
Autochthonous Trypanosoma spp. in European Mammals: A Brief Journey amongst the Neglected Trypanosomes


Abstract
:
1. Introduction
2. General Taxonomy of the Genus Trypanosoma
3. Trypanosoma Species Naturally Occurring in Domestic and Wild Mammals in Europe
3.1. Trypanosoma theileri
3.2. Trypanosoma melophagium
3.3. Trypanosoma lewisi
3.4. Trypanosoma nabiasi
3.5. Trypanosomes of Small Rodents
3.6. Trypanosomes of Bats
3.7. Trypanosoma pestenai
3.8. Trypanosoma spp.
4. Closing Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Votýpka, J.; d’Avila-Levy, C.M.; Grellier, P.; Maslov, D.; Lukeš, J.; Yurchenko, V. New Approaches to Systematics of Trypanosomatidae: Criteria for Taxonomic (Re)description. Trends Parasitol. 2015, 31, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Hoare, C.A. The Trypanosomes of Mammals, 1st ed.; Blackwell Scientific Publications: Oxford, UK; Edinburgh, UK, 1972; pp. 125–140, 214–219, 219–245, 277–282. [Google Scholar]
- Radwanska, M.; Vereecke, N.; Deleeuw, V.; Pinto, J.; Magez, S. Salivarian trypanosomosis: A review of parasites involved, their global distribution and their interaction with the innate and adaptive mammalian host immune system. Front. Immunol. 2018, 9, 2253. [Google Scholar] [CrossRef] [PubMed]
- Linder, A.K.; Priotto, G. The Unknown Risk of Vertical Transmission in Sleeping Sickness—A Literature Review. PLoS Negl. Trop. Dis. 2010, 4, e783. [Google Scholar] [CrossRef] [Green Version]
- Brun, R.; Blum, J.; Chappuis, F.; Burri, C. Human African trypanosomiasis. Lancet 2010, 375, 148–159. [Google Scholar] [CrossRef] [Green Version]
- Blum, J.A.; Neumayr, A.L.; Hatz, C.F. Human African trypanosomiasis in endemic populations and travelers. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 905–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashcroft, M.T. The importance of African wild animals as reservoirs of trypanosomiasis. East Afr. Med. J. 1959, 36, 289–297. [Google Scholar]
- Auty, H.; Anderson, N.E.; Picozzi, K.; Lembo, T.; Mubanga, J.; Hoare, R.; Fyumagwa, R.D.; Mable, B.; Hamill, L.; Cleaveland, S.; et al. Trypanosome Diversity in Wildlife Species from the Serengeti and Luangwa Valley Ecosystems. PLoS Negl. Trop. Dis. 2012, 6, e1828. [Google Scholar] [CrossRef] [Green Version]
- Junyent, J.M.G.; Rozman, M.; Corachan, M.; Estruch, R.; Urbano-Marquez, A. An unusual course of west African trypanosomiasis in a Caucasian man. Trans. R. Soc. Trop. Med. Hyg. 1987, 81, 931–932. [Google Scholar] [CrossRef]
- Braendli, B.; Dankwa, E.; Junghanss, T. East African sleeping sickness (Trypanosoma rhodesiense infection) in 2 Swiss travelers to the tropics. Swiss Med. Wkly. 1990, 120, 1348–1352. [Google Scholar]
- Iborra, C.; Danis, M.; Bricaire, F.; Caumes, E.A. Traveler Returning from Central Africa with Fever and a Skin Lesion. Clin. Infect. Dis. 1999, 28, 679–680. [Google Scholar] [CrossRef] [Green Version]
- Jamonneau, V.; Garcia, A.; Ravel, S.; Cuny, G.; Oury, B.; Solano, P.; N’Guessan, P.; N’Dri, L.; Sanon, R.; Frézil, J.L.; et al. Genetic characterization of Trypanosoma brucei gambiense and clinical evolution of human African trypanosomiasis in Côte d’Ivoire. Trop. Med. Int. Health 2002, 7, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Ripamonti, D.; Massari, M.; Arici, C.; Gabbi, E.; Farina, C.; Brini, M.; Capatti, C.; Suter, F. African sleeping sickness in tourists returning from Tanzania: The first 2 Italian cases from a small outbreak among European travelers. Clin. Infect. Dis. 2002, 34, 18–22. [Google Scholar] [CrossRef] [Green Version]
- Bisoffi, Z.; Beltrame, A.; Monteiro, G.; Arzese, A.; Marocco, S.; Rorato, G.; Anselmi, M.; Viale, P. African Trypanosomiasis Gambiense, Italy. Emerg. Infect. Dis. 2005, 11, 1745–1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautret, P.; Clerinx, J.; Caumes, E.; Simon, F.; Jensenius, M.; Loutan, L.; Shlagenhauf, P.; Castelli, F.; Freedman, D.; Miller, A.; et al. Imported human African trypanosomiasis in Europe, 2005–2009. Eurosurveillance 2010, 14, 19327. [Google Scholar] [CrossRef]
- Gómez-Junyent, J.; Pinazo, M.J.; Castro, P.; Fernández, S.; Mas, J.; Chaguaceda, C.; Pellicé, M.; Muñoz, J. Human African Trypanosomiasis in a Spanish traveler returning from Tanzania. PLoS Negl. Trop. Dis. 2017, 11, e0005324. [Google Scholar] [CrossRef] [Green Version]
- Huits, R.; De Ganck, G.; Clerinx, J.; Büscher, P.; Bottieau, E. A veterinarian with fever, rash and chancre after holidays in Uganda. J. Travel Med. 2018, 25, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.-M.; Qian, Z.-Y.; Hide, G.; Lai, D.-H.; Lun, Z.-R.; Wu, Z.-D. Human African trypanosomiasis: The current situation in endemic regions and the risks for nonendemic regions from imported cases. Parasitology 2020, 147, 922–931. [Google Scholar] [CrossRef] [PubMed]
- Simarro, P.P.; Franco, J.R.; Cecchi, G.; Paone, M.; Diarra, A.; Postigo, J.A.R.; Jannin, J.G. Human African Trypanosomiasis in Non-Endemic Countries. J. Travel Med. 2012, 19, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Angheben, A. Chagas Disease in the Mediterranean Area. Curr. Trop. Med. Rep. 2017, 4, 223–234. [Google Scholar] [CrossRef]
- Coura, J.R. The main sceneries of Chagas disease transmission. The vectors, blood and oral transmissions—A comprehensive review. Mem. Inst. Oswaldo Cruz 2015, 110, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Jansen, A.M.; Xavier, S.C.C.; Roque, A.L.R. Trypanosoma cruzi transmission in the wild and its most important reservoir hosts in Brazil. Parasites Vectors 2018, 11, 502. [Google Scholar] [CrossRef]
- Jansen, A.M.; Xavier, S.C.C.; Roque, A.L.R. Landmarks of the Knowledge and Trypanosoma cruzi Biology in the Wild Environment. Front. Cell. Infect. Microbiol. 2020, 10, 10. [Google Scholar] [CrossRef]
- Angheben, A.; Anselmi, M.; Gobbi, F.; Marocco, S.; Monteiro, G.; Buonfrate, D.; Tais, S.; Talamo, M.; Zavarise, G.; Strohmeyer, M.; et al. Chagas disease in Italy: Breaking an epidemiological silence. Eurosurveillance 2011, 16, 19969. [Google Scholar] [CrossRef] [Green Version]
- Gobbi, F.; Angheben, A.; Anselmi, M.; Postiglione, C.; Repetto, E.; Buonfrate, D.; Marocco, S.; Tais, S.; Chiampan, A.; Mainardi, P.; et al. Profile of Trypanosoma cruzi Infection in a Tropical Medicine Reference Center, Northern Italy. PLoS Negl. Trop. Dis. 2014, 8, e3361. [Google Scholar] [CrossRef]
- Repetto, E.C.; Zachariah, R.; Kumar, A.; Angheben, A.; Gobbi, F.; Anselmi, M.; Al Rousan, A.; Toricco, C.; Ruiz, R.; Ledezma, G.; et al. Neglect of a Neglected Disease in Italy: The Challenge of Access-to-Care for Chagas Disease in Bergamo Area. PLoS Negl. Trop. Dis. 2015, 9, e0004103. [Google Scholar] [CrossRef]
- Antinori, S.; Galimberti, L.; Bianco, R.; Grande, R.; Galli, M.; Corbellino, M. Chagas disease in Europe: A review for the internist globalized world. Eur. J. Intern. Med. 2017, 43, 6–15. [Google Scholar] [CrossRef]
- Basile, L.; Jansà, J.M.; Carlier, Y.; Salamanca, D.D.; Angheben, A.; Bartoloni, A.; Seixas, J.; Van Gool, T.; Cañavate, C.; Flores-Chávez, M.; et al. Chagas disease in European countries: The challenge of a surveillance system. Eurosurveillance 2011, 16, 19968. [Google Scholar] [CrossRef] [Green Version]
- Taylor, D.B. Stable Fly (Stomoxys calcitrans). In Reference Module in Biomedical Science; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar] [CrossRef]
- Jones, T.W.; Dávila, A.M. Trypanosoma vivax—Out of Africa. Trends Parasitol. 2001, 17, 99–101. [Google Scholar] [CrossRef]
- Chamond, N.; Cosson, A.; Blom-Potar, M.C.; Jouvion, G.; D’Archivio, S.; Medina, M.; Droin-Bergère, S.; Huerre, M.; Minoprio, P. Trypanosoma vivax Infections: Pushing Ahead with Mouse Models for the Study of Nagana. I. Parasitological, Hematological and Pathological Parameters. PLoS Negl. Trop. Dis. 2010, 4, e792. [Google Scholar] [CrossRef] [Green Version]
- Szöör, B.; Silvester, E.; Matthews, K.R. A Leap Into the Unknown—Early Events in African Trypanosome Transmission. Trends Parasitol. 2020, 36, 266–278. [Google Scholar] [CrossRef] [Green Version]
- Lun, Z.R.; Lai, D.H.; Li, F.J.; Lukes, J.; Ayala, F.J. Trypanosoma brucei: Two steps to spread out from Africa. Trends Parasitol. 2010, 26, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Carnes, J.; Anupama, A.; Balmer, O.; Jackson, A.; Lewis, M.; Brown, R.; Cestari, I.; Desquesnes, M.; Gendrin, C.; Hertz-Fowler, C.; et al. Genome and Phylogenetic Analyses of Trypanosoma evansi Reveal Extensive Similarity to T. brucei and Multiple Independent Origins for Dyskinetoplasty. PLoS Negl. Trop. Dis. 2015, 9, e3404. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.Z.; Lun, Z.R.; Zhu, X.Q.; Hide, G.; Lai, D.H. Further evidence from SSCP and ITS DNA sequencing support Trypanosoma evansi and Trypanosoma equiperdum as subspecies or even strains of Trypanosoma brucei. Infect. Genet. Evol. 2016, 41, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Desquesnes, M.; Holzmuller, P.; Lai, D.H.; Dargantes, A.; Lun, Z.R.; Jittaplapong, S. Trypanosoma evansi and surra: A review and perspectives on origin, history, distribution, taxonomy, morphology, hosts, and pathogenic effects. Biomed. Res. Int. 2013, 194176. [Google Scholar] [CrossRef] [Green Version]
- Molina, J.M.; Ruiz, A.; Juste, M.C.; Corbera, J.A.; Amador, R.; Gutiérrez, C. Seroprevalence of Trypanosoma evansi in dromedaries (Camelus dromedarius) from the Canary Islands (Spain) using an antibody Ab-ELISA. Prev. Vet. Med. 1999, 47, 53–59. [Google Scholar] [CrossRef]
- Gutiérrez, C.; Juste, M.C.; Corbera, J.A.; Magnus, E.; Verloo, D.; Montoya, J.A. Camel trypanosomosis in the Canary Island: Assessment of seroprevalence and infection rates using the card agglutination test (CATT/T. evansi) and parasite detection tests. Vet. Parasitol. 2000, 90, 155–159. [Google Scholar] [CrossRef]
- Tamarit, A.; Gutiérrez, C.; Arroyo, R.; Jimenes, V.; Zagalá, G.; Bosch, I.; Sirvent, J.; Alberola, J.; Alonso, I.; Caballero, C. Trypanosoma evansi infection in mainland Spain. Vet. Parasitol. 2010, 167, 74–76. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, C.; Desquesnes, M.; Touratier, L.; Büscher, P. Trypanosoma evansi: Recent outbreaks in Europe. Vet. Parasitol. 2010, 174, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, N.F.; Tejedor-Junco, M.T.; Hernández-Trujillo, Y.; González, M.; Gutiérrez, C. The role of wild rodents in the transmission of Trypanosoma evansi infection in an endemic area of the Canary Islands (Spain). Vet. Parasitol. 2010, 174, 323–327. [Google Scholar] [CrossRef]
- Desquesnes, M.; Bossard, G.; Patrel, D.; Herder, S.; Patout, O.; Lepetitcolin, E.; Thevenon, S.; Berthier, D.; Pavlovic, D.; Brugidou, R.; et al. First outbreak of Trypanosoma evansi in camels in metropolitan France. Vet. Record 2008, 162, 750–752. [Google Scholar] [CrossRef]
- Defontis, M.; Richartz, J.; Engelmann, N.; Bauer, C.; Schwierk, V.M.; Büscher, P.; Moritz, A. Canine Trypanosoma evansi infection introduced into Germany. Vet. Clin. Pathol. 2012, 41, 369–374. [Google Scholar] [CrossRef] [PubMed]
- EFSA AHAW Panel (EFSA Panel on Animal Health and Welfare); More, S.; Bøtner, A.; Butterworth, A.; Calistri, P.; Depner, K.; Edwards, S.; Garin-Bastuji, B.; Good, M.; Gortazar Schmidt, C.; et al. Scientific Opinion on the assessment of listing and categorization of animal diseases within the framework of the Animal Health Law (Regulation (EU) No 2016/429): Trypanosoma evansi infections (including Surra). EFSA J. 2017, 15, 4892. [Google Scholar] [CrossRef]
- Scott, D.W.; Miller, W.H. Viral and Protozoal Skin Disease. In Equine Dermatology, 2nd ed.; Saunders, Elsevier Science: St. Louis, MO, USA, 2003; pp. 376–394. [Google Scholar]
- Buffard, M.; Schneider, G. Le trypanosome de la dourine. Arc Parasitol. 1900, 3, 124–133. [Google Scholar]
- Claes, F.; Büscher, P.; Touratier, L.; Goddeeris, B.M. Trypanosoma equiperdum: Master of disguise or historical mistake? Trends Parasitol. 2005, 21, 316–321. [Google Scholar] [CrossRef]
- Pascucci, I.; Di Provvido, A.; Cammà, C.; di Francesco, G.; Calistri, P.; Tittarelli, M.; Ferri, N.; Scacchia, M.; Caporale, V. Diagnosis of dourine in outbreaks in Italy. Vet. Parasitol. 2012, 193, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Calistri, P.; Narcisi, V.; Atzeni, M.; de Massis, F.; Tittarelli, M.; Mercante, M.T.; Ruggieri, E.; Scacchia, M. Dourine Reemergence in Italy. J. Equine Vet. Sci. 2012, 33, 83–89. [Google Scholar] [CrossRef]
- Ahmed, Y.; Hagos, A.; Merga, B.; Van Soom, A.; Duchateau, L.; Goddeeris, B.M.; Govaere, J. Trypanosoma equiperdum in the horse—A neglected threat? Vlaams Diergeneeskundig Tijdschrift 2018, 87, 66–87. [Google Scholar] [CrossRef]
- Gizaw, Y.; Megersa, M.; Fayera, T. Dourine: A neglected disease of equids. Trop. Anim. Health Prod. 2017, 49, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.R.; Engstler, M.; Wolf, M. 18S rDNA Sequence-Structure Phylogeny of the Trypanosomatida (Kinetoplastea, Euglenozoa) with Special Reference on Trypanosoma. Res. Sq. 2020, 15. [Google Scholar] [CrossRef]
- Roberts, L.; Janovy, J., Jr. Foundations of Parasitology, 6th ed.; McGraw-Hill: New York, NY, USA, 2000; pp. 55–83. [Google Scholar]
- Kaufer, A.; Ellis, J.; Stark, D.; Barratt, J. The evolution of trypanosomatid taxonomy. Parasites Vectors 2017, 10, 287. [Google Scholar] [CrossRef]
- Votýpka, J.; Suková, E.; Kraeva, N.; Ishemgulova, A.; Duží, I.; Lukeš, J.; Yurchenko, V. Diversity of Trypanosomatids (Kinetoplastea: Trypanosomatidae) Parasitizing Fleas (Insecta: Siphonaptera) and Description of a New Genus Blechomonas gen.n. Protist 2013, 164, 763–781. [Google Scholar] [CrossRef]
- Deplazes, P.; Eckert, J.; Mathis, A.; von Samson-Himmelstierna, G.; Zahner, H. Phylum Euglenozoa. In Parasitology in Veterinary Medicine, 1st ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2016; pp. 59–79. [Google Scholar] [CrossRef]
- Gibson, W. Liaisons dangereuses: Sexual recombination among pathogenic trypanosomes. Res. Microbiol. 2015, 166, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Marquardt, W.C.; Demaree, R.S.; Grieve, R.B. Trypanosomes and Trypanosomiasis. In Parasitology & Vector Biology, 2nd ed.; Harcourt/Academic Press: San Diego, CA, USA, 2000; pp. 37–55. [Google Scholar]
- Stevens, J.R.; Noyes, H.A.; Schofield, C.J.; Gibson, W. The molecular evolution of Trypanosomatidae. Adv. Parasitol. 2001, 48, 1–56. [Google Scholar] [CrossRef]
- Gibson, W. Species concepts for trypanosomes: From morphological to molecular definitions? Kinetoplast. Biol. Dis. 2003, 2, 10. [Google Scholar] [CrossRef] [Green Version]
- Osório, A.L.A.R.; Madruga, C.R.; Desquesnes, M.; Soares, C.O.; Ribeiro, L.R.R.; Costa, S.C.G.D. Trypanosoma (Duttonella) vivax: Its biology, epidemiology, pathogenesis, and introduction in the New World—A review. Mem. Inst. Oswaldo Cruz 2008, 103, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Garcia, H.A.; Rodrigues, C.M.; Rodrigues, A.C.; Pereira, D.L.; Pereira, C.L.; Camargo, E.P.; Hamilton, P.B.; Teixeira, M.M. Remarkable richness of trypanosomes in tsetse flies (Glossina morsitans morsitans and Glossina pallidipes) from the Gorongosa National Park and Niassa National Reserve of Mozambique revealed by fluorescent fragment length barcoding (FFLB). Infect. Genet. Evol. 2018, 63, 370–379. [Google Scholar] [CrossRef]
- Hutchinson, R.; Gibson, W. Rediscovery of Trypanosoma (Pycnomonas) suis, a tsetse-transmitted trypanosome closely related to T. brucei. Infect. Genet. Evol. 2015, 36, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Auty, H.; Torr, S.J.; Michoel, T.; Jayaraman, S.; Morrison, L.J. Cattle trypanosomosis: The diversity of trypanosomes and implications for disease epidemiology and control. Rev. Sci. Tech. 2015, 34, 587–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echeverria, J.T.; Soares, R.L.; Crepaldi, B.A.; de Oliveira, G.G.; da Silva, P.M.P.; Pupin, R.C.; Martins, T.B.; Kellermann Cleveland, H.P.; Ramos, C.A.N.; Borges, F.A. Clinical and therapeutic aspects of an outbreak of canine trypanosomiasis. Rev. Bras. Parasitol. Vet. 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Mansfield, J.M. Nonpathogenic Trypanosomes of mammals. In Parasitic Protozoa, Taxonomy, Kinetoplastids and Flagellates of Fish; Kreier, J.P., Ed.; Academic Press, Inc.: New York, NY, USA, 1977; Volume 1, pp. 297–327. [Google Scholar]
- Sutherland, C.S.; Yukich, J.; Goeree, R.; Tediosi, F.A. Literature Review of Economic Evaluations for a Neglected Tropical Disease: Human African Trypanosomiasis (“Sleeping Sickness”). PLoS Negl. Trop. Dis. 2015, 9, e0003397. [Google Scholar] [CrossRef] [Green Version]
- De Fuentes-Vincente, J.A.; Gutiérrez-Cabrera, A.E.; Flores-Villegas, A.L.; Lowenberger, C.; Benelli, G.; Salazar-Schettino, P.M.; Córdoba-Aguilar, A. What makes an effective Chagas disease vector? Factors underlying Trypanosoma cruzi-triatomine interactions. Acta Trop. 2018, 183, 23–31. [Google Scholar] [CrossRef]
- Molyneux, D.H. The morphology and biology of Trypanosoma (Herpetosoma) evotomys of the bank-vole, Clethrionomys glareolus. Parasitology 1969, 59, 843. [Google Scholar] [CrossRef]
- Lun, Z.R.; Reid, S.A.; Lai, D.H.; Li, F.J. Atypical human trypanosomiasis: A neglected disease or just an unlucky accident? Trends Parasitol. 2009, 25, 107–108. [Google Scholar] [CrossRef]
- Desquesnes, M.; Yangtara, S.; Kunphukhieo, P.; Chalermwong, P.; Jittapalapong, S.; Herder, S. Zoonotic trypanosomes in South East Asia: Attempts to control Trypanosoma lewisi using veterinary drugs. Exp. Parasitol. 2016, 165, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; Mcveigh, R.; O’Neill, K.; Robbertse, B.; et al. NCBI Taxonomy: A comprehensive update on curation, resources and tools. Database 2020, baaa062. [Google Scholar] [CrossRef]
- Linhart, P.; Banďouchová, H.; Zukal, J.; Votypka, J.; Kokurewicz, T.; Dundarova, H.; Apoznanski, G.; Heger, T.; Kubickova, A.; Nemcova, M.; et al. Trypanosomes in eastern and central European bats. Acta Vet. Brno 2020, 89, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Bettencourt, A.; França, C. Sur un trypanosome de la chauve-souris. CR Soc. Biol. 1905, 57, 306. [Google Scholar]
- Gardner, R.A.; Molyneux, D.H. Schizotrypanum in British bats. Parasitology 1988, 97, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.R.; Miles, M.A. Trypanosoma (Schizotrypanum) dionisii breve n. subsp. from Chiroptera. Syst. Parasitol. 1979, 1, 61–65. [Google Scholar] [CrossRef]
- Mitkovska, V.; Chassovnikarova, T.; Dimitrov, H. First record of Trypanosoma infection in Mediterranean mouse (Mus macedonicus, Petrov & Ružić, 1983) in Bulgaria. ZooNotes 2014, 64, 1–6. [Google Scholar]
- Krampitz, H.E. Kritisches zur Taxonomie und Systematik parasitischer Säugetier-Trypanosomen mit besonderer Beachtung einiger der in Wülmäusen verbreiteten spezifischen Formen. Zschr. Tropenmed. 1961, 12, 117. [Google Scholar]
- Walter, G.; Liebisch, A. Studies of the ecology of some blood protozoa of wild small mammals in North Germany. Acta Trop. 1980, 37, 31–40. [Google Scholar]
- Elton, C. The health and parasites of a wild mouse population. Proc. Zool. Lond. 1931, 101, 657–721. [Google Scholar] [CrossRef]
- Turner, C.M.R.; Cox, F.E.G. Interspecific interactions between blood parasites in a wild rodent community. Ann. Trop. Med. PH 1985, 79, 463–465. [Google Scholar] [CrossRef] [PubMed]
- Cox, F.E.G. Protozoan parasites of British small rodents. Mammal. Rev. 1987, 17, 59–66. [Google Scholar] [CrossRef]
- Noyes, H.A.; Ambrose, P.; Barker, F.; Begon, M.; Bennet, M.; Bown, K.J.; Kemp, S.J. Host specificity of Trypanosoma (Herpetosoma) species: Evidence that bank voles (Clethrionomys glareolus) carry only one T.(H.) evotomys 18S rRNA genotype but wood mice (Apodemus sylvaticus) carry at least two polyphyletic parasites. Parasitology 2002, 124, 185–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karbowiak, G.; Sinski, E. The occurrence and morphological characteristics of a Trypanosma evotomys strain from North Poland. Acta Parasitol. 1996, 41, 105–107. [Google Scholar]
- Bajer, A.; Welc-Falęciakc, R.; Bednarska, M.; Alsarraf, M.; Behnke-Borowczyk, J.; Siński, E.; Behnke, J.M. Long-Term Spatiotemporal Stability and Dynamic Changes in the Haemoparasite Community of Bank Voles (Myodes glareolus) in NE Poland. Environ. Microbiol. 2014, 68, 196–211. [Google Scholar] [CrossRef] [Green Version]
- Karbowiak, G.; Stanko, M.; Fričová, J.; Wita, I.; Hapunik, J.; Peťko, B. Blood parasites of the striped field mouse Apodemus agrarius and their morphological characteristics. Biologia 2009, 64, 1219. [Google Scholar] [CrossRef]
- Laveran, A.; Pettit, A. Sur le trypanosome du mulot, Mus sylvaticus. CR Soc. Biol. 1909, 67, 564. [Google Scholar]
- Krampitz, H.E. Ueber das Europäische Waldmaustrypanosom, Trypanosoma grosi Laveran et Pettit 1909 (Promonadina, Trypanosomidae). Zschr. Parasitenk. 1959, 19, 232. [Google Scholar] [PubMed]
- Šebek, Z. Blood Parasites of Small Mammals in Western Hungary. Parasitol. Hung. 1978, 11, 17–22. [Google Scholar]
- Alharbi, B. Arthropod-Borne Infections in the United Kingdom and Saudi Arabia. Ph.D. Thesis, University of Salford, Salford, UK, 2018. [Google Scholar]
- Gros, G. Observations et inductions microscopiques sur quelques parasites. Bull. Soc. Imp. Nat. Mosc. 1845, 18, 380. [Google Scholar]
- Yakimoff, W.L.; Korssak, D.W. Hämatoparasitologische Notizen. II. Ein Trypanosoma der Feldmaus im ostasiatischen Ufergebiete. Cbl. Bakt. 1910, 55, 370. [Google Scholar]
- Karbowiak, G.; Wita, I. Trypanosoma (Herpetosoma) grosi kosewiense subsp.n., the Parasite of the Yellow-Necked Mouse Apodemus flavicollis (Melchior, 1834). Acta Protozool. 2004, 43, 173–178. [Google Scholar] [CrossRef]
- Rislakki, V. Studies on the Prevalence and Effect of Trypasnosoma lewisi Infection in Finnish Rats. Acta Vet. Scand. 1971, 12, 448–450. [Google Scholar] [CrossRef] [PubMed]
- Wiger, R. Seasonal and annual variations in the prevalence of blood parasites in cyclic species of small rodents in Norway with special reference to Clethrionomys glareolus. Ecography 1979, 2. [Google Scholar] [CrossRef]
- Karbowiak, G.; Wita, I.; Czaplińska, U. The occurrence and ultrastructure of Trypanosoma (Herpetosoma) lewisi (Kent, 1880) Laveran and Mesnil, 1901, the parasite of rats (Rattus norvegicus) in Poland. Wiad. Parazytol. 2009, 55, 249–258. [Google Scholar]
- Martincović, F.; Matanović, K.; Rodrigues, A.C.; Garcia, H.A.; Teixeira, M.M.G. Trypanosoma (Megatrypanum) melophagium in the sheep ked Melophagus ovinus from organic farms in Croatia: Phylogenetic interferences support restriction to sheep and sheep keds and close relationship with trypanosomes from other ruminant species. J. Eukaryot. Microbiol. 2012, 59, 134–144. [Google Scholar] [CrossRef]
- Pfeiffer, E. Ueber trypanosomenahnliche Plagellaten im Darm von Melophagus ovinus. Zschr. Hyg. Infektkr. 1905, 30, 324–330. [Google Scholar] [CrossRef] [Green Version]
- Behn, P. Weitere Trypasomenbefunde beim Schafe. Berl. Tierärztl. Wschr. 1912, 50, 934. [Google Scholar]
- Büscher, G.; Friedhoff, K.T. The Morphology of Ovine Trypanosoma melophagium (Zoomastigophorea: Kinetoplastida) 1. J. Protozool. 1984, 31, 98–101. [Google Scholar] [CrossRef]
- Douwes, J.A. Trypanosomen bij het Schaap (Trypanosoma Melophagium, Flu). Tijdschr. Diegeneesk 1920, 47, 408. [Google Scholar]
- Bozhenko, V.P.; Zeiss, A.L. Trypanosomiasis of sheep. Rev. Microbiol. 1928, 7, 417–420. [Google Scholar]
- Woodcock, H.M. A reply to Miss Porter’s note entitled “Some remarks on the genera Crithidia, Herpetomonas and Trypanosoma”. Parasitology 1911, 4, 150. [Google Scholar] [CrossRef]
- Hoare, C.A. An experimental study of the sheep-trypanosome (T. melophagium Flu, 1908) and its transmission by the sheep-ked (Melophagus ovinus L.). Parasitology 1923, 15, 365. [Google Scholar] [CrossRef]
- Small, R.W. A review of Melophagus ovinus (L.), the sheep ked. Vet. Parasitol. 2005, 130, 141–155. [Google Scholar] [CrossRef]
- Craig, B.H.; Tempest, L.J.; Pilkington, J.G.; Pemberton, J.M. Metazoan-protozoan parasite co-infections and host body weight in St Kilda Soay sheep. Parasitology 2008, 135, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Gibson, W.; Pilkington, J.G.; Pemberton, J.M. Trypanosoma melophagium from the sheep ked Melophagus ovinus on the island of St. Kilda. Parasitology 2010, 137, 1799–1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katic, R.V. Researches on Trypanosoma melophagium and its Incidence in Yugoslavia. Jug. Vet. Glasnik 1940, 20, 373–384. [Google Scholar]
- Jolyet, F.; Nabias, B. Sur un hématozoarie du lapin domestique. J. Méd. Bordx. 1891, 20, 325. [Google Scholar]
- Díaz-Sáez, V.; Merino-Espinosa, G.; Morales-Yuste, M.; Corpas-López, V.; Pratlong, F.; Morillas-Márquez, F.; Martín-Sánchez, J. High rates of Leishmania infantum and Trypanosoma nabiasi infection in wild rabbits (Oryctolagus cuniculus) in sympatric and syntrophic conditions in an endemic canine leishmaniasis area: Epidemiological consequences. Vet. Parasitol. 2014, 202, 119–127. [Google Scholar] [CrossRef]
- Merino-Espinosa, G.; Corpas-López, V.; Morillas-Márquez, F.; Díaz-Sáez, V.; Martín-Sánchez, J. Genetic variability and infective ability of the rabbit trypanosome, Trypanosoma nabiasi Railliet 1895, in southern Spain. Infect. Genet. Evol. 2016, 45, 98–104. [Google Scholar] [CrossRef]
- Grewal, M.S. The life-cycle of the British rabbit trypanosome, Trypanosoma nabiasi Railliet, 1985. Parasitology 1957, 47, 100. [Google Scholar] [CrossRef] [PubMed]
- Rioux, J.-A.; Albaret, J.-L.; Bres, A.; Dumas, A. Présence de Trypanosoma pestanai Bettencourt et França, 1905, chez les Blaireaux du sud de la France. Ann. Parasitol. Hum. Comp. 1966, 41, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Dirie, M.F.; Bornstein, S.; Wallbanks, K.R.; Stiles, J.K.; Molyneux, D.H. Zymogram and life-history studies on trypanosomes of the subgenus Megatrypanum. Parasitol. Res. 1990, 76, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Dyachenko, V.; Steinmann, M.; Bangoura, B.; Selzer, M.; Munderloh, U.; Daugschies, A. Co-infection of Trypanosoma pestanai and Anaplasma phagocytophilum in a dog from Germany. Vet. Parasitol. Reg. 2017, 9, 110–114. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, G.; Shiel, R.; O’Rourke, L.; Murphy, D.; Corner, L.; Costello, E.; Gormley, E. Bronchoalveolar lavage cytology from captive badgers. Vet. Clin. Pathol. 2009, 38, 381–387. [Google Scholar] [CrossRef]
- Peirce, M.A.; Neal, C. Trypanosoma (Megatrypanum) pestanai in British badgers (Meles meles). Int. J. Parasitol. 1974, 4, 439–440. [Google Scholar] [CrossRef]
- Macdonald, D.W.; Anwar, M.; Newman, C.; Woodroffe, R.; Johnson, P.J. Inter-annual differences in the age-related prevalences of Babesia and Trypanosoma parasites of European badgers (Meles meles). J. Zool. 1999, 247, 65–70. [Google Scholar] [CrossRef]
- Lizundia, R.; Newman, C.; Buesching, C.D.; Ngugi, D.; Blake, D.; Wa Sin, Y.; Macdonald, D.W.; Wilson, A.; McKeever, D. Evidence for a role of the Host-Specific Flea (Paraceras melis) in the Transmission of Trypanosoma (Megatrypanum) pestanai to the European Badger. PLoS ONE 2011, 6, e16977. [Google Scholar] [CrossRef]
- Sin, Y.W.; Annavi, G.; Dugdale, H.L.; Newman, C.; Burke, T.; Macdonald, D.W. Pathogen burden, co-infection and major histocompatibility complex variability in the European badger (Meles meles). Mol. Ecol. 2014, 23, 5072–5088. [Google Scholar] [CrossRef] [PubMed]
- Ideozu, E.J.; Whiteoak, A.M.; Tomlinson, A.J.; Robertson, A.; Delahay, R.J.; Hide, G. High prevalence of trypanosomes in European badgers detected using ITS-PCR. Parasites Vectors 2015, 8, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kingston, N.; Bobek, B.; Perzanowski, K.; Wita, I.; Maki, L. Description of Trypanosoma (Megatrypanum) stefanskii sp. n. from Roe Deer (Capreolus capreolus) in Poland. Proc. Helminthol. Soc. Wash. 1992, 59, 89–95. [Google Scholar]
- Verloo, D.; Brandt, J.; Van Meirvenne, N.; Büscher, P. Comparative in vitro isolation of Trypanosoma theileri from cattle in Belgium. Vet. Parasitol. 2000, 89, 129–132. [Google Scholar] [CrossRef]
- Bittner, L.; Wöckell, A.; Snedec, T.; Delling, C.; Böttcher, D.; Klose, K.; Köller, G.; Starke, A. Case report: Trace Mineral Deficiency with concurrent Detection of Trypanosoma theileri in a Suckler Cow Herd in Germany. Anim Biol. 2019, 21, 84. [Google Scholar]
- Frank, G. Ueber den Befund von Trypanosomen bei einem in Stein-Wingert (Westerwald, Regierungsbezirk Wiesbaden) verendeten Rinde. Zschr. Infektionskrankh Haustiere 1909, 5, 313. [Google Scholar]
- Böse, R.; Friedhoff, K.T.; Olbrich, S.; Büscher, G.; Domeyer, I. Transmission of Trypanosoma theileri to cattle by Tabanidae. Parasitol. Res. 1987, 73, 421–424. [Google Scholar] [CrossRef]
- Böse, R.; Petersen, K.; Pospichal, H.; Buchanan, N.; Tait, A. Characterization of Megatrypanum trypanosomes from European Cervidae. Parasitology 1993, 107, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amato, B.; Mira, F.; Lo Presti, V.D.M.; Guercio, A.; Russotto, L.; Gucciardi, F.; Vitale, M.; Lena, A.; Loria, G.R.; Puleio, R.; et al. A case of bovine trypanosomiasis caused by Trypanosoma theileri in Sicily, Italy. Parasitol. Res. 2019, 118, 2723–2727. [Google Scholar] [CrossRef]
- Greco, A.; Loria, G.R.; Dara, S.; Luckins, T.; Sparagano, O. First Isolation of Trypanosoma theileri in Sicilian cattle. Vet. Res. Commun. 2000, 24, 471–475. [Google Scholar] [CrossRef]
- Doherty, M.L.; Windle, H.; Vooheis, H.P.; Larkin, H.; Casey, M.; Clery, D.; Murray, M. Clinical disease associated with Trypanosoma theileri infection in a calf in Ireland. Vet. Record 1993, 132, 653–656. [Google Scholar] [CrossRef]
- Werszko, J.; Szewczyk, T.; Steiner-Bogdaszewska, Ż.; Wróblewski, P.; Karbowiak, G.; Laskowski, Z. Molecular detection of Megatrypanum trypanosomes in tabanid flies. Med. Vet. Entomol. 2019, 5p. [Google Scholar] [CrossRef]
- Karbowiak, G.; Wita, I.; Czaplińska, U. Trypanosoma (Megatrypanum) Wrublewskii, Wladimiroff et Yakimoff 1909, the parasite of European bison (Bison bonasus L). In Proceedings of the Trudy 5 Respublikanskoj Nauchno-Prakticheskoj Konferentsii, Vitebsk, Belarus, 21–22 September 2006; pp. 268–270. (In Russian). [Google Scholar]
- Karbowiak, G.; Wita, I.; Czaplińska, U. Pleomorfizm of trypanosomes occurring in Bison bonasus L. Wiad Parazytol. 2007, 53, 54. [Google Scholar]
- Karbowiak, G.; Demiaszkiewicz, A.W.; Pyziel, A.M.; Wita, I.; Moskowa, B.; Werszko, J.; Bień, J.; Goździk, K.; Lachowicz, J.; Cabaj, W. The parasitic fauna of the European bison (Bison bonasus) (Linnaeus, 1758) and their impact on the conservation. Part 1: The summarising list of parasites noted. Acta Parasitol. 2014, 53, 363–371. [Google Scholar] [CrossRef]
- Wladimiroff, A.; Yakimoff, W. Bemerkung zur vorstehenden Mitteilung Wrublewskis. Zentralbl Bakteriol. Orig A 1909, 48, 164. [Google Scholar]
- Espinosa, A.V.; Gutiérrez, C.; Gracia, E.; Moreno, B.; Chacón, G.; Sanz, P.V.; Büscher, P.; Touratier, L. Presencia de Trypanosoma theileri en bovinos en España. Albéitar Publ. Vet. Indep. 2006, 93, 36–37. [Google Scholar]
- Villa, A.; Gutierrez, C.; Garcia, E.; Moreno, B.; Chaćon, G.; Sanz, P.V.; Büscher, P.; Touratier, L. Presence of Trypanosoma theileri in Spanish Cattle. Ann. N. Y. Acad. Sci. 2008, 1149, 352–354. [Google Scholar] [CrossRef] [PubMed]
- Chatton, E.; Courrier, R. Un Schizotrypanum chez les chauve-souris (Vesperugo pipistrellus) en Basse-Alsace. Schizotrypanose et goître endémique. CR Soc. Biol. 1921, 84, 943. [Google Scholar]
- Dionisi, A. La malaria in alcune specie di pipistrelli. Atti Soc. Studi Malaria 1899, 1, 133. [Google Scholar]
- Battaglia, M. Alcune ricerche su due tripanosomi (Trypanosoma vespertilionis-Trypanosoma lewisi). Ann. Med. Navale 1904, 10, 517. [Google Scholar]
- Gonder, R. Trypanosoma vespertilionis (Battaglia). Centralbl. f Bakteriol. 1910, 53, 293–302. [Google Scholar]
- Mettam, A.E. On the presence of a trypanosome in an Irish bat. Dublin J. Med. Sci. 1907, 124, 417–419. [Google Scholar] [CrossRef]
- Petrie, G.F. Observations relating to the structure and geographical distribution of certain trypanosomes. J. Hyg. 1905, 5, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Coles, A.C. Blood parasites found in mammals, birds and fishes in England. Parasitology 1914, 7, 17–61. [Google Scholar] [CrossRef]
- Molyneux, D.H.; Bafort, J.M. Observations on the trypanosome of Pipistrellus pipistrellus in Britain, Trypanosoma (Schizotrypanum) vespertilionis. Ann. Soc. Belg. Med. Trop. 1971, 51, 335–348. [Google Scholar]
- Calzolari, M.; Rugna, G.; Clementi, E.; Carra, E.; Pinna, M.; Bergamini, F.; Fabbi, M.; Dottori, M.; Sacchi, L.; Votýpka, J. Isolation of a Trypanosome Related to Trypanosoma theileri (Kinetoplastea: Trypanosomadidae) from Phlebotomus perfiliewi (Diptera: Pychodidae). BioMed Res. Int. 2018, 8p. [Google Scholar] [CrossRef] [Green Version]
- Werszko, J.; Steiner-Bogdaszewska, Ż.; Jeżewski, W.; Szewczyk, T.; Kuryło, G.; Wołkowycki, M.; Wróblewski, P.; Karbowiak, G. Molecular detection of Trypanosoma spp. in Lipoptena cervi and Lipoptena fortisetosa (Diptera: Hippoboscidae) and their potential role in the transmission of pathogens. Parasitology 2020, 147, 1629–1635. [Google Scholar] [CrossRef]
- Neumüller, M.; Nilsson, K.; Påhlson, C. Trypanosoma spp. In Swedish game animals. Parasitol. Res. 2012, 110, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Ganyukova, A.I.; Zolotarev, A.V.; Malysheva, M.N.; Frolov, A.O. First record of Trypanosoma theileri-like flagellates in horseflies from Northwest Russia. Protistology 2018, 12, 223–230. [Google Scholar] [CrossRef]
- Friedhoff, K.T.; Petrich, J.; Hoffmann, M.; Büscher, G. Trypanosomes in Cervidae in Germany. Zentralbl. Bakteriol. Mikrobiol. Hyg. A 1984, 256, 286–287. [Google Scholar] [CrossRef]
- Hoffmann, M.; Büscher, G.; Friedhoff, K.T. Stercorarian Trypanosomes from Deer (Cervidae) in Germany. J. Protozool. 1984, 31, 581–584. [Google Scholar] [CrossRef]
- Szentiványi, T.; Markotter, W.; Dietrich, M.; Clément, L.; Ançay, L.; Brun, L.; Genzoni, E.; Kearney, T.; Seamark, E.; Estók, P.; et al. Host conservation through their parasites: Molecular surveillance of vector-borne microorganisms in bats using ectoparasitic bat flies. Parasite 2020, 27, 72. [Google Scholar] [CrossRef] [PubMed]
- Kingston, N.; Drozdz, J.; Rutkowska, M. Trypanosoma sp. in red deer (Cervus elaphus) and elk (Alces alces) in Poland. Proc. Helrninthol. Soc. Wash. 1985, 52, 144–145. [Google Scholar]
- Kingston, N.; Bobek, B. A trypanosome in roe deer, Capreolus capreolus, in southern Poland. Proc. Helminthol. Soc. Wash. 1985, 52, 143. [Google Scholar]
- Olmeda, A.S.; San Miguel, J.M.; Luzón, M. Primera denuncia de Trypanosoma (Megatrypanum) en ciervos (Cervus elaphus) de la Península Ibérica. Galemys 2001, 13, 149–153. [Google Scholar]
- Reglero, M.; Vicente, J.; Rouco, C.; Villafuerte, R.; Gortazar, C. Trypanosoma spp. infection in wild rabbits (Oryctolagus cuniculus) during a restocking program in southern Spain. Vet. Parasitol. 2007, 149, 178–184. [Google Scholar] [CrossRef]
- Bray, D.P.; Bown, K.J.; Stockley, P.; Hurst, J.L.; Bennett, M.; Birtles, R.J. Haemoparasites of common shrews (Sorex araneus) in northwest England. Parasitology 2007, 134, 819–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoyte, H.M. The morphology of Trypanosoma theileri in the blood of cattle, and the rediscovery of Theileria mutans in England. Z. Parasitenkd. 1972, 38, 183–199. [Google Scholar] [CrossRef]
- Wells, E.A. Isolation of Trypanosoma theileri Laveran 1902, from Cattle in Scotland. Nature 1965, 4986, 847. [Google Scholar] [CrossRef] [PubMed]
- Laveran, A. Sur un nouveau trypanosome des bovidés (Trypanosoma theileri). CR Acad. Sci. 1902, 134, 512–514. [Google Scholar]
- Theiler, A. A new Trypanosoma, and the disease caused by it. J. Comp. Pathol. Ther. 1903, 16, 193–209. [Google Scholar] [CrossRef]
- Herbert, I.V. Trypanosoma theileri Laveran, 1902. A cosmopolitan parasite of cattle. Vet. Bull. 1964, 34, 563. [Google Scholar]
- Turner, A.W.; Murname, D. Trypanosomes in the Blood of Victorian Animals. I. A Preliminary Note on the Occurrence of Trypanosoma theileri in the Blood of Cattle. J. Sci. Ind. Res. 1930, 3, 120–121. [Google Scholar]
- Woo, P.; Soltys, M.A.; Gillick, A.C. Trypanosomes in Cattle in Southern Ontario. Can. J. Vet. Res. 1970, 34, 142–147. [Google Scholar]
- Matthews, D.M.; Kingston, N.; Maki, L.; Nelms, G. Trypanosoma theileri Laveran, 1902, in Wyoming cattle. Am. J. Vet. Res. 1979, 40, 623–629. [Google Scholar] [PubMed]
- Kingston, N.; Nikander, S. Poron, Rangifer tarandus, Trypanosoma sp. Suomessa (Trypanosoma sp. in reindeer (Rangifer tarandus) in Finland). Suomen Elainlddkarilehti 1985, 91, 1. [Google Scholar]
- Garcia, H.A.; Rodrigues, A.C.; Martinkovic, F.; Minervino, A.H.; Campaner, M.; Nunes, V.L.B.; Paiva, F.; Hamilton, P.B.; Teixeira, M.M.G. Multilocus phylogeographical analysis of Trypanosoma (Megatrypanum) genotypes from sympatric cattle and water buffalo populations supports evolutionary host constraint and close phylogenetic relationships with genotypes found in other ruminants. Int. J. Parasitol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Kocan, A.A. Blood-Inhabiting Protozoan Parasites. In Parasitic Diseases of Wild Mammals, 2nd ed.; Samuel, W.M., Margo, J.P., Kocan, A.A., Eds.; Iowa State University Press: Iowa City, IA, USA, 2001; pp. 520–522. [Google Scholar]
- Galuppi, R.; Bonoli, C.; Aureli, S.; Cassini, R.; Marcer, F.; Foley, J.E.; Tampieri, M.P. Comparison of diagnostic methods to detect piroplasms in asymptomatic cattle. Vet. Parasitol. 2012, 183, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.W.; Murnane, D. On the presence of the non-pathogenetic Trypanosoma melophagium in the blood of Victorian sheep, and its transmission by Melophagus ovinus. Aust. J. Exp. Biol. Med. Sci. 1930, 7, 5–8. [Google Scholar] [CrossRef]
- Flu, P.C. Ueber die Flagellaten im Darm von Melophagium ovinus. Protist 1908, 12, 147. [Google Scholar]
- Nöller, W. Beitrag zur Kenntnis des Schaftrypanosomas. Arch. Schiffs Tropenhyg. 1919, 23, 99. [Google Scholar]
- Kleine, F.K. Beitrang zum Kenntnis des Trypanosoma melophagium, Flu. Deutsch Tierärztl. 1919, 27, 408. [Google Scholar]
- Nalbantoglu, S.; Karaer, Z. Trypanosoma melophagium in blood cell culture. Ankara Univ. Vet. Fak. Derg. 2008, 55, 173–176. [Google Scholar]
- Chaussat, J.B. Recerches Microscopiques Appliquées à la Pathologiée des Hématozoàires. Ph.D. Thesis, Faculté de Médicine de Paris, Paris, France, 1850. [Google Scholar]
- Lewis, T.R. Flagellated organisms in the blood of healthy rats. Q. J. Microsc. Sci. 1879, 19, 109. [Google Scholar]
- Minchin, E.A.; Thompson, J.D. The Rat-Trypanosoma lewisi, in its relation to the Rat-Flea, Ceratophyllus fasciatus. J. Cell Sci. 1915, 2, 463–681. [Google Scholar]
- Zhang, X.; Li, S.J.; Li, Z.; He, C.Y.; Hide, G.; Lai, D.H.; Lun, Z.R. Cell cycle and cleavage events during in vitro cultivation of bloodstream forms of Trypanosoma lewisi, a zoonotic pathogen. Cell Cycle 2019, 18, 552–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truc, P.; Büscher, P.; Cuny, G.; Gonzatti, M.I.; Jannin, J.; Joshi, P.; Juyal, P.; Lun, Z.R.; Mattioli, R.; Pays, E.; et al. Atypical human infections by animal trypanosomes. PLoS Negl. Trop. Dis. 2013, 7, e2256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lun, Z.R.; Wen, Y.Z.; Uzureau, P.; Lecordier, L.; Lai, D.H.; Lan, Y.G.; Desquesnes, M.; Geng, G.Q.; Yang, T.B.; Zhou, W.L.; et al. Resistance to normal human serum reveals Trypanosoma lewisi as an underestimated human pathogen. Mol. Biochem. Parasitol. 2015, 199, 58–61. [Google Scholar] [CrossRef]
- Brumpt, E. Evolution de Trypanosoma Lewisi, Duttoni, Nabiasi, Blanchardi, chez les puces et les punaises. Transmission par les dejections. Comparaison avec T. cruzi. Bull. Soc. Pathol. Exot. 1913, 6, 167–171. [Google Scholar]
- Channon, H.A.; Wright, H.D. Observations on Trypanosomiasis of Rabbits and its natural Mode of Transmission. J. Pathol. Bacteriol. 1927, 30, 253–260. [Google Scholar] [CrossRef]
- Watson, E.A.; Hadwen, S. Trypanosomoses found in Canadian mammals. Parasitology 1912, 5, 21. [Google Scholar] [CrossRef]
- Thiroux, A. Rescherches morphologiques et expérimentales sur Trypanosoma duttoni (Thiroux) Ann. Inst. Pasteur. 1905, 19, 564. [Google Scholar]
- Monroy, F.P.; Dusanic, D.G. The Kidney Form of Trypanosoma musculi: A Distinct Stage in the Life Cycle? Parasitol. Today 2000, 6, 107–110. [Google Scholar] [CrossRef]
- Zhang, X.; Hong, X.K.; Li, S.J.; Lai, D.H.; Hide, G.; Lun, Z.R.; Wen, Y.Z. The effect of normal human serum on the mouse trypanosome Trypanosoma musculi in vitro and in vivo. Exp. Parasitol. 2018, 184, 115–120. [Google Scholar] [CrossRef]
- Austen, J.; Van Kampen, E.; Egan, S.; O’Dea, M.; Jackson, B.; Ryan, U.; Irwin, P.J.; Prada, D. First report of Trypanosoma dionisii (Trypanosomatidae) identified in Australia. Parasitology 2020, 147, 1801–1809. [Google Scholar] [CrossRef] [PubMed]
- Soria, C.A.; Dusanic, D.G. Comparative studies of Trypanosoma vespertilionis Battaglia and Trypanosoma dionisii Bettencourt & França. J. Protozool. 1975, 22, 509–513. [Google Scholar] [CrossRef] [PubMed]
- Bettencourt, A.; França, C. Sur un trypanosome du blairau (Meles taxus, Schreib). CR Soc. Biol. 1905, 57, 306. [Google Scholar]


{kind=link}
{kind=link}
| Group | Subgenus/Clade | Species | Subspecies |
|---|---|---|---|
| Salivaria | Duttonella | T. vivax | |
| Nannomonas | T. congolese | ||
| T. godfreyi | |||
| T. simiae | |||
| Pycnomonas | T. suis | ||
| Trypanozoon | T. brucei | T. brucei brucei | |
| T. brucei gambiense | |||
| T. brucei rhodensiense | |||
| T. brucei equiperdum | |||
| T. brucei evansi | |||
| Stercoraria | Herpetosoma | T. lewisi | |
| T. rangeli | |||
| Megatrypanum | T. melophagium | ||
| Schizotrypanum | T. cruzi | ||
| T. dionisii | T. dionisii breve | ||
| T. vespertilionis | |||
| Trypanosomes with unspecified subgenus | T. evotomys | ||
| T. grayi | |||
| T. grosi | T. grosi kosewiense | ||
| T. pestenai | |||
| T. theileri | |||
| T. nabiasi | |||
| Unclassified trypanosomes | Fish trypanosomes | ||
| Other unclassified trypanosomes |
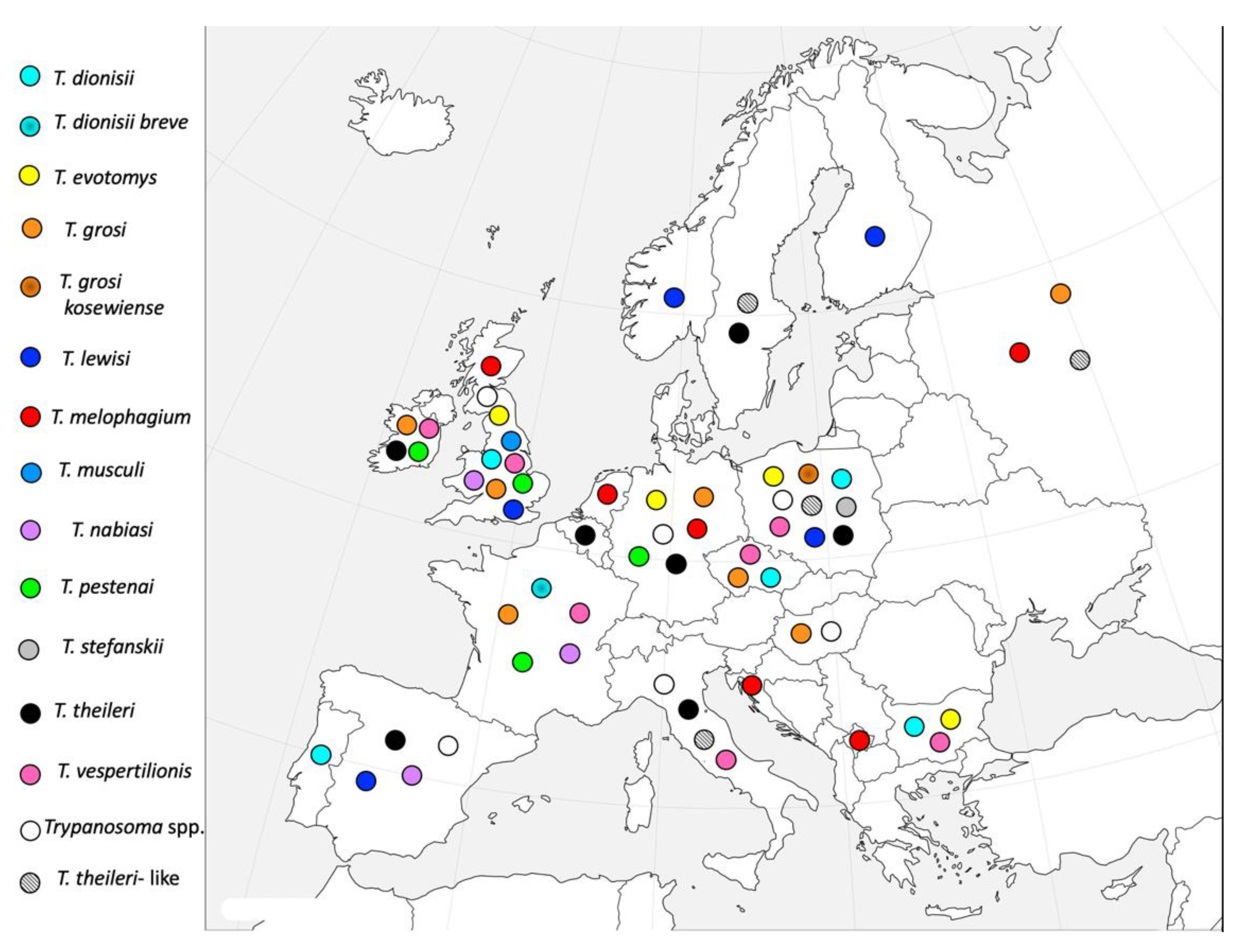
| Trypanosoma sp. | Country | Host | References |
|---|---|---|---|
| T. dionisii | Bulgaria | Bat and bat flies (Nycteribia shmidlii) | [73] |
| Czech Republic | Bat and bat flies | [73] | |
| Poland | Bat and bat flies | [73] | |
| Portugal | Bat | [74] | |
| United Kingdom | Bat and bat flies | [75] | |
| T. dionisii breve | France | Bat | [76] |
| T. evotomys | Bulgaria | Mus macedonicus | [77] |
| Germany | Voles | [78,79] | |
| United Kingdom | Voles | [69,80,81,82,83] | |
| Poland | Voles | [84,85] | |
| T. grosi | Czech Republic | Apodemus agrarius | [86] |
| France | Apodemus sylvaticus | [87] | |
| Germany | Voles | [88] | |
| Hungary | Voles | [89] | |
| Ireland | Small rodents | [90] | |
| United Kingdom | Small rodents | [80,82,90] | |
| Russia | Apodemus sylvaticus | [91,92] | |
| T. grosi kosewiense | Poland | Voles | [93] |
| T. lewisi | Finland | Rattus norvegicus, R. rattus | [94] |
| Norway | Clethrionomys glareolus, Microtus agrestis; Apodemus sylvaticus | [95] | |
| Poland | Rattus norvegicus | [96] | |
| United Kingdom | Rattus norvegicus | [82] | |
| Spain | Rattus norvegicus, R. rattus | [41] | |
| T. melophagium | Croatia | Sheep ked (Mallophagus melophagium) | [97] |
| Germany | Sheep ked | [98] | |
| Sheep | [99,100] | ||
| Holland | Sheep | [101] | |
| Russia | Sheep | [102] | |
| United Kingdom | Sheep | [103,104,105,106] | |
| Sheep ked | [107] | ||
| Former Yugoslavia (Croatia and Kosovo) | Sheep | [108] | |
| T. musculi | United Kingdom | Mouse (Mus musculus) | [82] |
| T. nabiasi | France | Wild and domestic rabbits | [109] |
| Spain | Rabbits (Oryctolagus cuniculus) | [110,111] | |
| United Kingdom | Rabbits | [112] | |
| T. pestenai | France | European badger (Meles meles) | [113,114] |
| Germany | Dog | [115] | |
| Ireland | Badger | [116] | |
| United Kingdom | Badger | [117,118,119,120,121] | |
| T. stefanskii | Poland | Roe deer (Capreolus capreolus) | [122] |
| T. theileri | Belgium | Cattle | [123] |
| Germany | Cattle | [124,125] | |
| Fallow deer (Dama dama), red deer (Cervus elaphus), roe deer | [126,127] | ||
| Italy | Cattle | [128] | |
| River buffalo (Bubalus bubalis) | [129] | ||
| Ireland | Calf | [130] | |
| Poland | Cattle | [131] | |
| European bison (Bison bonasus) | [132,133,134,135] | ||
| Sweden | Cattle | [122] | |
| Spain | Cattle | [136,137] | |
| T. vespertilionis | Bulgaria | Bat and bat flies | [73] |
| Czech Republic | Bat and bat flies | [73] | |
| France | Bat | [138] | |
| Italy | Bat | [139,140,141] | |
| Ireland | Bat | [142] | |
| Poland | Bat and bat flies | [73] | |
| United Kingdom | Bat | [143,144,145] | |
| Bat and bat flies | [75] | ||
| T. theileri-like | Italy | Sandflies (Phlebotomus perfiliewi) | [146] |
| Poland | Deer ked (Lipoptena fortisetosa and L. cervi) | [147] | |
| Sweden | Roe deer, fallow deer, European elk, red deer, wild boar (Sus scrofa) | [148] | |
| Russia | Horseflies (Hybomitra tarandina, H. muehlfeldi, H. bimaculate, Chrysops divaricatus) | [149] | |
| Trypanosoma sp. | Germany | Fallow deer, red deer, roe deer | [150,151] |
| Italy | Bat and Cimex spp. | [152] | |
| Hungary | Bat and Cimex spp. | [152] | |
| Poland | Roe deer, red deer, European elk (Alces alces) | [153] | |
| Roe deer | [154] | ||
| Spain | Bat and Cimex spp. | [152] | |
| Red deer | [155] | ||
| Wild rabbits | [156] | ||
| United Kingdom | Common shrews (Sorex araneus) | [157] | |
| Cattle | [114,158,159] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magri, A.; Galuppi, R.; Fioravanti, M. Autochthonous Trypanosoma spp. in European Mammals: A Brief Journey amongst the Neglected Trypanosomes. Pathogens 2021, 10, 334. https://doi.org/10.3390/pathogens10030334
Magri A, Galuppi R, Fioravanti M. Autochthonous Trypanosoma spp. in European Mammals: A Brief Journey amongst the Neglected Trypanosomes. Pathogens. 2021; 10(3):334. https://doi.org/10.3390/pathogens10030334
Chicago/Turabian StyleMagri, Alice, Roberta Galuppi, and Marialetizia Fioravanti. 2021. "Autochthonous Trypanosoma spp. in European Mammals: A Brief Journey amongst the Neglected Trypanosomes" Pathogens 10, no. 3: 334. https://doi.org/10.3390/pathogens10030334