
Molecular Detection of Novel Borrelia Species, Candidatus Borrelia javanense, in Amblyomma javanense Ticks from Pangolins

Abstract
:1. Introduction
2. Results
2.1. Identification of Borrelia sp. in Ticks
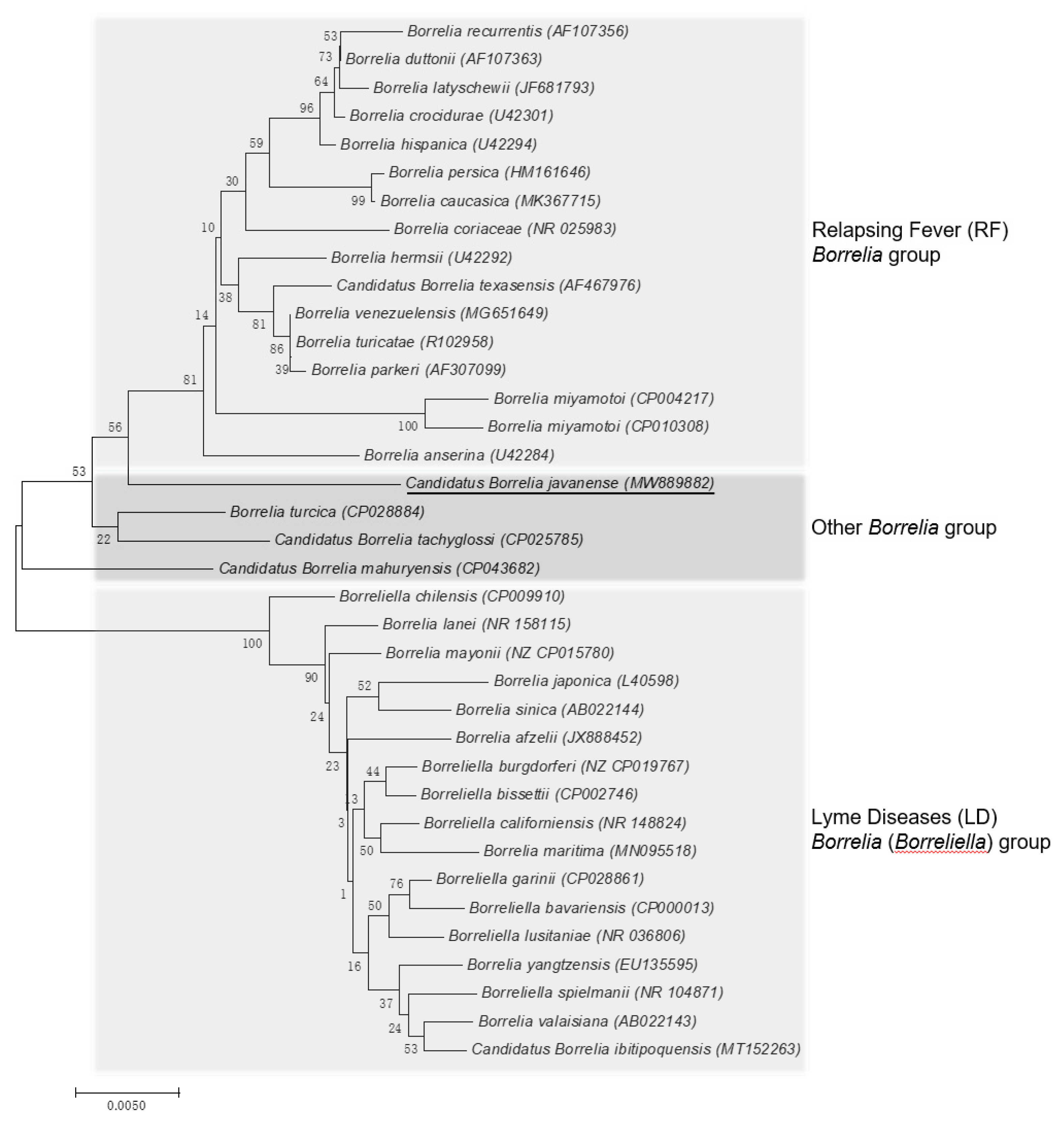
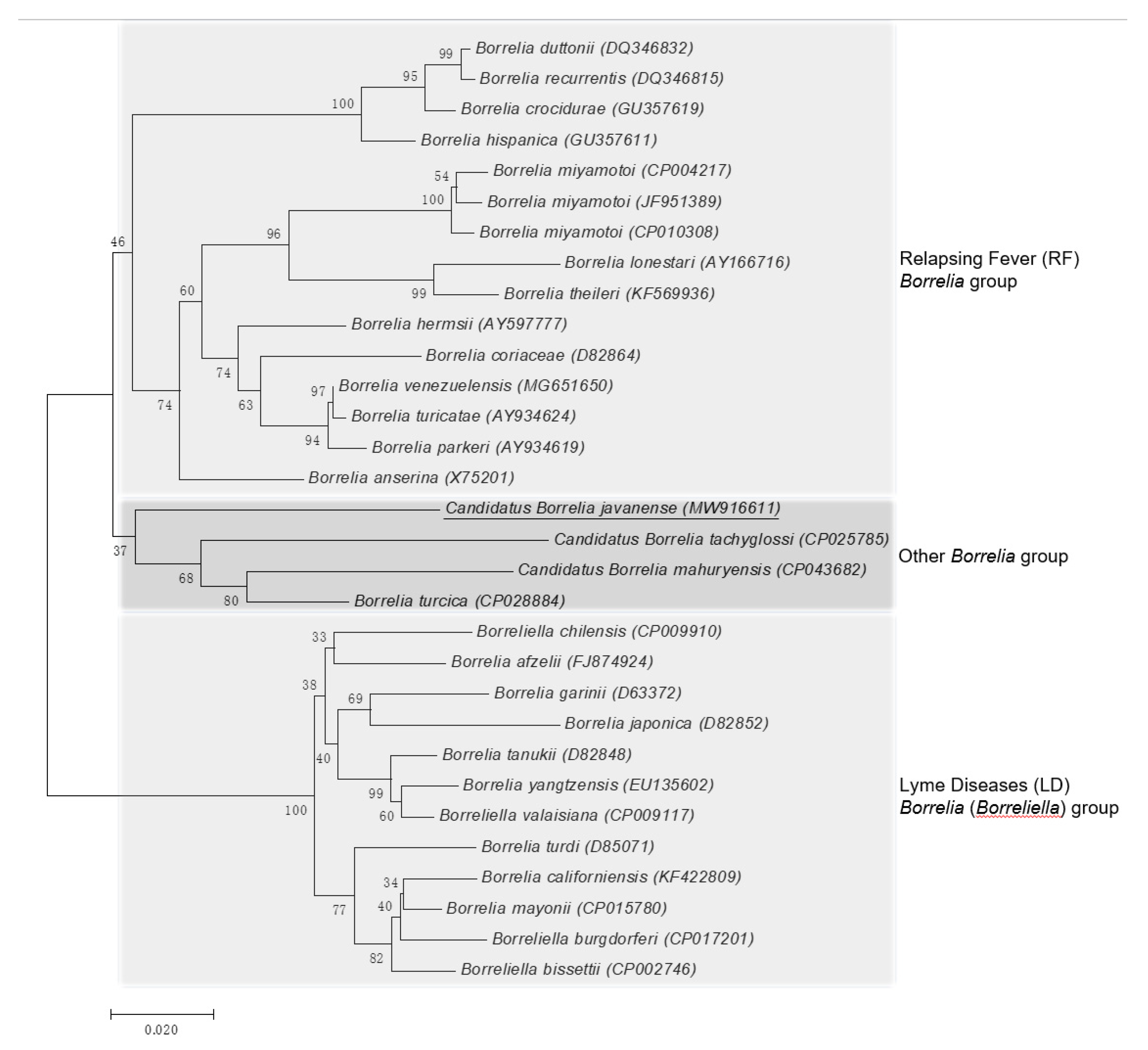
2.2. Genetic Characteristics of Borrelia sp.
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Tick Sample Collection and Identification
4.3. DNA Extraction, Borrelia-Specific PCR, and Sequencing
4.4. Sequence Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Talagrand-Reboul, E.; Boyer, P.H.; Bergstrom, S.; Vial, L.; Boulanger, N. Relapsing Fevers: Neglected Tick-Borne Diseases. Front. Cell Infect. Microbiol. 2018, 8, 98. [Google Scholar] [CrossRef] [Green Version]
- Margos, G.; Marosevic, D.; Cutler, S.; Derdakova, M.; Diuk-Wasser, M.; Emler, S.; Fish, D.; Gray, J.; Hunfeldt, K.-P.; Jaulhac, B.; et al. There is inadequate evidence to support the division of the genus Borrelia. Int. J. Syst. Evol. Microbiol. 2017, 67, 1081–1084. [Google Scholar] [CrossRef]
- Ras, N.M.; Lascola, B.; Postic, D.; Cutler, S.J.; Rodhain, F.; Baranton, G.; Raoult, D. Phylogenesis of relapsing fever Borrelia spp. Int. J. Syst. Bacteriol. 1996, 46, 859–865. [Google Scholar] [CrossRef]
- Takano, A.; Nakao, M.; Masuzawa, T.; Takada, N.; Yano, Y.; Ishiguro, F.; Fujita, H.; Ito, T.; Ma, X.; Oikawa, Y.; et al. Multilocus sequence typing implicates rodents as the main reservoir host of human-pathogenic Borrelia garinii in Japan. J. Clin. Microbiol. 2011, 49, 2035–2039. [Google Scholar] [CrossRef] [Green Version]
- Adeolu, M.; Gupta, R.S. A phylogenomic and molecular marker based proposal for the division of the genus Borrelia into two genera: The emended genus Borrelia containing only the members of the relapsing fever Borrelia, and the genus Borreliella gen. nov. containing the members of the Lyme disease Borrelia (Borrelia burgdorferi sensu lato complex). Antonie Leeuwenhoek 2014, 105, 1049–1072. [Google Scholar] [CrossRef]
- Guner, E.S.; Watanabe, M.; Hashimoto, N.; Kadosaka, T.; Kawamura, Y.; Ezaki, T.; Kawabata, H.; Imai, Y.; Kaneda, K.; Masuzawa, T. Borrelia turcica sp. nov., isolated from the hard tick Hyalomma aegyptium in Turkey. Int. J. Syst. Evol. Microbiol. 2004, 54, 1649–1652. [Google Scholar] [CrossRef] [PubMed]
- Loh, S.-M.; Gofton, A.W.; Lo, N.; Gillett, A.; Ryan, U.M.; Irwin, P.J.; Oskam, C.L. Novel Borrelia species detected in echidna ticks, Bothriocroton concolor, in Australia. Parasit Vectors 2016, 9, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loh, S.M.; Gillett, A.; Ryan, U.; Irwin, P.; Oskam, C. Molecular characterization of ‘Candidatus Borrelia tachyglossi’ (family Spirochaetaceae) in echidna ticks, Bothriocroton concolor. Int. J. Syst. Evol. Microbiol. 2017, 67, 1075–1080. [Google Scholar] [CrossRef] [Green Version]
- Panetta, J.L.; Sima, R.; Calvani, N.E.D.; Hajdusek, O.; Chandra, S.; Panuccio, J.; Slapeta, J. Reptile-associated Borrelia species in the goanna tick (Bothriocroton undatum) from Sydney, Australia. Parasit Vectors 2017, 10, 616. [Google Scholar] [CrossRef] [Green Version]
- Takano, A.; Goka, K.; Une, Y.; Shimada, Y.; Fujita, H.; Shiino, T.; Watanabe, H.; Kawabata, H. Isolation and characterization of a novel Borrelia group of tick-borne borreliae from imported reptiles and their associated ticks. Environ. Microbiol. 2010, 12, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, E.A.; Williamson, P.C.; Billingsley, P.M.; Seals, J.P.; Ferguson, E.E.; Allen, M.S. Frequency and Distribution of Rickettsiae, Borreliae, and Ehrlichiae Detected in Human-Parasitizing Ticks, Texas, USA. Emerg. Infect. Dis. 2016, 22, 312–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacheco, A.; Cordeiro, M.D.; Cepeda, M.B.; Luz, H.R.; Cardozo, S.V.; Berto, B.P.; Guterres, A.; Fonseca, A.H. Hemoparasites in ticks of wild birds of Serra dos Orgaos National Park, state of Rio de Janeiro, Brazil. Rev. Bras. Parasitol. Vet. 2019, 28, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Cicuttin, G.L.; De Salvo, M.N.; Venzal, J.M.; Nava, S. Borrelia spp. in ticks and birds from a protected urban area in Buenos Aires city, Argentina. Ticks Tick Borne Dis. 2019, 10, 101282. [Google Scholar] [CrossRef]
- Lee, J.K.; Smith, W.C.; McIntosh, C.; Ferrari, F.G.; Moore-Henderson, B.; Varela-Stokes, A. Detection of a Borrelia species in questing Gulf Coast ticks, Amblyomma maculatum. Ticks Tick Borne Dis. 2014, 5, 449–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gofton, A.W.; Margos, G.; Fingerle, V.; Hepner, S.; Loh, S.M.; Ryan, U.; Irwin, P.; Oskam, C.L. Genome-wide analysis of Borrelia turcica and ‘Candidatus Borrelia tachyglossi’ shows relapsing fever-like genomes with unique genomic links to Lyme disease Borrelia. Infect. Genet. Evol. 2018, 66, 72–81. [Google Scholar] [CrossRef]
- Kaenkan, W.; Nooma, W.; Chelong, I.A.; Baimai, V.; Trinachartvanit, W.; Ahantarig, A. Reptile-associated Borrelia spp. In Amblyomma ticks, Thailand. Ticks Tick Borne Dis. 2020, 11, 101315. [Google Scholar] [CrossRef] [PubMed]
- Trinachartvanit, W.; Hirunkanokpun, S.; Sudsangiem, R.; Lijuan, W.; Boonkusol, D.; Baimai, V.; Ahantarig, A. Borrelia sp. phylogenetically different from Lyme disease- and relapsing fever-related Borrelia spp. in Amblyomma varanense from Python reticulatus. Parasit Vectors 2016, 9, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kingry, L.C.; Anacker, M.; Pritt, B.; Bjork, J.; Respicio-Kingry, L.; Liu, G.; Sheldon, S.; Boxrud, D.; Strain, A.; Oatman, S.; et al. Surveillance for and Discovery of Borrelia Species in US Patients Suspected of Tickborne Illness. Clin. Infect. Dis. 2018, 66, 1864–1871. [Google Scholar] [CrossRef]
- Binetruy, F.; Garnier, S.; Boulanger, N.; Talagrand-Reboul, E.; Loire, E.; Faivre, B.; Noel, V.; Buysse, M.; Duron, O. A novel Borrelia. species, intermediate between Lyme disease and relapsing fever groups, in neotropical passerine-associated ticks. Sci Rep. 2020, 10, 10596. [Google Scholar] [CrossRef] [PubMed]
- Mihalca, A.D.; Gherman, C.M.; Cozma, V. Coendangered hard-ticks: Threatened or threatening? Parasit Vectors 2011, 4, 71. [Google Scholar] [CrossRef] [Green Version]
- Kollars, T.M., Jr.; Sithiprasasna, R. New host, and distribution record of Amblyomma javanense (Acari: Ixodidae) in Thailand. J. Med. Entomol. 2000, 37, 640. [Google Scholar] [CrossRef]
- Guglielmone, A.A.; Nava, S. Names for Ixodidae (Acari: Ixodoidea): Valid, synonyms, incertae sedis, nomina dubia, nomina nuda, lapsus, incorrect and suppressed names--with notes on confusions and misidentifications. Zootaxa 2014, 3767, 1–256. [Google Scholar] [CrossRef]
- Hassan, M.; Sulaiman, M.H.; Lian, C.J. The prevalence and intensity of Amblyomma javanense infestation on Malayan pangolins (Manis javanica Desmarest) from Peninsular Malaysia. Acta Trop. 2013, 126, 142–145. [Google Scholar] [CrossRef] [PubMed]
- Voltzit, O.V.; Keirans, J.E. A review of Asian Amblyomma species (Acari, Ixodida, Ixodidae). Acarina 2002, 10, 95–136. [Google Scholar]
- Kwak, M.L.; Hsu, C.D.; Douay, G.; Ahmad, A.A. The first authenticated record of the pangolin tick Amblyomma javanense (Acari: Ixodidae) in Singapore, with notes on its biology and conservation. Exp. Appl. Acarol. 2018, 76, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Platonov, A.E.; Karan, L.S.; Kolyasnikova, N.M.; Makhneva, N.A.; Toporkova, M.G.; Maleev, V.V.; Fish, D.; Krause, P.J. Humans infected with relapsing fever spirochete Borrelia miyamotoi, Russia. Emerg. Infect. Dis. 2011, 17, 1816–1823. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′-3′) | Annealing Temperature | Expected Product Size | Reference |
|---|---|---|---|---|---|
| 16S rRNA | External | ||||
| Bor-0F | AAAATAACGAAGAGTTTGATCCTGG | This study | |||
| Bor-1533R | GTGATCCAGCCACACTTTCCAGTA | 55 °C | 1533 bp | This study | |
| Internal | |||||
| Brm1 | CGCTGTAAACGATGCACACTTGGTGTTAATC | [26] | |||
| Brm2 | CGGCAGTCTCGTCTGAGTCCCCATCT | 60 °C | 353 bp | [26] | |
| Brm1-R | GATTAACACCAAGTGTGCATCGTTTACAGCG | 55 °C | 790 bp/Bor-0F | This study | |
| flaB | External | ||||
| FLA120F | AGAATTAATMGHGCWTCTGATGATG | [26] | |||
| Flab764R | GCATCTTCGATCTTTGAAAGTGACATATT | 55 °C | 645 bp | This study | |
| Internal | |||||
| Flab625R | CTGGAGCTGCTTGAGCACCTTCT | 55 °C | 506 bp/FLA120F | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, B.-G.; Wu, A.-Q.; Jiang, J.-F.; Yuan, T.-T.; Xu, Q.; Lv, C.-L.; Chen, J.-J.; Sun, Y.; Fang, L.-Q.; Ruan, X.-D.; et al. Molecular Detection of Novel Borrelia Species, Candidatus Borrelia javanense, in Amblyomma javanense Ticks from Pangolins. Pathogens 2021, 10, 728. https://doi.org/10.3390/pathogens10060728
Jiang B-G, Wu A-Q, Jiang J-F, Yuan T-T, Xu Q, Lv C-L, Chen J-J, Sun Y, Fang L-Q, Ruan X-D, et al. Molecular Detection of Novel Borrelia Species, Candidatus Borrelia javanense, in Amblyomma javanense Ticks from Pangolins. Pathogens. 2021; 10(6):728. https://doi.org/10.3390/pathogens10060728
Chicago/Turabian StyleJiang, Bao-Gui, Ai-Qiong Wu, Jia-Fu Jiang, Ting-Ting Yuan, Qiang Xu, Chen-Long Lv, Jin-Jin Chen, Yi Sun, Li-Qun Fang, Xiang-Dong Ruan, and et al. 2021. "Molecular Detection of Novel Borrelia Species, Candidatus Borrelia javanense, in Amblyomma javanense Ticks from Pangolins" Pathogens 10, no. 6: 728. https://doi.org/10.3390/pathogens10060728