
Interferon Gamma Inhibits Equine Herpesvirus 1 Replication in a Cell Line-Dependent Manner

Abstract
:1. Introduction
2. Results
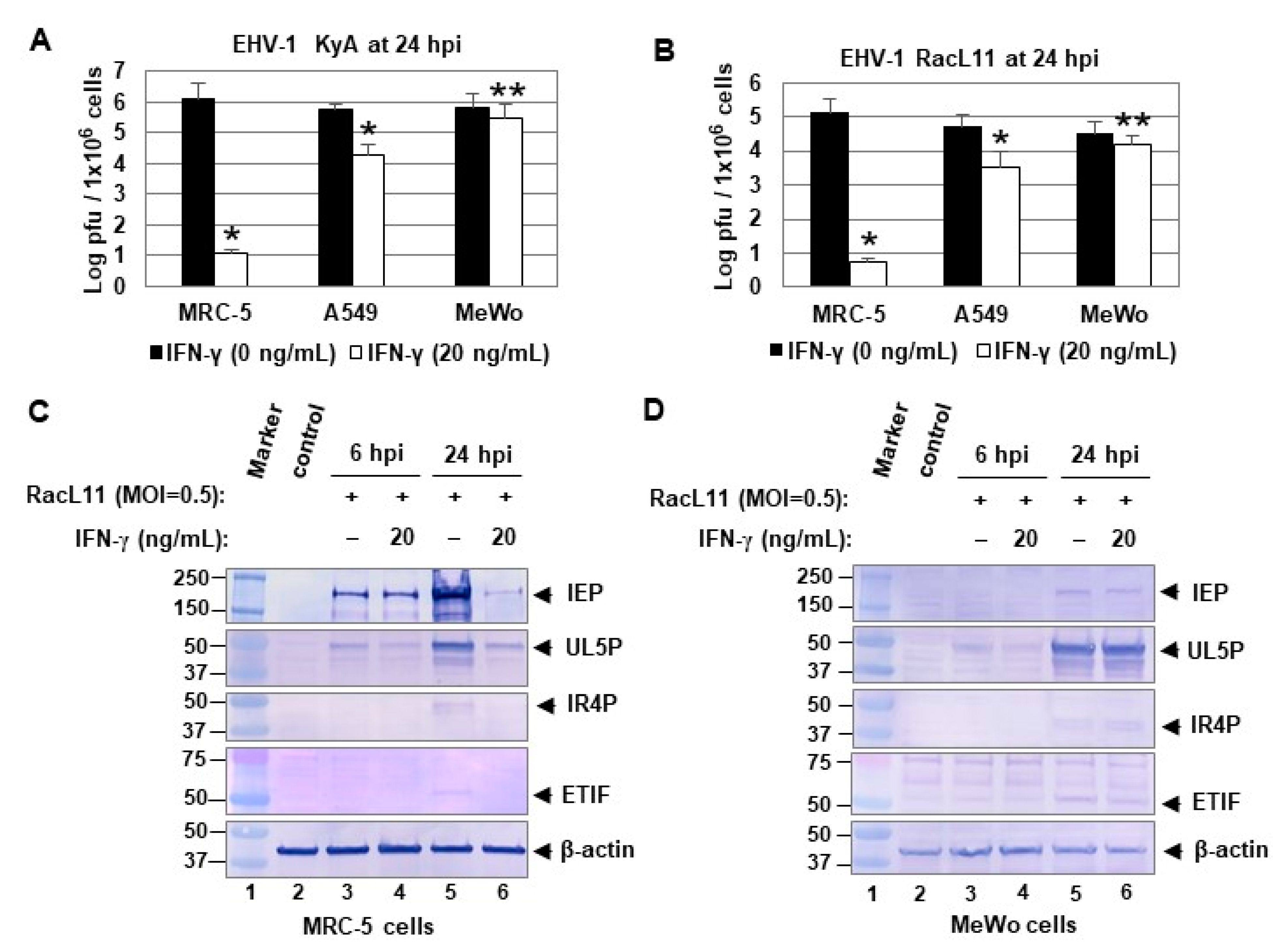
2.1. IFN-γ Treatment Inhibits EHV-1 Replication in a Cell Line-Dependent Manner
2.2. IFN-γ Blocks the IEP-Mediated Trans-Activation by Inhibiting the Expression of IEP in MH-S Cells
2.3. EHV-1 Inhibition by IFN-γ in MH-S, NBL6, and MRC-5 Cells Is Not Due to Nitric Oxide
2.4. IFN-γ Inhibits EHV-1 Replication in MRC-5 Cells
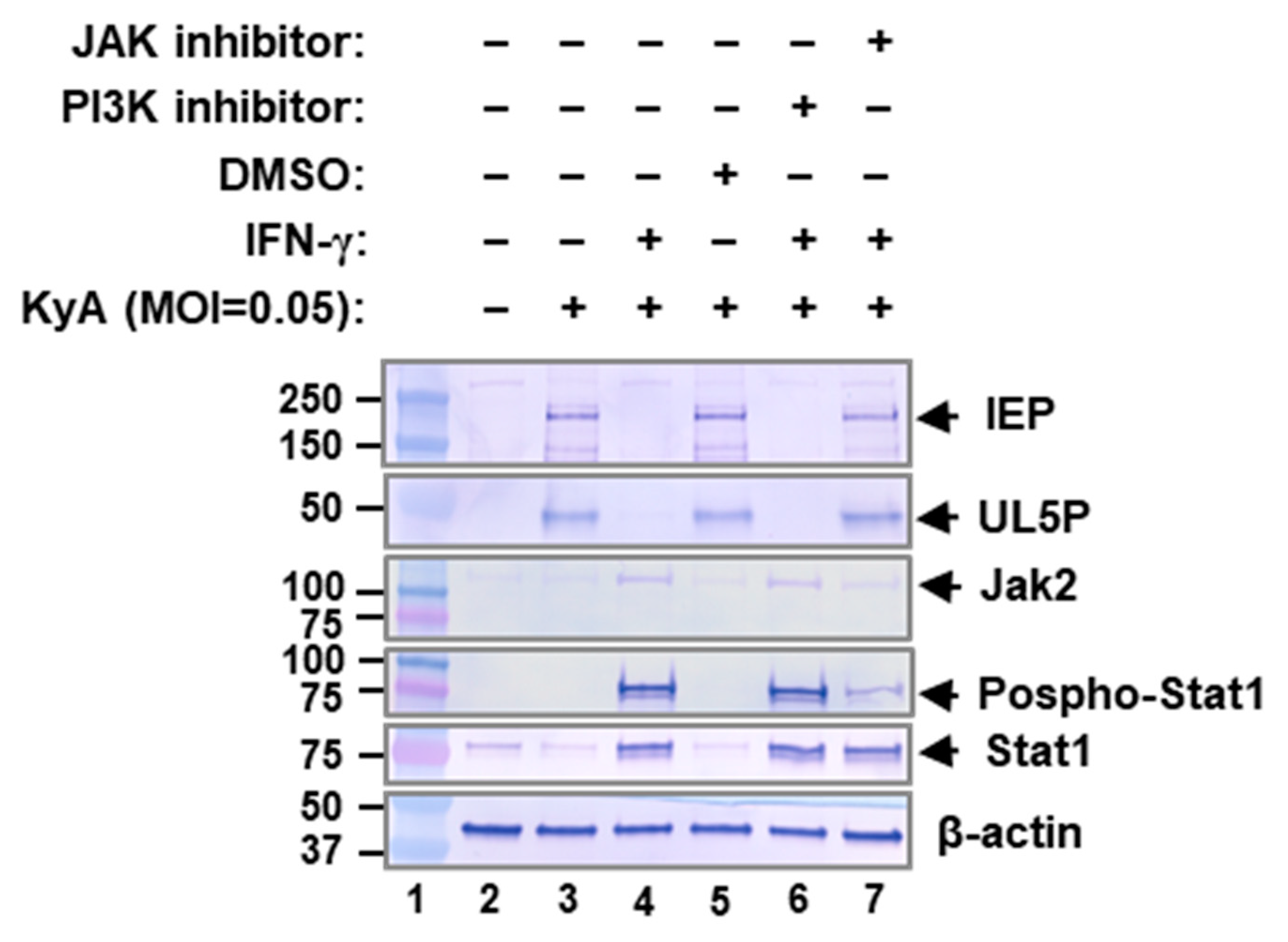
2.5. JAK/STAT1 Signaling Pathway Mediates EHV-1 Inhibition
2.6. Innate Immunity Related Genes and Antiviral Interferon-Stimulated Genes Are Upregulated in the IFN-γ-Treated MH-S Cells
3. Discussion
4. Materials and Methods
4.1. Viruses and Cell Culture
4.2. Plaque Assays
4.3. Western Blot Analysis
4.4. Luciferase Reporter and Mammalian Expression Plasmids
4.5. Luciferase Reporter Assays
4.6. Nitric Oxide (NO) Assay
4.7. Microarray Analysis
4.8. Microarray Data Accession Number
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Allen, G.; Bryans, J. Molecular epizootiology, pathogenesis, and prophylaxis of equine herpesvirus-1 infections. Prog. Vet. Microbiol. Immunol. 1985, 2, 78–144. [Google Scholar]
- Carroll, C.; Westbury, H. Isolation of equine herpesvirus 1 from the brain of a horse affected with paresis. Aust. Vet. J. 1985, 62, 345–346. [Google Scholar] [CrossRef]
- Jackson, T.; Kendrick, J. Paralysis of horses associated with equine herpesvirus 1 infection. J. Am. Vet. Med. Ass. J. 1971, 158, 1351–1357. [Google Scholar]
- Bryans, J.T.; Allen, G.P. Equine viral rhinopneumonitis. Rev. Sci. Tech. Off. Int. Epizoot. 1986, 5, 837–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crandell, R.; Mock, R.; Lock, T. Vaccination of pregnant ponies against equine rhinopneumonitis. Am. J. Vet. Res. 1980, 41, 994–996. [Google Scholar]
- Farrar, M.A.; Schreiber, R.D. The molecular cell biology of interferon-gamma and its receptor. Annu. Rev. Immunol. 1993, 11, 571–611. [Google Scholar] [CrossRef]
- Schoenborn, J.R.; Wilson, C.B. Regulation of interferon-γ during innate and adaptive immune responses. Adv. Immunol. 2007, 96, 41–101. [Google Scholar] [PubMed]
- Spellberg, B.; Edwards, J.E. Type 1/Type 2 immunity in infectious diseases. Clin. Infect. Dis. 2001, 32, 76–102. [Google Scholar] [CrossRef] [PubMed]
- Ealick, S.E.; Cook, W.J.; Vijay-Kumar, S.; Carson, M.; Nagabhushan, T.L.; Trotta, P.P.; Bugg, C.E. Three-dimensional structure of recombinant human interferon-gamma. Science 1991, 252, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Platanias, L.C. Mechanisms of type-I-and type-II-interferon-mediated signalling. Nat. Rev. Immunol. 2005, 5, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Haan, C.; Kreis, S.; Margue, C.; Behrmann, I. Jaks and cytokine receptors—An intimate relationship. Biochem. Pharmacol. 2006, 72, 1538–1546. [Google Scholar] [CrossRef] [PubMed]
- Rodig, S.J.; Meraz, M.A.; White, J.M.; Lampe, P.A.; Riley, J.K.; Arthur, C.D.; King, K.L.; Sheehan, K.C.; Yin, L.; Pennica, D. Disruption of the Jak1 gene demonstrates obligatory and nonredundant roles of the Jaks in cytokine-induced biologic responses. Cell 1998, 93, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Yeh, T.; Pellegrini, S. The Janus kinase family of protein tyrosine kinases and their role in signaling. Cell. Mol. Life Sci. 1999, 55, 1523–1534. [Google Scholar] [CrossRef] [PubMed]
- Boehm, U.; Klamp, T.; Groot, M.; Howard, J. Cellular responses to interferon-gamma. Annu. Rev. Immunol. 1997, 15, 749–795. [Google Scholar] [CrossRef]
- Colle, C.F., III; Flowers, C.C.; O’Callaghan, D.J. Open reading frames encoding a protein kinase, homolog of glycoprotein gX of pseudorabies virus, and a novel glycoprotein map within the unique short segment of equine herpesvirus type 1. Virology 1992, 188, 545–557. [Google Scholar] [CrossRef]
- Frampton, A.R.; Smith, P.M.; Zhang, Y.; Matsumura, T.; Osterrieder, N.; O’Callaghan, D.J. Contribution of gene products encoded within the unique short segment of equine herpesvirus 1 to virulence in a murine model. Virus Res. 2002, 90, 287–301. [Google Scholar] [CrossRef]
- Lewis, J.B.; Thompson, Y.G.; Feng, X.; Holden, V.R.; O’callaghan, D.; Caughman, G.B. Structural and antigenic identification of the ORF12 protein (αTIF) of equine herpesvirus 1. Virology 1997, 230, 369–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumura, T.; O’Callaghan, D.; Kondo, T.; Kamada, M. Lack of virulence of the murine fibroblast adapted strain, Kentucky A (KyA), of equine herpesvirus type 1 (EHV-1) in young horses. Vet. Microbiol. 1996, 48, 353–365. [Google Scholar] [CrossRef]
- Smith, P.M.; Kahan, S.M.; Rorex, C.B.; von Einem, J.; Osterrieder, N.; O’Callaghan, D.J. Expression of the full-length form of gp2 of equine herpesvirus 1 (EHV-1) completely restores respiratory virulence to the attenuated EHV-1 strain KyA in CBA mice. J. Virol. 2005, 79, 5105–5115. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.M.; Zhang, Y.; Grafton, W.D.; Jennings, S.R.; O’Callaghan, D.J. Severe murine lung immunopathology elicited by the pathogenic equine herpesvirus 1 strain RacL11 correlates with early production of macrophage inflammatory proteins 1α, 1β, and 2 and tumor necrosis factor alpha. J. Virol. 2000, 74, 10034–10040. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.K.; Shakya, A.K.; O’Callaghan, D.J. Immunization with attenuated equine herpesvirus 1 strain KyA induces innate immune responses that protect mice from lethal challenge. J. Virol. 2016, 90, 8090–8104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coombs, D.K.; Patton, T.; Kohler, A.K.; Soboll, G.; Breathnach, C.; Townsend, H.G.; Lunn, D.P. Cytokine responses to EHV-1 infection in immune and non-immune ponies. Vet. Immunol. Immunopathol. 2006, 111, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Bartels, T.; Steinbach, F.; Hahn, G.; Ludwig, H.; Borchers, K. In situ study on the pathogenesis and immune reaction of equine herpesvirus type 1 (EHV-1) infections in mice. Immunology 1998, 93, 329–334. [Google Scholar] [CrossRef]
- Kim, S.K.; Shakya, A.K.; O’Callaghan, D.J. Intranasal treatment with CpG-B oligodeoxynucleotides protects CBA mice from lethal equine herpesvirus 1 challenge by an innate immune response. Antivir. Res. 2019, 169, 104546. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.H.; Caughman, G.B.; O’Callaghan, D.J. Characterization of the regulatory functions of the equine herpesvirus 1 immediate-early gene product. J. Virol. 1992, 66, 936–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.H.; Holden, V.R.; O’Callaghan, D.J. Nuclear localization and transcriptional activation activities of truncated versions of the immediate-early gene product of equine herpesvirus 1. J. Virol. 1995, 69, 3857–3862. [Google Scholar] [CrossRef] [Green Version]
- Buczynski, K.A.; Kim, S.K.; O’Callaghan, D.J. Initial characterization of 17 viruses harboring mutant forms of the immediate-early gene of equine herpesvirus 1. Virus Genes 2005, 31, 229–239. [Google Scholar] [CrossRef]
- Harris, N.; Buller, R.M.; Karupiah, G. Gamma interferon-induced, nitric oxide-mediated inhibition of vaccinia virus replication. J. Virol. 1995, 69, 910–915. [Google Scholar] [CrossRef] [Green Version]
- Harty, R.N.; O’Callaghan, D.J. An early gene maps within and is 3′ coterminal with the immediate-early gene of equine herpesvirus 1. J. Virol. 1991, 65, 3829–3838. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.K.; Shakya, A.K.; O’Callaghan, D.J. Full trans-activation mediated by the immediate-early protein of equine herpesvirus 1 requires a consensus TATA box, but not its cognate binding sequence. Virus Res. 2016, 211, 222–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.H.; Zhao, Y.; O’Callaghan, D.J. The equine herpesvirus type 1 immediate-early gene product contains an acidic transcriptional activation domain. Virology 1994, 202, 760–770. [Google Scholar] [CrossRef]
- Caughman, G.B.; Staczek, J.; O’Callaghan, D.J. Equine herpesvirus type 1 infected cell polypeptides: Evidence for immediate early/early/late regulation of viral gene expression. Virology 1985, 145, 49–61. [Google Scholar] [CrossRef]
- Gray, W.L.; Baumann, R.P.; Robertson, A.T.; Caughman, G.B.; O’Callaghan, D.J.; Staczek, J. Regulation of equine herpesvirus type 1 gene expression: Characterization of immediate early, early, and late transcription. Virology 1987, 158, 79–87. [Google Scholar] [CrossRef]
- Gray, W.L.; Baumann, R.P.; Robertson, A.T.; O’Callaghan, D.J.; Staczek, J. Characterization and mapping of equine herpesvirus type 1 immediate early, early, and late transcripts. Virus Res. 1987, 8, 233–244. [Google Scholar] [CrossRef]
- Henry, B.E.; Robinson, R.A.; Dauenhauer, S.A.; Atherton, S.S.; Hayward, G.S.; O’Callaghan, D.J. Structure of the genome of equine herpesvirus type 1. Virology 1981, 115, 97–114. [Google Scholar] [CrossRef]
- Shakya, A.K.; O’Callaghan, D.J.; Kim, S.K. Interferon gamma inhibits varicella-zoster virus replication in a cell line-dependent manner. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodwin, M.M.; Canny, S.; Steed, A.; Virgin, H.W. Murine gammaherpesvirus 68 has evolved gamma interferon and stat1-repressible promoters for the lytic switch gene 50. J. Virol. 2010, 84, 3711–3717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoggins, J.W.; Rice, C.M. Interferon-stimulated genes and their antiviral effector functions. Curr. Opin. Virol. 2011, 1, 519–525. [Google Scholar] [CrossRef]
- Chen, S.; Bonifati, S.; Qin, Z.; Gelais, C.S.; Kodigepalli, K.M.; Barrett, B.S.; Kim, S.H.; Antonucci, J.M.; Ladner, K.J.; Buzovetsky, O. SAMHD1 suppresses innate immune responses to viral infections and inflammatory stimuli by inhibiting the NF-κB and interferon pathways. Proc. Natl. Acad. Sci. USA 2018, 115, E3798–E3807. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Zhu, M.; Pan, X.; Zhu, Y.; Yan, H.; Jiang, T.; Shen, Y.; Dong, X.; Zheng, N.; Lu, J. Inhibition of Hepatitis B virus replication by SAMHD1. Biochem. Biophys. Res. Commun. 2014, 450, 1462–1468. [Google Scholar] [CrossRef]
- Sze, A.; Belgnaoui, S.M.; Olagnier, D.; Lin, R.; Hiscott, J.; van Grevenynghe, J. Host restriction factor SAMHD1 limits human T cell leukemia virus type 1 infection of monocytes via STING-mediated apoptosis. Cell Host Microbe 2013, 14, 422–434. [Google Scholar] [CrossRef] [Green Version]
- Wagner, B.; Robeson, J.; McCracken, M.; Wattrang, E.; Antczak, D.F. Horse cytokine/IgG fusion proteins—Mammalian expression of biologically active cytokines and a system to verify antibody specificity to equine cytokines. Vet. Immunol. Immunopathol. 2005, 105, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Gutmann, S.; Zawatzky, R.; Müller, M. Characterisation and quantification of equine interferon gamma. Vet. Immunol. Immunopathol. 2005, 104, 105–115. [Google Scholar] [CrossRef]
- Smith, P.M.; Zhang, Y.; Jennings, S.R.; O’Callaghan, D.J. Characterization of the cytolytic T-lymphocyte response to a candidate vaccine strain of equine herpesvirus 1 in CBA mice. J. Virol. 1998, 72, 5366–5372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Smith, P.M.; Jennings, S.R.; O’Callaghan, D.J. Quantitation of virus-specific classes of antibodies following immunization of mice with attenuated equine herpesvirus 1 and viral glycoprotein D. Virology 2000, 268, 482–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colle, C.F.; Tarbet, E.B.; Grafton, W.D.; Jennings, S.R.; O’Callaghan, D.J. Equine herpesvirus-1 strain KyA, a candidate vaccine strain, reduces viral titers in mice challenged with a pathogenic strain, RacL. Virus Res. 1996, 43, 111–124. [Google Scholar] [CrossRef]
- Ito, S.; Ishii, K.J.; Gursel, M.; Shirotra, H.; Ihata, A.; Klinman, D.M. CpG oligodeoxynucleotides enhance neonatal resistance to Listeria infection. J. Immunol. 2005, 174, 777–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wattrang, E.; Palm, A.-K.; Wagner, B. Cytokine production and proliferation upon in vitro oligodeoxyribonucleotide stimulation of equine peripheral blood mononuclear cells. Vet. Immunol. Immunopathol. 2012, 146, 113–124. [Google Scholar] [CrossRef]
- Holden, V.R.; Yalamanchili, R.R.; Harty, R.N.; O’Callaghan, D. ICP22 homolog of equine herpesvirus 1: Expression from early and late promoters. J. Virol. 1992, 66, 664–673. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.K.; Ahn, B.C.; Albrecht, R.A.; O’Callaghan, D.J. The unique IR2 protein of equine herpesvirus 1 negatively regulates viral gene expression. J. Virol. 2006, 80, 5041–5049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Holden, V.R.; Smith, R.H.; O’Callaghan, D.J. Regulatory function of the equine herpesvirus 1 ICP27 gene product. J. Virol. 1995, 69, 2786–2793. [Google Scholar] [CrossRef] [Green Version]
- Roizman, B. The family Herpesviridae: A brief introduction. Fields Virol. 2001, 2, 2381–2397. [Google Scholar]
- Nathan, C. Nitric oxide as a secretory product of mammalian cells. FASEB J. 1992, 6, 3051–3064. [Google Scholar] [CrossRef]
- Nathan, C.F.; Hibbs, J.B., Jr. Role of nitric oxide synthesis in macrophage antimicrobial activity. Curr. Opin. Immunol. 1991, 3, 65–70. [Google Scholar] [CrossRef]
- Karupiah, G.; Xie, Q.W.; Buller, R.M.; Nathan, C.; Duarte, C.; MacMicking, J.D. Inhibition of viral replication by interferon-gamma-induced nitric oxide synthase. Science 1993, 261, 1445–1448. [Google Scholar] [CrossRef] [PubMed]
- Trilling, M.; Le, V.T.; Zimmermann, A.; Ludwig, H.; Pfeffer, K.; Sutter, G.; Smith, G.L.; Hengel, H. Gamma interferon-induced interferon regulatory factor 1-dependent antiviral response inhibits vaccinia virus replication in mouse but not human fibroblasts. J. Virol. 2009, 83, 3684–3695. [Google Scholar] [CrossRef] [Green Version]
- Der, S.D.; Zhou, A.; Williams, B.R.; Silverman, R.H. Identification of genes differentially regulated by interferon α, β, or γ using oligonucleotide arrays. Proc. Natl. Acad. Sci. USA 1998, 95, 15623–15628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, G.C.; Peters, G.A. Viral stress-inducible genes. Adv. Virus Res. 2007, 70, 233–263. [Google Scholar]
- O’Callaghan, D.J.; Cheevers, W.P.; Gentry, G.A.; Randall, C.C. Kinetics of cellular and viral DNA synthesis in equine abortion (herpes) virus infection of L-M cells. Virology 1968, 36, 104–114. [Google Scholar] [CrossRef]
- Perdue, M.L.; Kemp, M.C.; Randall, C.C.; O’Callaghan, D.J. Studies of the molecular anatomy of the L-M cell strain of equine herpes virus type 1: Proteins of the nucleocapsid and intact virion. Virology 1974, 59, 201–216. [Google Scholar] [CrossRef]
- Reczko, E.; Mayr, A. On the fine structure of a virus of the herpes group isolated from horses (short report). Arch. Für Die Gesamte Virusforsch. 1963, 13, 591. [Google Scholar] [CrossRef]
- Kim, S.K.; Kim, S.; Dai, G.; Zhang, Y.; Ahn, B.C.; O’Callaghan, D.J. Identification of functional domains of the IR2 protein of equine herpesvirus 1 required for inhibition of viral gene expression and replication. Virology 2011, 417, 430–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Fritsh, E.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Press: New York, NY, USA, 1989. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Function and Name | Fold Change a | Description | |
|---|---|---|---|
| MH-S | L-M | ||
| Immune and inflammatory responses | |||
| C1qb | 38.1 | ― | Complement component 1, q subcomponent, beta polypeptide |
| Mx1 | 18.1 | ― | MX dynamin-like GTPase 1 |
| Irgm1 | 7 | 20.2 | Immunity-related GTPase family M member 1 |
| Fcgr1 | 6.1 | ― | Fc receptor, IgG, high affinity I |
| C1qa | 6.6 | ― | Complement component 1, q subcomponent, alpha polypeptide |
| Samhd1 | 5.9 | ― | SAM domain and HD domain, 1 |
| Il18rap | 4 | ― | Interleukin 18 receptor accessory protein |
| Tlr3 | 3.9 | ― | Toll-like receptor 3 |
| C3 | 3.8 | ― | Complement component 3 |
| C1qc | 3.5 | ― | Complement component 1, q subcomponent, C chain |
| Tmem173 | 2.9 | ― | Transmembrane protein 173 |
| Cfb | 2.8 | ― | Complement factor B |
| Ifih1 | 2.6 | 4.3 | Interferon induced with helicase C domain 1 |
| Clec7a | 2.2 | ― | C-type lectin domain family 7, member a |
| Ciita | 2.1 | ― | Class II transactivator |
| Tnfaip3 | 2.1 | ― | Tumor necrosis factor, alpha-induced protein 3 |
| Tnfrsf1a | 2 | ― | Tumor necrosis factor receptor superfamily, member 1a |
| CC chemokines and Chemokine receptors | |||
| Cxcl9 | 431.9 | ― | Chemokine (C-X-C motif) ligand 9 |
| Cxcl11 | 119.7 | ― | Chemokine (C-X-C motif) ligand 11 |
| Cxcl10 | 17.6 | ― | Chemokine (C-X-C motif) ligand 10 |
| Ccl12 | 14.6 | ― | Chemokine (C-C motif) ligand 12 |
| Ccl8 | 7.5 | ― | Chemokine (C-C motif) ligand 8 |
| Ccrl2 | 4.5 | ― | Chemokine (C-C motif) receptor-like 2 |
| Ccl7 | 3.2 | ― | Chemokine (C-C motif) ligand 7 |
| Cytokines and cytokine receptors | |||
| Fas | 10.4 | ― | TNF receptor superfamily member 6 |
| Tnfaip2 | 5 | 3.5 | Tumor necrosis factor, alpha-induced protein 2 |
| Il15 | 4.4 | 3.1 | Interleukin 15 |
| Tnfsf10 | 4 | 3.8 | Tumor necrosis factor superfamily, member 10 |
| Il15ra | 4.3 | 2.8 | Interleukin 15 receptor, alpha chain |
| Il12rb1 | 4.3 | 2.5 | Interleukin 12 receptor, beta 1 |
| Tnfrsf14 | 3.9 | 4.6 | Tumor necrosis factor receptor superfamily, member 14 |
| Il18rap | 4 | ― | Interleukin 18 receptor accessory protein |
| Il18bp | 3.6 | 4.3 | Interleukin 18 binding protein |
| l15ra | 4.3 | 2.8 | Interleukin 15 receptor, alpha chain |
| Il13ra1 | 2.7 | ― | Interleukin 13 receptor, alpha 1 |
| Il18 | 2.8 | ― | Interleukin 18 |
| Il10ra | 2.6 | ― | Interleukin 10 receptor, alpha |
| Tnf | 2.5 | ― | Tumor necrosis factor |
| Tnfsf13b | 2 | 3.1 | Tumor necrosis factor superfamily, member 13b |
| Miscellaneous | |||
| SOCS1 | 7.4 | 7.4 | Suppressor of cytokine signalling-1 |
| TLR3 | 4.3 | 2 | Toll-like receptor 3 |
| TAT1 | 5.3 | 3.1 | Signal transducer and activator of transcription 1 |
| STAT3 | 2.3 | 3.1 | Signal transducer and activator of transcription 3 |
| Gene Name | Fold Change a | Target Viruses b | |
|---|---|---|---|
| MH-S | L-M | ||
| GBP1 | 143 | 285 | EMCV, HCV(r), VSV |
| GBP 2 | 111.6 | 260.7 | EMCV, HCV(r), VSV |
| IRF1 | 18.5 | 13.2 | Numerous RNA and DNA viruses |
| MX1 | 18.1 | ― | CVB, FLUAV, HCV(r), HPIV3, LACV, MV, SFV, VSV |
| SAMHD1 | 5.9 | ― | HIV, SeV, HBV, HTLV1 |
| IFIT2 | 5.3 | ― | FLUAV, HPV, MHV, RVFV, SINV, VSV, WNV |
| NAMPT | 4.6 | ― | VEEV, WNV |
| RSAD2 (viperin) | 4 | 8.6 | DENV, DENV(v), FLUAV, HCMV, HCV(r), SINV |
| TREX1 | 4 | ― | YFN |
| IFIT1 | 3.5 | 2.1 | FLUAV, HPV, MHV, RVFV, SINV, VSV, WNV |
| DDX60 | 3.2 | ― | HCV, PV |
| IFIH1 (MDA5) | 2.6 | 4.3 | Numerous RNA and DNA viruses |
| OASL2 | 2.5 | 15.4 | HCV |
| IRF7 | 2.4 | 3.1 | Numerous RNA and DNA viruses |
| ISG20 | 2.3 | 2.9 | FLUAV, HIV-1, HSV-1, JEV, MHV-68, SINV |
| EIF2AK2 (PKR) | 2 | 2.4 | Numerous RNA and DNA viruses |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.K.; Shakya, A.K.; O’Callaghan, D.J. Interferon Gamma Inhibits Equine Herpesvirus 1 Replication in a Cell Line-Dependent Manner. Pathogens 2021, 10, 484. https://doi.org/10.3390/pathogens10040484
Kim SK, Shakya AK, O’Callaghan DJ. Interferon Gamma Inhibits Equine Herpesvirus 1 Replication in a Cell Line-Dependent Manner. Pathogens. 2021; 10(4):484. https://doi.org/10.3390/pathogens10040484
Chicago/Turabian StyleKim, Seong K., Akhalesh K. Shakya, and Dennis J. O’Callaghan. 2021. "Interferon Gamma Inhibits Equine Herpesvirus 1 Replication in a Cell Line-Dependent Manner" Pathogens 10, no. 4: 484. https://doi.org/10.3390/pathogens10040484