1. Introduction
Bartonella spp. are Gram-negative, facultative, fastidious intracellular bacteria causing various pathological changes and bacteremia in their natural and accidental hosts. Several bloodsucking arthropods have been confirmed as vectors transmitting these pathogens including cat fleas (
Ctenocephalides felis) [
1], human fleas (
Pulex irritans) [
2], human lice (
Pediculus humanus) [
3], and sandflies (
Luztomyia verrucarum) [
4]. Additionally, some tick species are suggested as possible vectors of
Bartonella spp. such as the brown dog tick (
Rhipicephalus sanguineus) [
5] and the Ixodid tick (
Ixodes spp. [
6] and
Dermacentor spp. [
7]). Up to the present, a number of new species of
Bartonella and candidatus Bartonella have been described and more than 35 species have revealed their whole genome.
Human bartonelloses are caused by
B. henselae (cat-scratch disease),
B. bacilliformis (Carrion’s disease and verruga peruana), and
B. quintana (trench fever) [
8]. Humans play a role as accidental hosts being infected by
Bartonella spp. from various animal hosts, both companion and wild animals [
9]. Bartonellosis mainly affects immunocompromised patients, however, only regional lymphadenopathy occurs in immunocompetent people [
10]. At least 13 species are considered as zoonotic agents and the most zoonotic-prevalent species is
B. henselae, which can cause an asymptomatic infection among the hosts, such as carnivores, rodents, and ruminants [
10,
11,
12].
Dogs and cats are defined as family members sharing household environments with humans in both urban and rural areas, and they are considered as a source of zoonotic infection, especially of bartonellosis [
13]. In addition, dogs and cats are major mammals that can act as reservoirs of
Bartonella spp. [
14] and are defined as the main animals transmitting these pathogens to humans. Importantly, cats have been described as a primary reservoir animal of human bartonellosis [
15], but dogs are defined as accidental hosts of these pathogens due to a lack of clear evidence [
16]. The clinical signs in cats are mostly mild compared with dogs, however, some hematological parameters of both animals are changed [
14]. The evaluation of blood parameters in
Bartonella-infected animals, particularly cats and dogs, is still unclear. Several studies in other countries have evaluated the blood parameters in
Bartonella-infected animals. Importantly, blood parameters are frequently used to diagnose and monitor animal health. Even if domestic cats and dogs are popular in Thailand, there has been little hematological study comparing uninfected- and infected-
Bartonella animals.
In Thailand, dogs and cats that are both free-ranging and owned cohabit human communities. Bangkok is a province where the density of temples and household are considered high. Free-ranging animals were frequently found in temples which were a place for cultural activities and festivals, and owned animals were also found in the household. Significantly, the pattern of familiarity between humans and their belonging animals might induce zoonotic infection, particularly bartonellosis. The main proven modes of transmission in the human are animal scratch or bite with
Bartonella-contaminated vector feces, and blood-sucking arthropod bite [
14]. However, there have been few studies reporting evidence of
Bartonella in domestic animals in Thailand. The objectives of this study were to investigate the prevalence of
Bartonella spp. in domestic dogs and cats and to evaluate the hematological parameters of
Bartonella-infected dogs and cats in Bangkok, Thailand.
3. Discussion
This study detected evidence of
Bartonella spp. in domestic dogs and cats in Bangkok, Thailand. The results indicated that the prevalence of
Bartonella in the total cat population evaluated was 2.53%, however,
Bartonella spp. was observed only in free-ranging cats. The current study found two species of
Bartonella (
B. henselae and
B. clarridgeiae) which was similar with another study in Thailand that also reported that
B. henselae and
B. clarridgeiae as the main species in this region [
17]. Evidence of
Bartonella bacteremia was found in 13 cats, however, all dogs were negative to
Bartonella in this study. In cats, this positive proportion contrasted with other
Bartonella prevalence reported in Brazil [
18,
19], Greece [
20], Italy [
21,
22], Germany [
23], New Caledonia [
24], USA [
25,
26,
27,
28,
29], Saudi Arabia [
30], Israel [
31], Portugal [
32], France [
33,
34], and Thailand [
17], but was significantly similar with studies in Italy [
35,
36], Spain [
37], Scotland [
38], Ireland [
39], and China [
13]. Moreover, our study also demonstrated that the prevalence of
Bartonella spp. in dogs was not significantly different from studies in China [
13], Brazil [
18], Spain [
37], Cape Verde [
40], and Grenada [
41], but contrasted with other studies where
Bartonella spp. were detected in cardiac tissues [
42,
43] and blood [
42,
44,
45,
46]. Importantly, there was a chance of an event that
Bartonella was positive only in cardiac tissue but no evidence in blood [
42]. Conversely, the detection in dogs in the current study was different from another study conducted in Thailand [
46]. Additionally, dogs were defined as accidental hosts with some cardiac abnormalities [
47] and cats were the major reservoir hosts presenting only subclinical disease or non-specific signs [
48]. This explained why dogs were seldom detected as having
Bartonella DNA in their blood samples. Comparisons of prevalence differences between recent and previous studies are presented in
Supplementary Table S3. This different prevalence was affected from several different factors mainly including the detection technique, studied region, and host type. The geographic location and host type were considered as factors related to the prevalence of
Bartonella [
49]. Strayed animals frequently revealed a higher prevalence of
Bartonella than owned animals [
49] and these were caused by the difference of caring pattern, ectoparasite control, and ranging area. Thus, the negative prevalence in owned animals in this study was also caused by these aspects. Importantly, cyclic bacteremia of
Bartonella was the main factor causing a false negative. For the detection method, sensitivity was the impacted factor that caused the different prevalence. Moreover, the concentration of
Bartonella DNA might not match with the sensitivity of the test [
47] and caused the different prevalence rate. The real-time and nested PCR presented a higher sensitivity than conventional PCR for
Bartonella detection [
13]. However, cPCR was less time and cost consumption than others. Additionally, various primers for cPCR had been completely produced compared to the real-time and nested PCR.
The prevalence of
Bartonella spp. in the current study is quite low. The low prevalence in this study was suspected to be caused by direct pathogen detection from the bacteremia level in blood samples [
44,
50,
51] and cyclic bacteremia characteristic of
Bartonella spp. [
52] at the sampling time. Additionally, some studies mentioned the lack of PCR sensitivity for
Bartonella spp. detection in clinical samples, e.g., blood, other body fluids, and tissues [
53]. Compared with the bacterial culture, PCR was suggested as a highly successful technique for
Bartonella diagnosis in humans and experimental cats [
54]. Although nested PCR was more sensitive than cPCR, the available primers for cPCR that used to target
Bartonella spp. were quite more abundant. Due to the phylogenetic study, the longest sequence product of cPCR was more useful than the nested PCR for analysis. Additionally, the
Bartonella Alphaproteobacteria growth medium (BAPGM) pre-enrichment was suggested for the specimen culture in
Bartonella spp. to increase the sensitivity rate of PCR detection compared with the direct DNA extraction from pure samples [
46]. Even if BAPGM increased the PCR sensitivity, non-
Bartonella spp. had been also co-isolated with
Bartonella spp. especially environmental bacteria, non-pathogenic commensal organism, and skin normal flora [
55,
56], and contamination was considered.
Only free-ranging cats were found with
Bartonella DNA. Additionally, some studies concluded that the prevalence of
Bartonella infection in stray animals was higher than in pet animals [
16,
49]. However, the current study indicated no associated factor that was significantly related with
Bartonella-positive animals. This was consistent with other studies that found no difference in prevalence based on gender, residing location, living pattern, age, and ectoparasitic appearance [
21]. A contrasting result in Thailand reported that young cats and flea-infested cats were mostly found with
Bartonella spp [
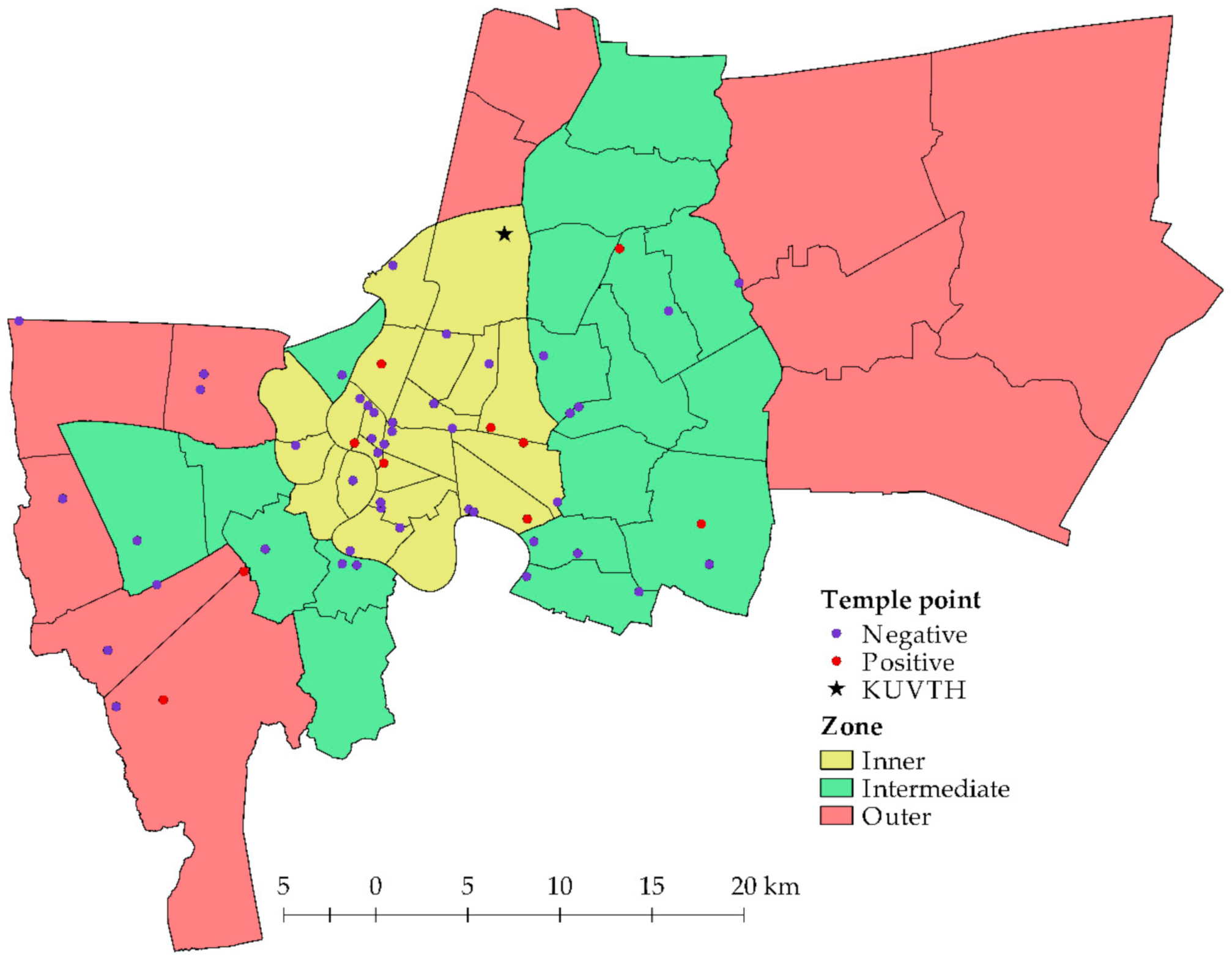
17]. We found a high positive proportion in the inner zone of Bangkok where there are many residences on crown land. This area should be considered for the introduction of disease transmission protection against zoonotic diseases, especially bartonellosis.
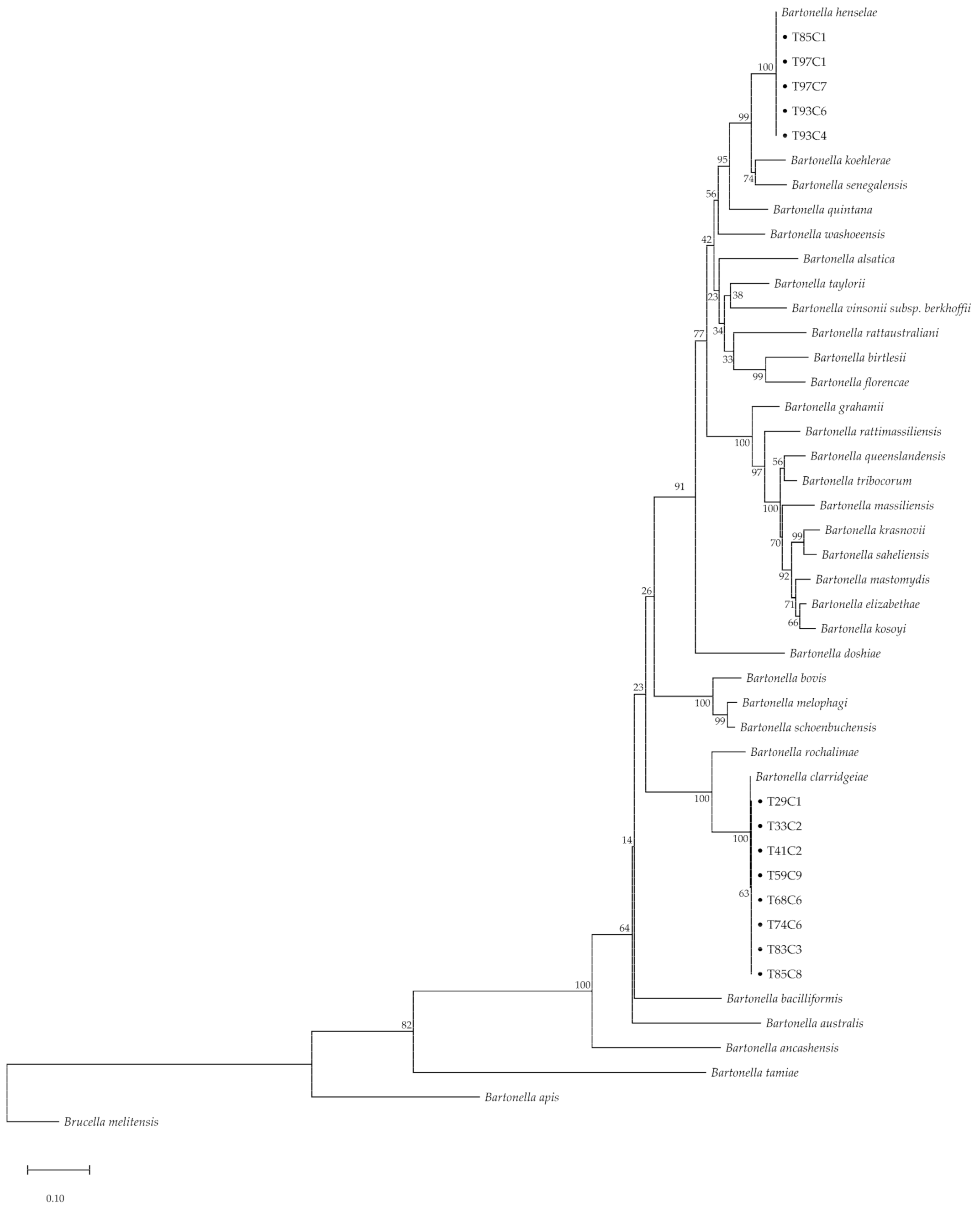
Bartonella henselae and
B. clarridgeiae were the major species detected in cats [
16]. These species have been reported in various countries in Southeast Asia [
57,
58,
59,
60]. In previous reports, Thailand had endocarditis patients infected with
B. henselae [
61] and
B. vinsonii subsp.
arupensis [
62]. Of these, there was evidence to support the impact of zoonotic
Bartonella infection in Thailand.
Bartonella henselae was reported as the main species causing zoonotic infection, especially via cat-scratch disease. In Southeast Asia, it has been suggested that
B. henselae and
B. clarridgeiae are the species affecting human health and of veterinary importance [
63]. In the main,
B. henselae and
B. clarridgeiae are transmitted to humans via cat fleas (
C. felis) [
64] which are generally found on cats and dogs worldwide [
65] and
C. felis is the major vector as the pathogen-transmissible insect for feline bartonellosis [
66].
The current study used primers targeting the
gltA gene of
Bartonella spp. for screening. Using
gltA detection, this gene had a high specificity for
Bartonella detection in extracted DNA samples from blood [
67]. The current BLAST results for the
gltA and r
ibC-amplified amplicons revealed a similar species of
Bartonella. Additionally, other studies mentioned that positive
Bartonella samples targeting the
ribC gene were also positive to
gltA in all samples due to the higher detection power of
gltA [
68,
69,
70]. However, there was no correlation for the BLAST results with the other housekeeping genes (
rpoB,
ftsZ, and
groEL) in the current study. This might have been due to the low discriminatory power of some genes, except for
rpoB, for species differentiation in the
Bartonella genus [
67]. On the other hand, the lack of correlated BLAST results might have been due to the co-infection of
Bartonella spp. in individual cats. From the results, it was difficult to make any conclusions regarding the mixed infection at this time. Nevertheless, co-infection should be investigated using other more sensitive techniques.
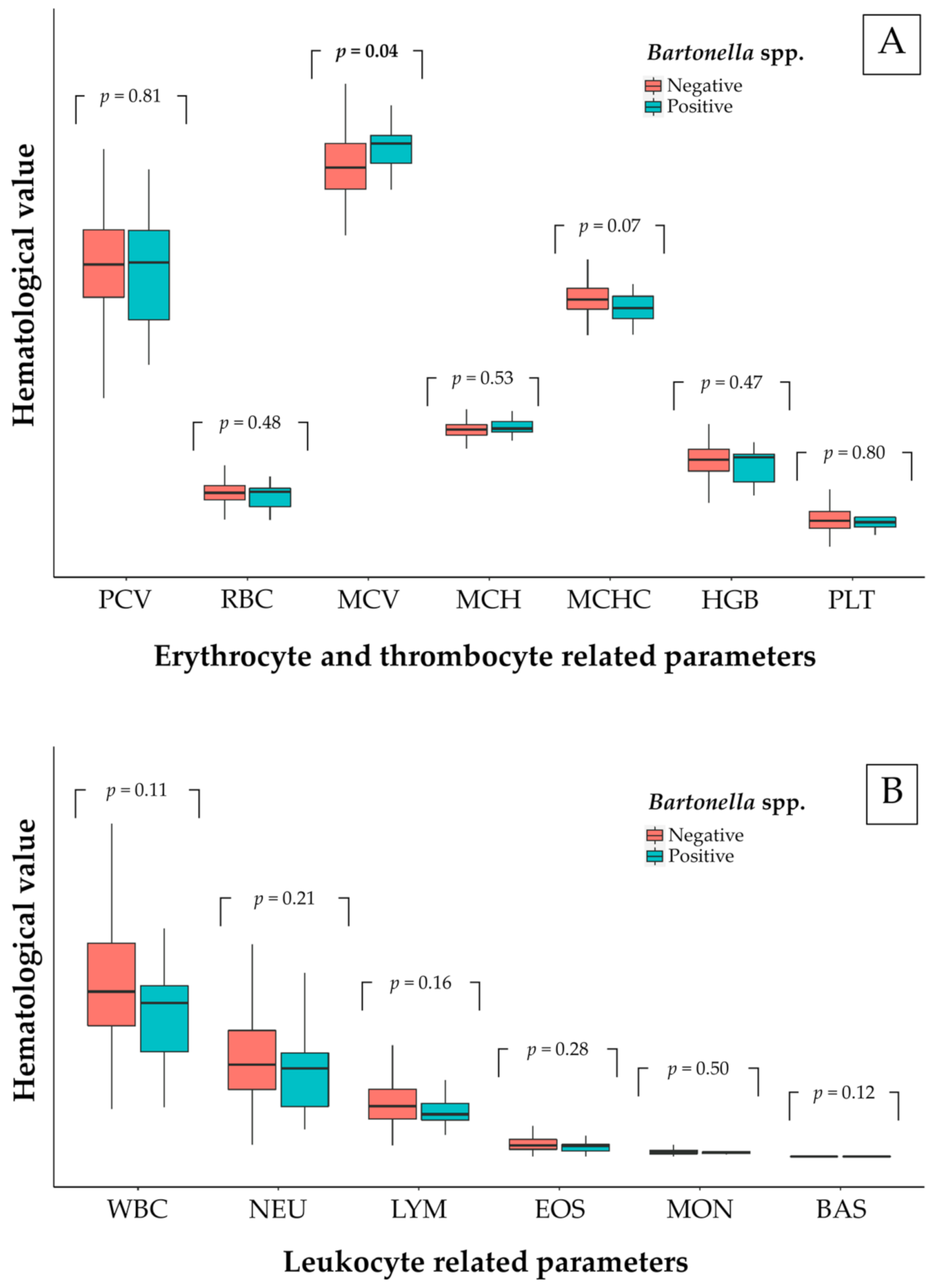
Bartonella-positive free-ranging cats had raised values for MCV. Over 36% of bacteremic cats had low values for HGB and MCHC. Another study in dogs mentioned that
Bartonella-infected blood was significantly low in hemoglobin (HGB), erythrocytic mass (RBC), and hematocrit (HCT) [
71]. The same trend was reported in a hematological study in camels, where the
Bartonella-positive animals were significantly low in HGB, MCH, and MCHC [
72]. The mechanisms of anemia were studied mainly in
B. bacilliformis and were described in various ways [
73].
Bartonella-positive animals frequently showed anemia, eosinophilia, neutrophilia, and thrombocytopenia in [
14,
74]. Additionally,
Bartonella-infected cats mostly showed mild or no abnormality of blood panels [
18]. However, one hematological study in domestic cats revealed that MCV was noticeably low [
75]. Interestingly, the hematological abnormalities caused by
Bartonella spp. were rarely described in naturally infected cats presented as healthy carriers [
11,
76]. For significantly raised MCV values, we suggested that its volume was increased from two main ways: (1) From multiplied daughter cells of
Bartonella in red blood cells in the replication stage [
11], and (2) from another previous infection or immune-mediated disorder [
28]. In addition, strong evidence suggested that heme compounds were necessary for
B. henselae growth and hemoglobin was a potential source of heme in vivo [
77] which explained the low values for HGB and MCHC in over 36% of
Bartonella-positive cats in this study. Nevertheless, there are some limitations including the criteria for owned animals’ inclusion. A significant inclusion criterion for blood donors of studied samples that might affect
Bartonella detection in owned animals required an ectoparasite control. Flea- and tick-controlled animals were possibly a chance of negative
Bartonella detection. Due to the ectoparasite infestation, there was a risk factor for
Bartonella infection in dogs and cats [
78]. However,
Bartonella detection in ectoparasite was not included in this study. For further study,
Bartonella spp. in animal-infesting ectoparasites should be examined to describe the dynamics of pathogen transmission.
4. Materials and Methods
4.1. Animal Ethics Consideration
Blood samples were collected by veterinarians and involved the gentle restraint technique. This animal research study was approved by the Kasetsart University Institutional Animal Care and Use Committee under the Ethical Review Board of the Office of National Research Council of Thailand (NRCT). All the laboratories used in this study met the standards followed by verification of the Institutional Biosafety Committee (IBC), Faculty of Veterinary Medicine, Kasetsart University (approval ID ACKU63-VET-048).
4.2. Definition of Surveyed Population
In this study, we surveyed both free-ranging and owned animals. Free-ranging animals were dogs and cats residing in temples of Bangkok. These animals were fed by animal care takers of each temple. Moreover, the ectoparasite control history of free-ranging animals was unknown. Owned animals were dogs and cats registered at the Blood Bank (BB) unit of Kasetsart University Veterinary Teaching Hospital (KUVTH) for blood donors. Owned animals lived only in the household and they roamed only in the area around their houses. The criteria for blood donors at BB of KUVTH included a limited age (1–7 years), flea and tick continue control, no blood receive history, no drug usage, and required body weight (dogs ≥ 17 kg and cats ≥ 4 kg).
4.3. Study Sites and Sample Collection
This cross-sectional study was conducted in the Bangkok metropolis of Thailand. The animal caretakers and owners of animals that resided in temples and registered with the BB at KUVTH, respectively were invited to participate in this study. Animal that had resided in temples were defined as free-ranging animals and animals that had donated blood at the BB of KUVTH were defined as owned animals. The temples that consented in this study were in three zones of Bangkok (
Figure 1). Thirty-four districts were intervened, and the number of animals per temples were based on the difficulty of handling and density of animals in the area. The data of the studied population are presented in
Supplementary Table S4. Blood samples from the free-ranging dogs were collected from the cephalic or saphenous vein depending on the size of the animal and in the donor dog samples were collected from the jugular vein. Additionally, cat samples consisted of punctured venous blood only from the jugular vein in both free-ranging and donor cats. Approximately 3 mL of blood was placed into a sterile blood collection tube with EDTA anticoagulant. Two hundred microliters of collected blood was separated for molecular detection of
Bartonella spp. and the remaining volume was used for hematological evaluation.
4.4. Animal’s General Data Collection
General data were collected for both free-ranging and owned animals including gender, living area, breed, age, and ectoparasite presentation at the surveyed time. Data of free-ranging animals were interviewed from the animal caretaker, however, data of owned animals were brought from the KUVTH record database and BB unit logbook. For ectoparasite presentation, they were examined at the time of blood collection and type of ectoparasite was recorded.
4.5. Genomic DNA Extraction
The separated blood samples were extracted for genomic DNA using commercial kits (FavorPrepTM Blood DNA Extraction Mini Kit, Favorgen Biotech Corporation, Pingtung, Taiwan) following the manufacturer’s instructions. The final volume of eluted solution (100 µL) was stored at −20 °C until the PCR-based detection.
4.6. PCR Quality Control
Each PCR reaction was conducted with negative and positive control samples. Nuclease-free water was used as the Bartonella negative control. B. henselae Houston-1 extracted DNA was provided from the National Institute of Health, Department of Medical Science, Ministry of Public Health, Thailand and used as the positive control sample.
4.7. Bartonella Screening Using PCR
Conventional PCR was performed to detect
Bartonella spp. in the extracted samples. First, all of the samples were screened for
Bartonella spp. using a primer set targeting a partial fragment of the citrate synthase (
gltA) gene (
Table 5). The amplification conditions were controlled by a thermocycler (Mastercycler
® nexus gradient, Eppendorf, Hamburg, Germany). The 25 µL of PCR mixture contained 0.5 µL of dNTPs solution (0.2 mM each), 1X of Taq Reaction buffer (mixed with MgSO
4), 4 pmol/µL of each primer, 0.04 U/µL of Taq DNA polymerase (Taq DNA Polymerase, Applied Biological Materials (ABM
®) Inc., Richmond, BC, Canada), 0.8% of dimethyl sulfoxide, and 3 µL of DNA template. The amplified products were kept at 4 °C until electrophoresis. Electrophoresis was conducted using agarose gel under a 0.5X TAE buffer. The process of electrophoresis was run at 100 V for 40–60 min depending on the expected amplicon size. The sample bands which were suspected as being positive for the
Bartonella amplicon size were purified using a DNA purification kit (Gel and PCR Purification System, BioFACT™, Daejeon, South Korea), according to the manufacturer’s instructions.
4.8. Other Housekeeping Gene Amplification
The
Bartonella-suspected samples were amplified for other housekeeping gene fragments (
Table 5) consisting of the beta-subunit of RNA polymerase (
rpoB), cell division protein (
ftsZ), 60 kDa chaperonin (
groEL), and riboflavin synthase (
ribC). The amplification cycles for each housekeeping gene are shown in
Table 6. Additionally, the PCR mixture, amplified amplicon, gel electrophoresis, and DNA purification were used as for the above description of
gltA.
4.9. DNA Sequencing and Phylogenetic Analyses
The purified amplicons were analyzed using Sanger’s sequencing technology by a commercial company (Macrogen
®, Seoul, Korea). The obtained DNA sequences were edited using the SnapGene
® Viewer software version 5.2.4 (
https://www.snapgene.com/snapgene-viewer, accessed on 20 March 2021) and the phylogenetic relationship was analyzed using the MEGA-X software (
https://www.megasoftware.net, accessed on 20 March 2021). Concatenated sequences were used to establish a phylogenetic tree under the fitted parameter model of the maximum likelihood method with 500 bootstrapping replications. All amplicon sequences were submitted to GenBank (accession numbers MW575344–MW575394).
4.10. Hematological Analyses
The remaining blood samples were sent to the Hematological Unit of KUVTH and to a private hematological company. All of the blood samples were analyzed using the laser flow cytometry technique by an automated hematology analyzer (Sysmex XT-2000iV™, IDEXX Bioresearch, Norderstedt, Germany). Additionally, a manual differential count from the Diff-Quick stained blood smear was also performed in all of the samples. Common blood panels were analyzed based on the leukocyte count (WBC) and the proportions of neutrophils (NEU), lymphocytes (LYM), eosinophils (EOS), monocytes (MON), and by the numbers of basophils (BAS), the erythrocytic count (RBC), hemoglobin concentration (HGB), packed cell volume (PCV), mean corpuscular volume (MCV), mean corpuscular hemoglobin (MCH), mean corpuscular hemoglobin concentration (MCHC), and platelet count (PLT).
4.11. Statistical Analyses
General data on the tested animals were recorded for age, sex, breed, roaming/living location, ectoparasitic infestation, and living pattern. The general and hematological data were statistically analyzed using the R programming language version 4.0.2. Variables of interest were tested as risk factors using Chi square or Fisher’s exact test depending on the nature of the recorded data. Only the significant factors were analyzed, and a multiple comparison used the Bonferroni correction method and calculated odds ratio. A comparison of hematological values was tested between Bartonella-positive and Bartonella-negative groups using the Mann-Whitney U test. All statistical analyses were processed using 95% confidence intervals and a p-value < 0.05 was considered as significant.








{kind=link}
{kind=link}
{kind=link}