microbiol. 2002, 40, 1174–1180. [Google Scholar] [CrossRef] [Green Version]
- Mathiesen, M.J.; Christiansen, M.; Holm, A.; Åsbrink, E.; Theisen, M.; Hansen, K.; Åsbrink, E.V.A. Peptide-Based OspC Enzyme-Linked Immunosorbent Assay for serodiagnosis of Lyme Borreliosis. J. Clin. Microbiol. 1998, 36, 3474–3479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panelius, J.; Lahdenne, P.; Saxén, H.; Carlsson, S.A.; Heikkilä, T.; Peltomaa, M.; Lauhio, A.; Seppälä, I. Diagnosis of Lyme neuroborreliosis with antibodies to recombinant proteins DbpA, BBK32, and OspC, and VIsE IR6 peptide. J. Neurol. 2003, 250, 1318–1327. [Google Scholar] [CrossRef]
- Gilmore, R.D.; Kappel, K.J.; Johnson, B.J.B. Molecular characterization of a 35-kilodalton protein of Borrelia burgdorferi, an antigen of diagnostic importance in early Lyme disease. J. Clin. Microbiol. 1997, 35, 86–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holec, L.; Gąsior, A.; Brillowska-Dąbrowska, A.; Kur, J. Toxoplasma gondii: Enzyme-linked immunosorbent assay using different fragments of recombinant microneme protein 1 (MIC1) for detection of immunoglobulin G antibodies. Exp. Parasitol. 2008, 119, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Holec-Gąsior, L.; Kur, J. Toxoplasma gondii: Recombinant GRA5 antigen for detection of immunoglobulin G antibodies using enzyme-linked immunosorbent assay. Exp. Parasitol. 2010, 124, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Holec-Gąsior, L. Toxoplasma gondii recombinant antigens as tools for serodiagnosis of human toxoplasmosis: Current status of studies. Clin. Vaccine Immunol. 2013, 20, 1343–1351. [Google Scholar] [CrossRef] [Green Version]
- Rauer, S.; Spohn, N.; Rasiah, C.; Neubert, U.; Vogt, A. Enzyme-Linked Immunosorbent Assay using recombinant OspC and the internal 14-kDa flagellin fragment for serodiagnosis of early Lyme disease. J. Clin. Microbiol. 1998, 36, 857–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magalhães, F.B.; Castro Neto, A.L.; Nascimento, M.B.; Santos, W.J.T.; Medeiros, Z.M.; Neto, A.S.L.; Costa, D.L.; Costa, C.H.N.; Dos Santos, W.L.C.; Pontes de Carvalho, L.C.; et al. Evaluation of a new set of recombinant antigens for the serological diagnosis of human and canine visceral leishmaniasis. PLoS ONE 2017, 12, e0184867. [Google Scholar] [CrossRef] [Green Version]
- Ferra, B.T.; Holec-Gąsior, L.; Gatkowska, J.; Dziadek, B.; Dzitko, K.; Grąźlewska, W.; Lautenbach, D. The first study on the usefulness of recombinant tetravalent chimeric proteins containing fragments of SAG2, GRA1, ROP1 and AMA1 antigens in the detection of specific anti-Toxoplasma gondii antibodies in mouse and human sera. PLoS ONE 2019, 14, e0217866. [Google Scholar] [CrossRef]
- Holec-Gąsior, L.; Ferra, B.; Drapała, D. MIC1-MAG1-SAG1 chimeric protein, a most effective antigen for detection of human toxoplasmosis. Clin. Vaccine Immunol. 2012, 19, 1977–1979. [Google Scholar] [CrossRef] [Green Version]
- Holec-Gąsior, L.; Ferra, B.; Drapała, D.; Lautenbach, D.; Kur, J. A new MIC1-MAG1 recombinant chimeric antigen can be used instead of the Toxoplasma gondii lysate antigen in serodiagnosis of human toxoplasmosis. Clin. Vaccine Immunol. 2012, 19, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Drapała, D.; Holec-Gasior, L.; Kur, J. New recombinant chimeric antigens, P35-MAG1, MIC1-ROP1, and MAG1-ROP1, for the serodiagnosis of human toxoplasmosis. Diagn. Microbiol. Infect. Dis. 2015, 82, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Ferra, B.; Holec-Gąsior, L.; Kur, J. A new Toxoplasma gondii chimeric antigen containing fragments of SAG2, GRA1, and ROP1 proteins—impact of immunodominant sequences size on its diagnostic usefulness. Parasitol. Res. 2015, 114, 3291–3299. [Google Scholar] [CrossRef] [Green Version]
- Del-Rei, R.P.; Leony, L.M.; Celedon, P.A.F.; Zanchin, N.I.T.; dos Reis, M.G.; de Miranda Gomes, Y.; Schijman, A.G.; Longhi, S.A.; Santos, F.L.N. Detection of anti-Trypanosoma cruzi antibodies by chimeric antigens in chronic Chagas disease-individuals from endemic South American countries. PLoS ONE 2019, 14, e0215623. [Google Scholar] [CrossRef]
- Santos, W.J.T.; Tavares, D.H.C.; Castro Neto, A.L.; Nascimento, M.B.; Dhalia, R.; Albuquerque, A.L.; Costa, C.H.N.; Magalhães, F.B.; Rezende, A.M.; Neto, O.P.M. Gene design, optimization of protein expression and preliminary evaluation of a new chimeric protein for the serological diagnosis of both human and canine visceral leishmaniasis. PLoS Negl. Trop. Dis. 2020, 14, e0008488. [Google Scholar] [CrossRef] [PubMed]
- Norek, A.; Janda, L. Epitope mapping of Borrelia burgdorferi OspC protein in homodimeric fold. Protein Sci. 2017, 26, 796–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnaboldi, P.M.; Seedarnee, R.; Sambir, M.; Callister, S.M.; Imparato, J.A.; Dattwylera, R.J. Outer surface protein C peptide derived from Borrelia burgdorferi sensu stricto as a target for serodiagnosis of early Lyme disease. Clin. Vaccine Immunol. 2013, 20, 474–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotłowski, R.; Holec-Gąsior, L. Identification of evolutionary conserved DNA sequence and corresponding S21 ribosomal protein region for diagnostic purposes of all Borrelia spirochetes. Acta Biochim. Pol. 2019, 66, 119–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, F.T.; Philipp, M.T. Epitope mapping of the immunodominant invariable region of Borrelia burgdorferi VlsE in three host species. Infect. Immun. 2000, 68, 2349–2352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes-Solecki, M.J.C.; Dunn, J.J.; Luft, B.J.; Castillo, J.; Dykhuizen, D.E.; Yang, X.; Glass, D.; Dattwyler, R.J.; Glass, J.D. Recombinant Chimeric Borrelia proteins for diagnosis of Lyme disease. J. Clin. Microbiol. 2000, 38, 2530–2535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes-Solecki, M.; Wormser, G.; Persing, D.; Berger, B.; Glass, J.; Yang, X.; Dattwyler, R. A first-tier rapid assay for the serodiagnosis of Borrelia burgdorferi infection. Arch. Intern. Med. 2001, 161, 2015–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradshaw, G.L.; Thueson, R.K.; Uriona, T.J. Improved serodiagnostic performance for Lyme disease by use of two recombinant proteins in ELISA as compared to standardized two tier testing. J. Clin. Microbiol. 2017, 55, 3046–3056. [Google Scholar] [CrossRef] [Green Version]
- Ebady, R.; Niddam, A.F.; Boczula, A.E.; Kim, Y.R.; Gupta, N.; Tang, T.T.; Odisho, T.; Zhi, H.; Simmons, C.A.; Skare, J.T.; et al. Biomechanics of Borrelia burgdorferi Vascular Interactions. Cell Rep. 2016, 16, 2593–2604. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.R.; LeBlanc, K.T.; Leong, J.M. Fibronectin binding protein BBK32 of the Lyme disease spirochete promotes bacterial attachment to glycosaminoglycans. Infect. Immun. 2006, 74, 435–441. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.; Brissette, C.A.; Bowman, A.; Stevenson, B. Borrelia burgdorferi BmpA is a laminin-binding protein. Infect. Immun. 2009, 77, 4940–4946. [Google Scholar] [CrossRef] [Green Version]
- Roessler, D.; Hauser, U.; Wilske, B. Heterogeneity of BmpA (P39) among European isolates of Borrelia burgdorferi sensu lato and influence of interspecies variability on serodiagnosis. J. Clin. Microbiol. 1997, 35, 2752–2758. [Google Scholar] [CrossRef] [Green Version]
- Brandt, K.S.; Patton, T.G.; Allard, A.S.; Caimano, M.J.; Radolf, J.D.; Gilmore, R.D. Evaluation of the Borrelia burgdorferi BBA64 protein as a protective immunogen in mice. Clin. Vaccine Immunol. 2014, 21, 526–533. [Google Scholar] [CrossRef] [Green Version]
- Dąbrowski, S.; Kur, J. Cloning, overexpression, and purification of the recombinant His-tagged SSB protein of Escherichia coli and use in polymerase chain reaction amplification. Protein Expr. Purif. 1999, 16, 96–102. [Google Scholar] [CrossRef]


{kind=link}
| Protein | Genospecies | Accession no. |
|---|---|---|
| BmpA | B. burgdorferi B31 B. burgdorferi JD1 B. burgdorferi ZS7 B. burgdorferi N40 B. burgdorferi 156a | AAC66757.1 ADQ30753.1 ACK74429.1 ADQ29420.1 EEC22066.1 |
| B. afzelii Pko B. afzelii K78 B. afzelii HLJ01 B. afzelii Tom3107 | ABH01648.1 AJY72370.1 AFU74676.1 WP_044052195.1 | |
| B. garinii 20047 B. garinii NMJW1 B. garinii BgVir B. garinii PBr | AZA27810.1 AFT83711.1 AEW68720.1 WP_004791868.1 | |
| B. bavariensis PBi | AAU07235.1 | |
| B. spielmanii A14S | WP_006433486.1 | |
| BBK32 | B. burgdorferi B31 B. burgdorferi N40 B. burgdorferi IA B. burgdorferi 156a B. burgdorferi PKa2 B. burgdorferi ZS7 | AAC66134.1 WP_014540529.1 AAL84596.1 ACL33888.1 ACR57084.1 WP_012614929.1 |
| B. afzelii PKo B. afzelii K78 B. afzelii ACA-1 B. afzelii 570 B. afzelii A91 | ACR57085.1 AJY72931.1 ACJ73236.1 AAL84590.1 AAL84589.1 | |
| B. garinii 40 B. garinii PHei B. garinii TN B. garinii 50 B. garinii 46 | AAL84593.1 ACO05729.1 ACR57087.1 AAL84595.1 AAL84594.1 | |
| B. bavariensis PBi | ACR57086.1 | |
| B. spielmanii A14S | WP_012666206.1 | |
| BBA64 | B. burgdorferi B31 B. burgdorferi N40 B. burgdorferi JD1 B. burgdorferi 156a B. burgdorferi ZS7 | AAC66255.2 ACS94806.1 ACS94870.1 ACL33793.1 ACK74216.1 |
| B. afzelii PKo B. afzelii K78 B. afzelii ACA-1 | AEL70678.1 AJY72875.1 ACJ73562.1 | |
| B. garinii BgVir B. garinii Far04 B. garinii PBr | AEW69221.1 ACL35142.1 ACL34799.1 | |
| B. bavariensis PBi | AAT93822.1 | |
| B. spielmanii A14S | ACN53346.1 |
| Primer Name | Primer Sequence | Comments |
|---|---|---|
| bmpABglII (Forward) | 5’-GTGACAGATCTCGAATTTAAAATTGAGCTTC-3’ | BglII site and fragment of bmpA |
| bmpA-bba64 (Reverse) | 5’-GATAAAATTTGCCCAAGATTAATAAATTCTTTAAGAAAC-3’ | Fragments of bba64 and bmpA |
| bmpA-bbba64 (Forward) | 5’-GTTTCTTAAAGAATTTATTAATCTTGGGCAAATTTTATC-3’ | Fragments of bmpA and bba64 |
| bba64XhoI (Reverse) | 5’-CATAACTCGAGCTGAATTGGAGCAAG-3’ | XhoI site and fragment of bba64 |
| bmpA-bbk32 (Reverse) | 5’-GATATCGATTGCTTAATCTAATAAATTCTTTAAGAAACTTC-3’ | Fragments of bbk32 and bmpA |
| bmpA-bbk32 (Forward) | 5’-GAAGTTTCTTAAAGAATTTATTAGATTAAGCAATCGATATC-3’ | Fragments of bmpA and bbk32 |
| bbk32XhoI (Reverse) | 5’-CATAACTCGAGGTACCAAACGCCATTC-3’ | XhoI site and fragment of bbk32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grąźlewska, W.; Ferra, B.; Rudzińska, M.; Holec-Gąsior, L. Borrelia burgdorferi BmpA-BBK32 and BmpA-BBA64: New Recombinant Chimeric Proteins with Potential Diagnostic Value. Pathogens 2021, 10, 767. https://doi.org/10.3390/pathogens10060767
Grąźlewska W, Ferra B, Rudzińska M, Holec-Gąsior L. Borrelia burgdorferi BmpA-BBK32 and BmpA-BBA64: New Recombinant Chimeric Proteins with Potential Diagnostic Value. Pathogens. 2021; 10(6):767. https://doi.org/10.3390/pathogens10060767
Chicago/Turabian StyleGrąźlewska, Weronika, Bartłomiej Ferra, Monika Rudzińska, and Lucyna Holec-Gąsior. 2021. "Borrelia burgdorferi BmpA-BBK32 and BmpA-BBA64: New Recombinant Chimeric Proteins with Potential Diagnostic Value" Pathogens 10, no. 6: 767. https://doi.org/10.3390/pathogens10060767