
Akt Interacts with Usutu Virus Polymerase, and Its Activity Modulates Viral Replication

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
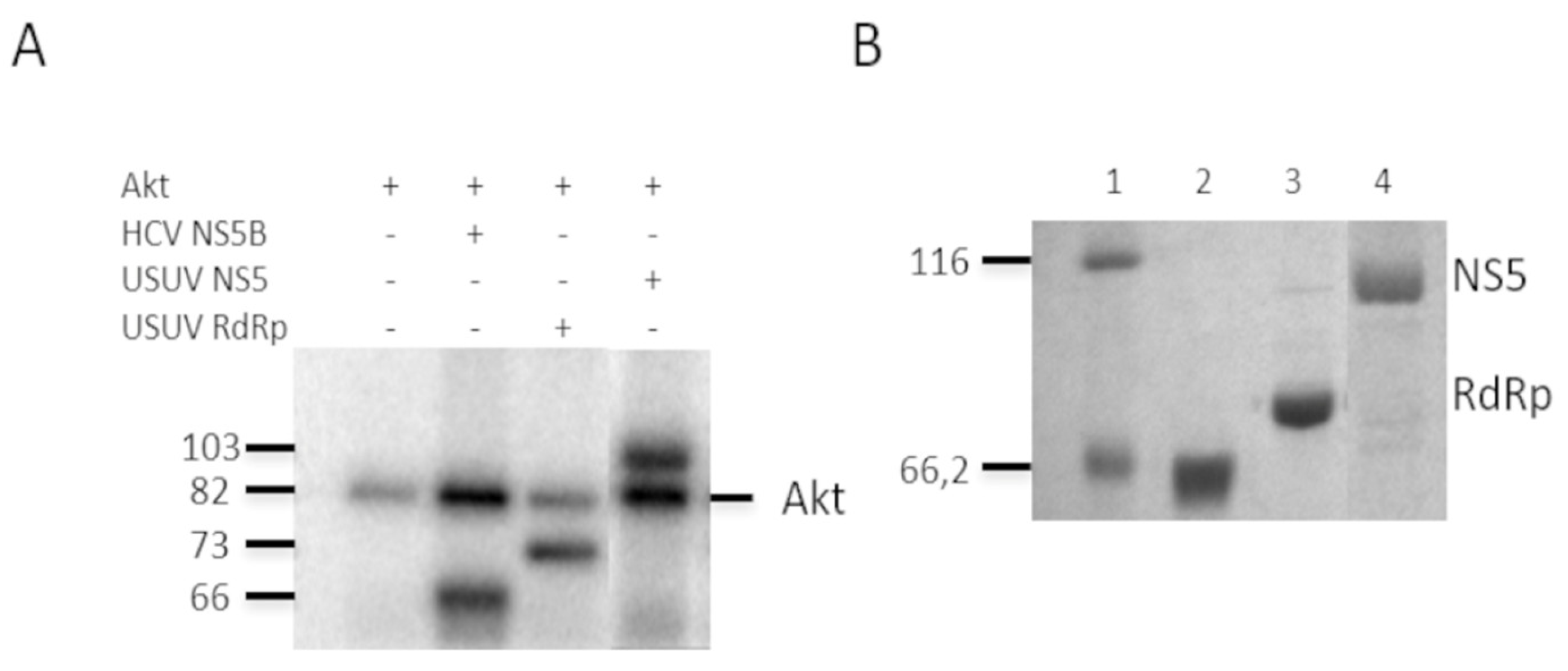
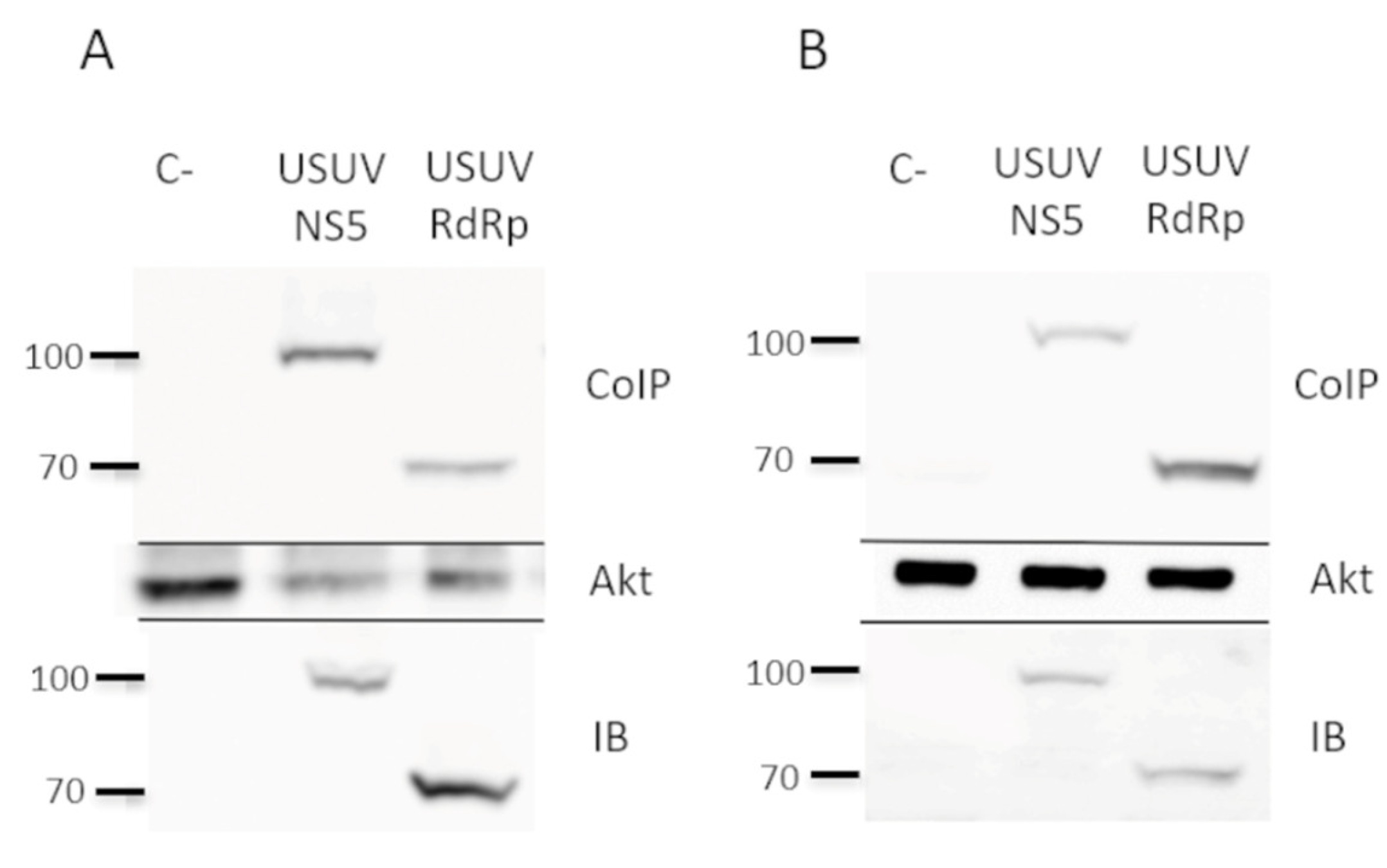
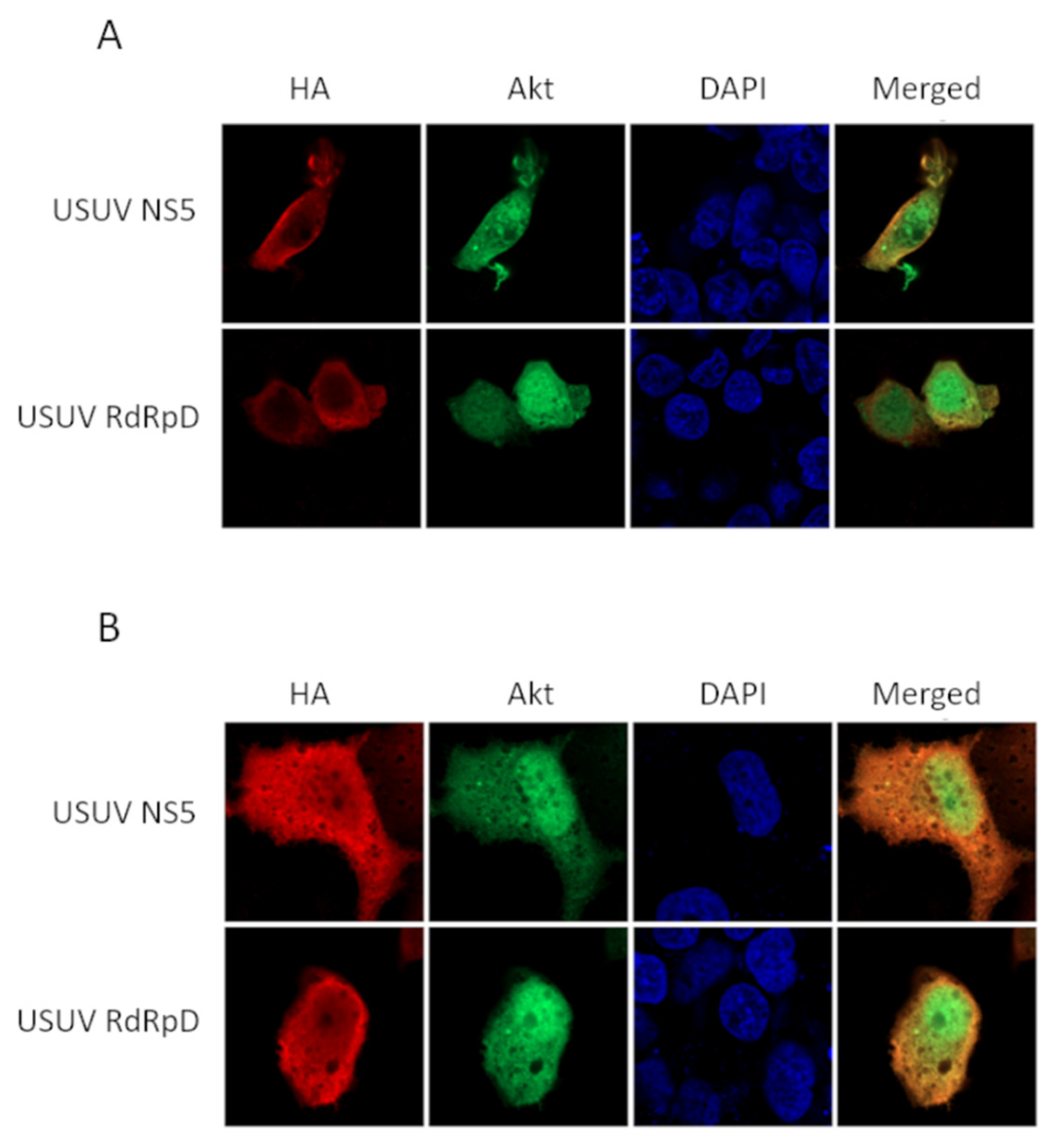
2.1. NS5 and Its RdRp Domain Are Phosphorylated by and Interact with Cellular Akt
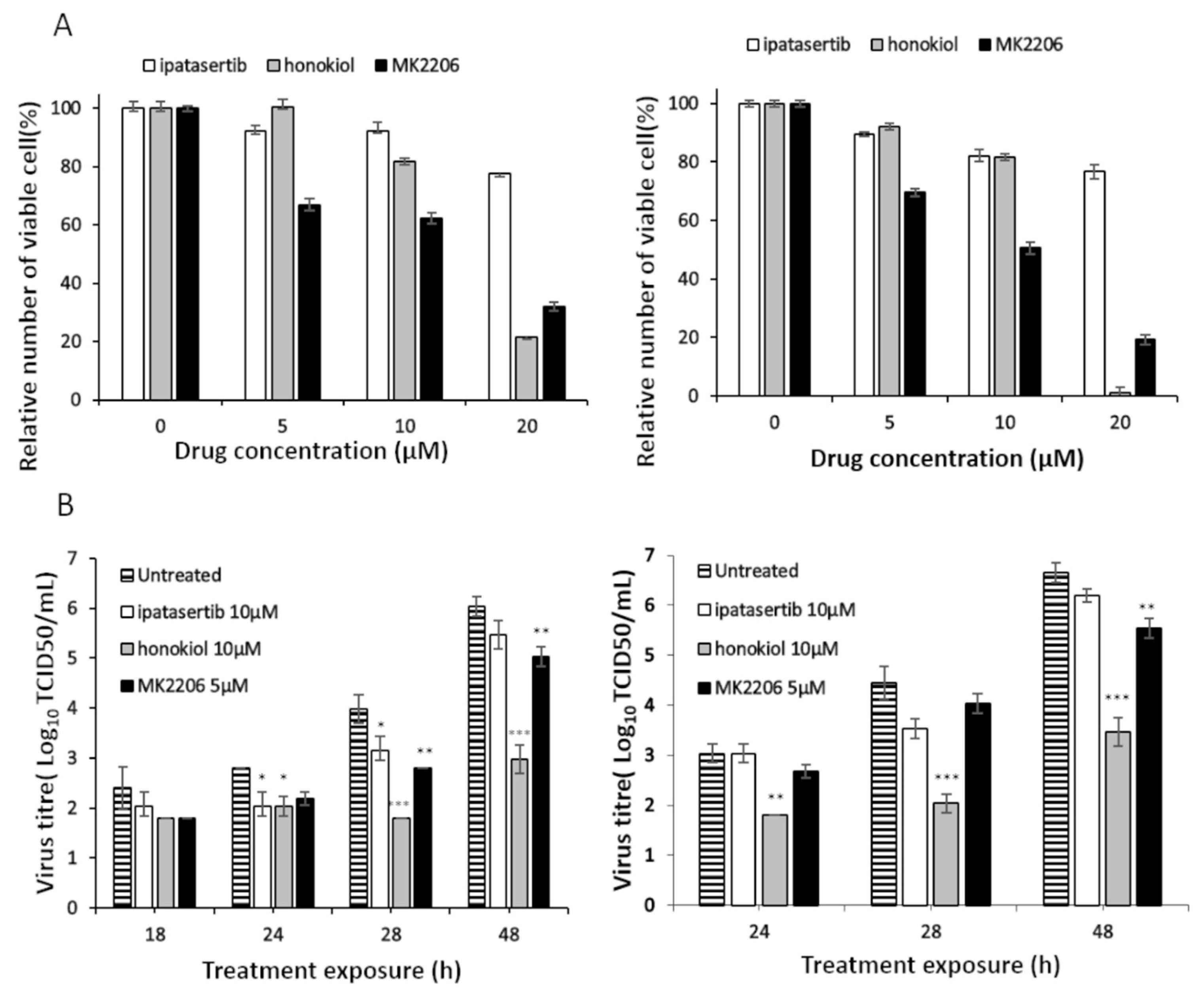
2.2. Effect of Akt Inhibition on USUV Replication
3. Discussion
4. Materials and Methods
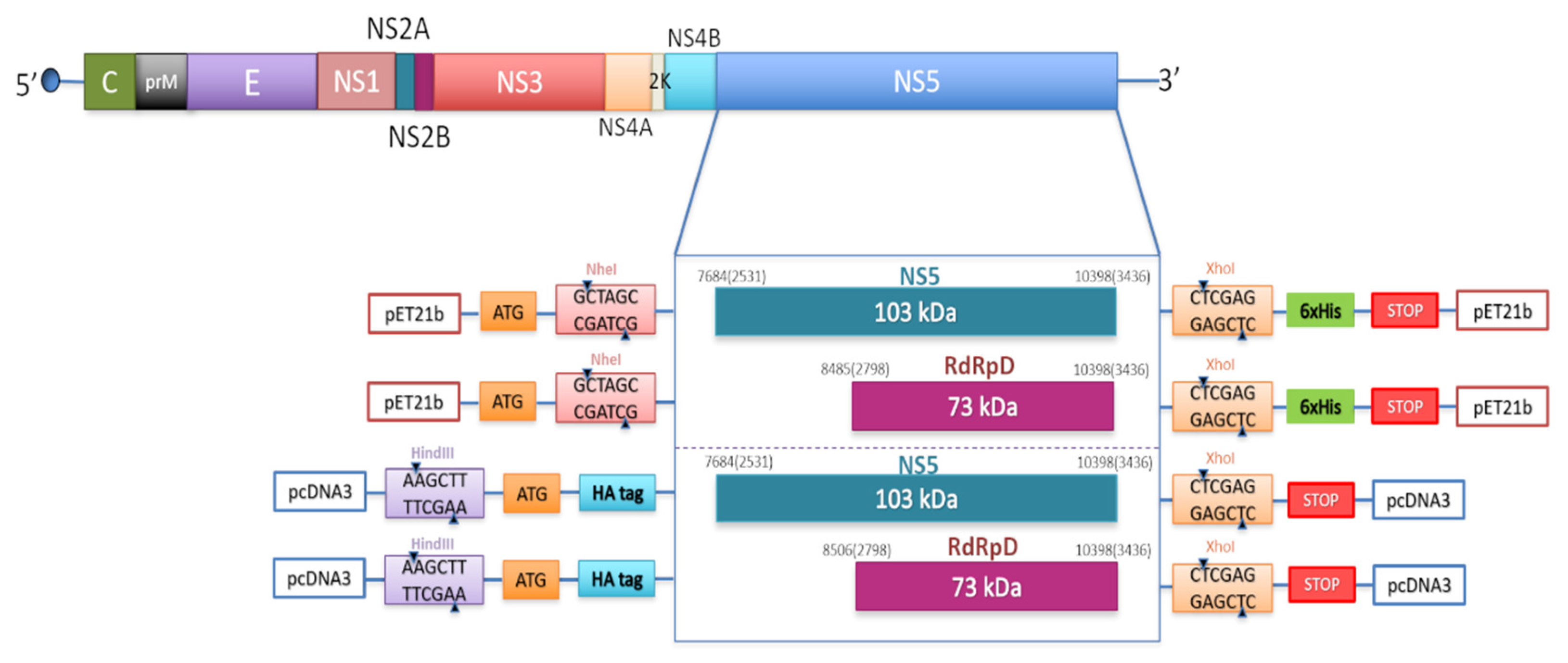
4.1. DNA Amplification by PCR, Cloning, and Purification
4.2. In Vitro Phosphorylation by Akt
4.3. Western-Blot and Co-Immunoprecipitation
4.4. Immunocytochemistry and Confocal Microscopy
4.5. USUVand Protocols for Infection
4.6. Virus Titration
4.7. Cell Viability
4.8. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flint, J.; Rall, G.F.; Racaniello, V.R.; Skalka, A.M. Principles of Virology, Volume II: Pathogenesis & Control; American Society for Microbiology: Washington, DC, USA, 2015. [Google Scholar]
- Wu, J.; Liu, W.; Gong, P. A Structural Overview of RNA-Dependent RNA Polymerases from the Flaviviridae Family. Int. J. Mol. Sci. 2015, 16, 12943–12957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierson, T.C.; Diamond, M.S. The continued threat of emerging flaviviruses. Nat. Microbiol. 2020, 5, 796–812. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, S.; Kuhn, R.J.; Rossmann, M.G. A structural perspective of the flavivirus life cycle. Nat. Rev. Genet. 2005, 3, 13–22. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, E.; Li, G. Approved Antiviral Drugs over the Past 50 Years. Clin. Microbiol. Rev. 2016, 29, 695–747. [Google Scholar] [CrossRef] [Green Version]
- Eyer, L.; Nencka, R.; De Clercq, E.; Seley-Radtke, K.; Růžek, D. Nucleoside analogs as a rich source of antiviral agents active against arthropod-borne flaviviruses. Antivir. Chem. Chemother. 2018, 26, 2040206618761299. [Google Scholar] [CrossRef] [PubMed]
- Guarner, J.; Hale, G.L. Four human diseases with significant public health impact caused by mosquito-borne flaviviruses: West Nile, Zika, dengue and yellow fever. Semin. Diagn. Pathol. 2019, 36, 170–176. [Google Scholar] [CrossRef]
- Nikolay, B.; Diallo, M.; Boye, C.S.B.; Sall, A.A. Usutu Virus in Africa. Vector-Borne Zoonotic Dis. 2011, 11, 1417–1423. [Google Scholar] [CrossRef]
- Pecorari, M.; Longo, G.; Gennari, W.; Grottola, A.; Sabbatini, A.M.; Tagliazucchi, S.; Savini, G.; Monaco, F.; Simone, M.; Lelli, R.; et al. First human case of Usutu virus neuroinvasive infection, Italy, August–September 2009. Eurosurveillance 2009, 14, 19446. [Google Scholar]
- Vilibic-Cavlek, T.; Savic, V.; Sabadi, D.; Peric, L.; Barbic, L.; Klobucar, A.; Miklausic, B.; Tabain, I.; Santini, M.; Vucelja, M.; et al. Prevalence and molecular epidemiology of West Nile and Usutu virus infections in Croatia in the ‘One health’ context, 2018. Transbound. Emerg. Dis. 2019, 66, 1946–1957. [Google Scholar] [CrossRef]
- Benzarti, E.; Garigliany, M. In Vitro and In Vivo Models to Study the Zoonotic Mosquito-Borne Usutu Virus. Viruses 2020, 12, 1116. [Google Scholar] [CrossRef]
- Vilibic-Cavlek, T.; Petrovic, T.; Savic, V.; Barbic, L.; Tabain, I.; Stevanovic, V.; Klobucar, A.; Mrzljak, A.; Ilic, M.; Bogdanic, M.; et al. Epidemiology of Usutu Virus: The European Scenario. Pathogens 2020, 9, 699. [Google Scholar] [CrossRef]
- Gould, E.; Pettersson, J.; Higgs, S.; Charrel, R.; De Lamballerie, X. Emerging arboviruses: Why today? One Health 2017, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Barzon, L. Ongoing and emerging arbovirus threats in Europe. J. Clin. Virol. 2018, 107, 38–47. [Google Scholar] [CrossRef]
- Roesch, F.; Fajardo, A.; Moratorio, G.; Vignuzzi, M. Usutu Virus: An Arbovirus on the Rise. Viruses 2019, 11, 640. [Google Scholar] [CrossRef] [Green Version]
- Vazquez, A.; Jimenez-Clavero, M.A.; Franco, L.; Donoso-Mantke, O.; Sambri, V.; Niedrig, M.; Zeller, H.; Tenorio, A. Usutu virus: Potential risk of human disease in Europe. Eurosurveillance 2011, 16, 19935. [Google Scholar] [PubMed]
- Neufeldt, C.J.; Cortese, M.; Acosta, E.G.; Bartenschlager, R. Rewiring cellular networks by members of the Flaviviridae family. Nat. Rev. Genet. 2018, 16, 125–142. [Google Scholar] [CrossRef]
- Lim, S.P.; Noble, C.G.; Shi, P.-Y. The dengue virus NS5 protein as a target for drug discovery. Antivir. Res. 2015, 119, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yang, C.; Zhang, W.; Mahalingam, S.; Wang, M.; Cheng, A. Flaviviridae virus nonstructural proteins 5 and 5A mediate viral immune evasion and are promising targets in drug development. Pharmacol. Ther. 2018, 190, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agudo, R.; Arias, A.; Domingo, E. 5-fluorouracil in lethal mutagenesis of foot-and-mouth disease virus. Futur. Med. Chem. 2009, 1, 529–539. [Google Scholar] [CrossRef]
- Más, A.; López-Galíndez, C.; Cacho, I.; Gómez, J.; Martínez, M.A. Unfinished Stories on Viral Quasispecies and Darwinian Views of Evolution. J. Mol. Biol. 2010, 397, 865–877. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Sharma, S.; Kumar, R.; Tripathi, B.N.; Barua, S.; Ly, H.; Rouse, B.T. Host-Directed Antiviral Therapy. Clin. Microbiol. Rev. 2020, 33, 3. [Google Scholar] [CrossRef] [PubMed]
- Saiz, J.-C.; De Oya, N.J.; Blázquez, A.-B.; Escribano-Romero, E.; Martín-Acebes, M.A. Host-Directed Antivirals: A Realistic Alternative to Fight Zika Virus. Viruses 2018, 10, 453. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Na, L.; Li, Y.; Chen, L. Roles of the PI3K/AKT/mTOR signalling pathways in neurodegenerative diseases and tumours. Cell Biosci. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valero, M.L.; Sabariegos, R.; Cimas, F.J.; Perales, C.; Domingo, E.; Sánchez-Prieto, R.; Mas, A. Hepatitis C Virus RNA-Dependent RNA Polymerase Interacts with the Akt/PKB Kinase and Induces Its Subcellular Relocalization. Antimicrob. Agents Chemother. 2016, 60, 3540–3550. [Google Scholar] [CrossRef] [Green Version]
- Osaki, M.; Oshimura, M.; Ito, H. PI3K-Akt pathway: Its functions and alterations in human cancer. Apoptosis 2004, 9, 667–676. [Google Scholar] [CrossRef]
- Alessi, D.R.; Cohen, P. Mechanism of activation and function of protein kinase B. Curr. Opin. Genet. Dev. 1998, 8, 55–62. [Google Scholar] [CrossRef]
- Liu, Z.; Tian, Y.; Machida, K.; Lai, M.M.C.; Luo, G.; Foung, S.K.H.; Ou, J.-H.J. Transient Activation of the PI3K-AKT Pathway by Hepatitis C Virus to Enhance Viral Entry. J. Biol. Chem. 2012, 287, 41922–41930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Liu, H.; Zou, J.; Zhang, B.; Yuan, Z. Dengue virus subgenomic RNA induces apoptosis through the Bcl-2-mediated PI3k/Akt signaling pathway. Virology 2014, 448, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.-J.; Liao, C.-L.; Lin, Y.-L. Flavivirus Activates Phosphatidylinositol 3-Kinase Signaling To Block Caspase-Dependent Apoptotic Cell Death at the Early Stage of Virus Infection. J. Virol. 2005, 79, 8388–8399. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Chakraborty, S.; Basu, A. Critical role of lipid rafts in virus entry and activation of phosphoinositide 3′ kinase/Akt signaling during early stages of Japanese encephalitis virus infection in neural stem/progenitor cells. J. Neurochem. 2010, 115, 537–549. [Google Scholar] [CrossRef]
- Albentosa-González, L.; Clemente-Casares, P.; Sabariegos, R.; Mas, A. Polymerase Activity, Protein-Protein Interaction, and Cellular Localization of the Usutu Virus NS5 Protein. Antimicrob. Agents Chemother. 2019, 64, e01573-19. [Google Scholar] [CrossRef] [Green Version]
- Munakata, T.; Liang, Y.; Kim, S.; McGivern, D.R.; Huibregtse, J.; Nomoto, A.; Lemon, S.M. Hepatitis C Virus Induces E6AP-Dependent Degradation of the Retinoblastoma Protein. PLOS Pathog. 2007, 3, e139. [Google Scholar] [CrossRef] [Green Version]
- Kukihara, H.; Moriishi, K.; Taguwa, S.; Tani, H.; Abe, T.; Mori, Y.; Suzuki, T.; Fukuhara, T.; Taketomi, A.; Maehara, Y.; et al. Human VAP-C Negatively Regulates Hepatitis C Virus Propagation. J. Virol. 2009, 83, 7959–7969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, L.V.; Ngo, H.T.; Lim, Y.-S.; Hwang, S.B. Hepatitis C virus non-structural 5B protein interacts with cyclin A2 and regulates viral propagation. J. Hepatol. 2012, 57, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Pfannkuche, A.; Büther, K.; Karthe, J.; Poenisch, M.; Bartenschlager, R.; Trilling, M.; Hengel, H.; Willbold, D.; Häussinger, D.; Bode, J.G. C-Src is required for complex formation between the hepatitis C virus-encoded proteins NS5A and NS5B: A prerequisite for replication. Hepatology 2011, 53, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Aizaki, H.; Hara, H.; Matsuda, M.; Ando, T.; Shimoji, T.; Murakami, K.; Masaki, T.; Shoji, I.; Homma, S.; et al. Chaperonin TRiC/CCT participates in replication of hepatitis C virus genome via interaction with the viral NS5B protein. Virology 2011, 410, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Kusakawa, T.; Shimakami, T.; Kaneko, S.; Yoshioka, K.; Murakami, S. Functional Interaction of Hepatitis C Virus NS5B with Nucleolin GAR Domain. J. Biochem. 2007, 141, 917–927. [Google Scholar] [CrossRef] [Green Version]
- Hillung, J.; Ruiz-López, E.; Bellón-Echeverría, I.; Clemente-Casares, P.; Mas, A. Characterization of the interaction between hepatitis C virus NS5B and the human oestrogen receptor alpha. J. Gen. Virol. 2012, 93, 780–785. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Chen, Y.-H.; Chow, L.-P.; Tsai, Y.-H.; Chen, P.-H.; Huang, C.-Y.F.; Chen, W.-T.; Hwang, L.-H. Heat Shock Protein 72 Is Associated with the Hepatitis C Virus Replicase Complex and Enhances Viral RNA Replication. J. Biol. Chem. 2010, 285, 28183–28190. [Google Scholar] [CrossRef] [Green Version]
- Morohashi, K.; Sahara, H.; Watashi, K.; Iwabata, K.; Sunoki, T.; Kuramochi, K.; Takakusagi, K.; Miyashita, H.; Sato, N.; Tanabe, A.; et al. Cyclosporin A Associated Helicase-Like Protein Facilitates the Association of Hepatitis C Virus RNA Polymerase with Its Cellular Cyclophilin B. PLoS ONE 2011, 6, e18285. [Google Scholar] [CrossRef]
- Fernandes, F.; Poole, D.S.; Hoover, S.; Middleton, R.; Andrei, A.-C.; Gerstner, J.; Striker, R. Sensitivity of hepatitis C virus to cyclosporine A depends on nonstructural proteins NS5A and NS5B. Hepatology 2007, 46, 1026–1033. [Google Scholar] [CrossRef]
- Qing, M.; Yang, F.; Zhang, B.; Zou, G.; Robida, J.M.; Yuan, Z.; Tang, H.; Shi, P.-Y. Cyclosporine Inhibits Flavivirus Replication through Blocking the Interaction between Host Cyclophilins and Viral NS5 Protein. Antimicrob. Agents Chemother. 2009, 53, 3226–3235. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Chen, Z.; Zhang, B.; Miao, H.; Zohaib, A.; Xu, Q.; Chen, H.; Cao, S. Heat Shock Protein 70 Is Associated with Replicase Complex of Japanese Encephalitis Virus and Positively Regulates Viral Genome Replication. PLoS ONE 2013, 8, e75188. [Google Scholar] [CrossRef] [Green Version]
- Khadka, S.; Vangeloff, A.D.; Zhang, C.; Siddavatam, P.; Heaton, N.S.; Wang, L.; Sengupta, R.; Sahasrabudhe, S.; Randall, G.; Gribskov, M.; et al. A Physical Interaction Network of Dengue Virus and Human Proteins. Mol. Cell. Proteom. 2011, 10, M111.012187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Di, D.; Huang, H.; Wang, X.; Xia, Q.; Ma, X.; Liu, K.; Li, B.; Shao, D.; Qiu, Y.; et al. NS5-V372A and NS5-H386Y variations are responsible for differences in interferon alpha/beta induction and co-contribute to the replication advantage of Japanese encephalitis virus genotype I over genotype III in ducklings. PLoS Pathog. 2020, 16, e1008773. [Google Scholar] [CrossRef] [PubMed]
- Harrison, A.R.; Moseley, G.W. The Dynamic Interface of Viruses with STATs. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Roby, J.A.; Esser-Nobis, K.; Dewey-Verstelle, E.C.; Fairgrieve, M.R.; Schwerk, J.; Lu, A.Y.; Soveg, F.W.; Hemann, E.A.; Hatfield, L.D.; Keller, B.C.; et al. Flavivirus Nonstructural Protein NS5 Dysregulates HSP90 to Broadly Inhibit JAK/STAT Signaling. Cells 2020, 9, 899. [Google Scholar] [CrossRef] [PubMed]
- Thurmond, S.; Wang, B.; Song, J.; Hai, R. Suppression of Type I Interferon Signaling by Flavivirus NS5. Viruses 2018, 10, 712. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Thurmond, S.; Zhou, K.; Sánchez-Aparicio, M.T.; Fang, J.; Lu, J.; Gao, L.; Ren, W.; Cui, Y.; Veit, E.C.; et al. Structural basis for STAT2 suppression by flavivirus NS5. Nat. Struct. Mol. Biol. 2020, 27, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Bühler, S.; Selisko, B.; Davidson, A.; Mulder, K.; Canard, B.; Miller, S.; Bartenschlager, R. Nuclear Localization of Dengue Virus Nonstructural Protein 5 Does Not Strictly Correlate with Efficient Viral RNA Replication and Inhibition of Type I Interferon Signaling. J. Virol. 2013, 87, 4545–4557. [Google Scholar] [CrossRef] [Green Version]
- Owen, K.L.; Brockwell, N.K.; Parker, B.S. JAK-STAT Signaling: A Double-Edged Sword of Immune Regulation and Cancer Progression. Cancers 2019, 11, 2002. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, J.S.; Rosler, K.M.; Harrison, D.A. The JAK/STAT signaling pathway. J. Cell Sci. 2004, 117, 1281–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Rehman, S.K.; Zhang, W.; Wen, A.; Yao, L.; Zhang, J. Autophagy is a therapeutic target in anticancer drug resistance. Biochim. Biophys. Acta (BBA)-Bioenergy 2010, 1806, 220–229. [Google Scholar] [CrossRef]
- Collins, D.C.; Chenard-Poirier, M.; Lopez, J.S.; Collins, M.C.-P.A.J.L.D. The PI3K Pathway at the Crossroads of Cancer and the Immune System: Strategies for Next Generation Immunotherapy Combinations. Curr. Cancer Drug Targets 2018, 18, 355–364. [Google Scholar] [CrossRef]
- Jhanwar-Uniyal, M.; Wainwright, J.V.; Mohan, A.L.; Tobias, M.E.; Murali, R.; Gandhi, C.D.; Schmidt, M.H. Diverse signaling mechanisms of mTOR complexes: mTORC1 and mTORC2 in forming a formidable relationship. Adv. Biol. Regul. 2019, 72, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Pandolfi, P.P. The PTEN(-)PI3K Axis in Cancer. Biomolecules 2019, 9, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simioni, C.; Martelli, A.M.; Zauli, G.; Vitale, M.; McCubrey, J.A.; Capitani, S.; Neri, L.M. Targeting the phosphatidylinositol 3-kinase/Akt/mechanistic target of rapamycin signaling pathway in B-lineage acute lymphoblastic leukemia: An update. J. Cell. Physiol. 2018, 233, 6440–6454. [Google Scholar] [CrossRef] [Green Version]
- Vergadi, E.; Ieronymaki, E.; Lyroni, K.; Vaporidi, K.; Tsatsanis, C. Akt Signaling Pathway in Macrophage Activation and M1/M2 Polarization. J. Immunol. 2017, 198, 1006–1014. [Google Scholar] [CrossRef] [Green Version]
- Weichhart, T.; Saemann, M.D. The PI3K/Akt/mTOR pathway in innate immune cells: Emerging therapeutic applications. Ann. Rheum. Dis. 2008, 67, iii70–iii74. [Google Scholar] [CrossRef]
- Xu, Z.; Han, X.; Ou, D.; Liu, T.; Li, Z.; Jiang, G.; Liu, J.; Zhang, J. Targeting PI3K/AKT/mTOR-mediated autophagy for tumor therapy. Appl. Microbiol. Biotechnol. 2020, 104, 575–587. [Google Scholar] [CrossRef]
- Ceresa, B.P.; Kao, A.W.; Santeler, S.R.; Pessin, J.E. Inhibition of Clathrin-Mediated Endocytosis Selectively Attenuates Specific Insulin Receptor Signal Transduction Pathways. Mol. Cell. Biol. 1998, 18, 3862–3870. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.Y.; Huang, W.R.; Chi, P.I.; Chiu, H.C.; Liu, H.J. Cell entry of bovine ephemeral fever virus requires activation of Src-JNK-AP1 and PI3K-Akt-NF-kappaB pathways as well as Cox-2-mediated PGE2 /EP receptor signalling to enhance clathrin-mediated virus endocytosis. Cell Microbiol. 2015, 17, 967–987. [Google Scholar] [CrossRef] [PubMed]
- Holla, P.; Ahmad, I.; Ahmed, Z.; Jameel, S. Hepatitis E Virus Enters Liver Cells Through a Dynamin-2, Clathrin and Membrane Cholesterol-Dependent Pathway. Traffic 2015, 16, 398–416. [Google Scholar] [CrossRef] [PubMed]
- Je, H.-S.; Zhou, J.; Yang, F.; Lu, B. Distinct Mechanisms for Neurotrophin-3-Induced Acute and Long-Term Synaptic Potentiation. J. Neurosci. 2005, 25, 11719–11729. [Google Scholar] [CrossRef]
- Reis, C.R.; Chen, P.H.; Srinivasan, S.; Aguet, F.; Mettlen, M.; Schmid, S.L. Crosstalk between Akt/GSK3beta signaling and dynamin-1 regulates clathrin-mediated endocytosis. EMBO J. 2015, 34, 2132–2146. [Google Scholar] [CrossRef] [Green Version]
- Smillie, K.J.; Cousin, M.A. Akt/PKB Controls the Activity-Dependent Bulk Endocytosis of Synaptic Vesicles. Traffic 2012, 13, 1004–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eden, J.-S.; Sharpe, L.J.; White, P.A.; Brown, A.J. Norovirus RNA-Dependent RNA Polymerase Is Phosphorylated by an Important Survival Kinase, Akt. J. Virol. 2011, 85, 10894–10898. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-J.; Kim, J.-H.; Kim, Y.-G.; Lim, H.-S.; Oh, J.-W. Protein Kinase C-related Kinase 2 Regulates Hepatitis C Virus RNA Polymerase Function by Phosphorylation. J. Biol. Chem. 2004, 279, 50031–50041. [Google Scholar] [CrossRef] [Green Version]
- Han, S.H.; Kim, S.-J.; Kim, E.-J.; Kim, T.-E.; Moon, J.-S.; Kim, G.-W.; Lee, S.-H.; Cho, K.; Yoo, J.S.; Son, W.S.; et al. Phosphorylation of Hepatitis C Virus RNA Polymerases Ser29 and Ser42 by Protein Kinase C-Related Kinase 2 Regulates Viral RNA Replication. J. Virol. 2014, 88, 11240–11252. [Google Scholar] [CrossRef] [Green Version]
- Blake, J.F.; Xu, R.; Bencsik, J.R.; Xiao, D.; Kallan, N.C.; Schlachter, S.; Mitchell, I.S.; Spencer, K.L.; Banka, A.L.; Wallace, E.M.; et al. Discovery and Preclinical Pharmacology of a Selective ATP-Competitive Akt Inhibitor (GDC-0068) for the Treatment of Human Tumors. J. Med. Chem. 2012, 55, 8110–8127. [Google Scholar] [CrossRef]
- Hirai, H.; Sootome, H.; Nakatsuru, Y.; Miyama, K.; Taguchi, S.; Tsujioka, K.; Ueno, Y.; Hatch, H.; Majumder, P.K.; Pan, B.-S.; et al. MK-2206, an Allosteric Akt Inhibitor, Enhances Antitumor Efficacy by Standard Chemotherapeutic Agents or Molecular Targeted Drugs In vitro and In vivo. Mol. Cancer Ther. 2010, 9, 1956–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherrin, C.; Haskell, K.; Howell, B.; Jones, R.; Leander, K.; Robinson, R.; Watkins, A.; Bilodeau, M.; Hoffman, J.; Sanderson, P.; et al. An allosteric Akt inhibitor effectively blocks Akt signaling and tumor growth with only transient effects on glucose and insulin levels in vivo. Cancer Biol. Ther. 2010, 9, 493–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banik, K.; Ranaware, A.M.; Deshpande, V.; Nalawade, S.P.; Padmavathi, G.; Bordoloi, D.; Sailo, B.L.; Shanmugam, M.K.; Fan, L.; Arfuso, F.; et al. Honokiol for cancer therapeutics: A traditional medicine that can modulate multiple oncogenic targets. Pharmacol. Res. 2019, 144, 192–209. [Google Scholar] [CrossRef] [PubMed]
- Jiao, P.; Zhou, Y.-S.; Yang, J.-X.; Zhao, Y.-L.; Liu, Q.-Q.; Yuan, C.; Wang, F.-Z. MK-2206 induces cell cycle arrest and apoptosis in HepG2 cells and sensitizes TRAIL-mediated cell death. Mol. Cell. Biochem. 2013, 382, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Bassi, M.R.; Sempere, R.N.; Meyn, P.; Polacek, C.; Arias, A. Extinction of Zika Virus and Usutu Virus by Lethal Mutagenesis Reveals Different Patterns of Sensitivity to Three Mutagenic Drugs. Antimicrob. Agents Chemother. 2018, 62, e00380-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissenbock, H.; Kolodziejek, J.; Url, A.; Lussy, H.; Rebel-Bauder, B.; Nowotny, N. Emergence of Usutu virus, an African mosquito-borne flavivirus of the Japanese encephalitis virus group, central Europe. Emerg. Infect. Dis. 2002, 8, 652–656. [Google Scholar] [CrossRef] [PubMed]
- Sempere, R.N.; Arias, A. Establishment of a Cell Culture Model of Persistent Flaviviral Infection: Usutu Virus Shows Sustained Replication during Passages and Resistance to Extinction by Antiviral Nucleosides. Viruses 2019, 11, 560. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′->3′) 1 | 5′-end Position 2 |
|---|---|---|
| pET-USUV-NS5_F | GGCGGCTAGCGGAAGACCAGGAGGAAGGAC | 7684 |
| pET-USUV-RdRp-F | GGCGGCTAGCGGGAAGCCCCAGCCACATAC | 8485 |
| pET-USUV-R | GCGGCTCGAGCAAAACCCTGTCCTCCTGGAC | 10,398 |
| pcDNA-USUV-NS5-F | AAGCTTGCCATGTACCCATACGATGTTCCAGATTACGCTGGAAGACCAGGAGGAAGGAC | 7684 |
| pcDNA-USUV-R | CTCGAGTTACAAAACCCTGTCCTCCTGGAC | 10,398 |
| pcDNA-USUV-RdRp-F | AAGCTTGCCATGTACCCATACGATGTTCCAGATTACGCTGGGAAGCCCCAGCCACATAC | 8485 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albentosa-González, L.; Sabariegos, R.; Arias, A.; Clemente-Casares, P.; Mas, A. Akt Interacts with Usutu Virus Polymerase, and Its Activity Modulates Viral Replication. Pathogens 2021, 10, 244. https://doi.org/10.3390/pathogens10020244
Albentosa-González L, Sabariegos R, Arias A, Clemente-Casares P, Mas A. Akt Interacts with Usutu Virus Polymerase, and Its Activity Modulates Viral Replication. Pathogens. 2021; 10(2):244. https://doi.org/10.3390/pathogens10020244
Chicago/Turabian StyleAlbentosa-González, Laura, Rosario Sabariegos, Armando Arias, Pilar Clemente-Casares, and Antonio Mas. 2021. "Akt Interacts with Usutu Virus Polymerase, and Its Activity Modulates Viral Replication" Pathogens 10, no. 2: 244. https://doi.org/10.3390/pathogens10020244