
The carB Gene of Escherichia coli BL21(DE3) is Associated with Nematicidal Activity against the Root-Knot Nematode Meloidogyne javanica

 ,
,
Abstract
:1. Introduction
2. Results
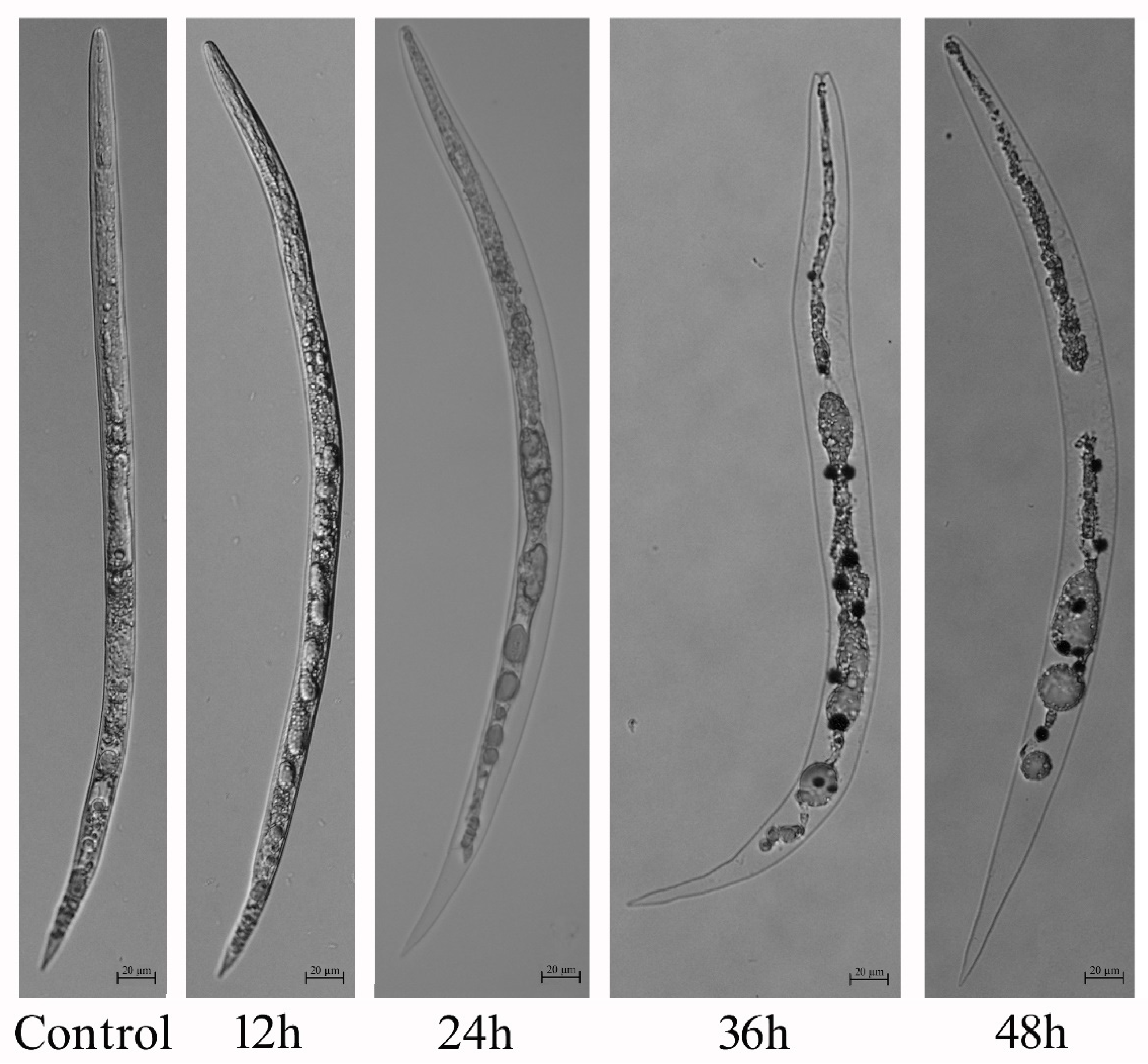
2.1. In Vitro Nematicidal Activity
2.2. Nematicidal Active Stability
2.3. Glasshouse Experiments
2.4. Genetics of Nematicidal Activity
3. Discussion
4. Materials and Methods
4.1. Strains and Plasmids
4.2. Preparation of Meloidogyne javanica
4.3. Antagonism of Strain BL21(DE3) against M. javanica In Vivo
4.4. Different Treatments of BL21(DE3) Supernatants
4.5. Antagonism of Strain BL21(DE3) against M. javanica In Vivo
4.6. DNA Manipulation and Transformation
4.7. Determination of the Transposon Insertion Site
4.8. Construction of Complementation Plasmid
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, W.; Sun, J.; Cui, J.; Wang, G.; Kong, L.; Peng, H.; Chen, S.; Peng, D. Efficacy evaluation of fungus Syncephalastrum racemosum and nematicide avermectin against the root-knot nematode Meloidogyne incognita on cucumber. PLoS ONE 2014, 9, e89717. [Google Scholar] [CrossRef] [Green Version]
- Sikora, R.; Pocasangre, L.; zum Felde, A.; Niere, B.; Vu, T.T.; Dababat, A.A. Mutualistic endophytic fungi and in-planta suppressiveness to plant parasitic nematodes. Biol. Control. 2008, 46, 15–23. [Google Scholar] [CrossRef]
- Nicol, J.M.; Turner, S.J.; Coyne, D.L.; Nijs, L.; Hockland, S.; Maafi, Z.T. Current nematode threats to world agriculture. In Genomics and Molecular Genetics of Plant-Nematode Interactions; Jones, J., Gheysen, G., Fenoll, C., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 21–43. [Google Scholar]
- Siddiqui, Z.A.; Mahmood, I. Role of bacteria in the management of plant parasitic nematodes: A review. Bioresource Technol. 1999, 69, 167–179. [Google Scholar] [CrossRef]
- Li, J.; Zou, C.; Xu, J.; Ji, X.; Niu, X.; Yang, J.; Huang, X.; Zhang, K. Molecular mechanisms of nematode-nematophagous microbe interactions: Basis for biological control of plant-parasitic nematodes. Annu. Rev. Phytopathol. 2015, 53, 67–95. [Google Scholar] [CrossRef] [PubMed]
- Colagiero, M.; Rosso, L.C.; Ciancio, A. Diversity and biocontrol potential of bacterial consortia associated to root-knot nematodes. Biol. Control. 2018, 120, 11–16. [Google Scholar] [CrossRef]
- Xiang, N.; Lawrence, K.S.; Donald, P.A. Biological control potential of plant growth-promoting rhizobacteria suppression of Meloidogyne incognita on cotton and Heterodera glycines on soybean: A review. J. Phytopathol. 2018, 166, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Dickson, D.W.; McSorley, R.; Mitchell, D.J.; Hewlett, T.E. Suppression of Meloidogyne arenaria race 1 by soil application of endospores of Pasteuria penetrans. J. Nematol. 1996, 28, 159–168. [Google Scholar] [PubMed]
- Kokalis-Burelle, N. Pasteuria penetrans for control of Meloidogyne incognita on tomato and cucumber, and M. arenaria on snapdragon. J. Nematol. 2015, 47, 207–213. [Google Scholar]
- Cronin, D.; Moenne-Loccoz, Y.; Fenton, A.; Dunne, C.; Dowling, D.N.; O’gara, F. Role of 2, 4-diacetylphloroglucinol in the interactions of the biocontrol pseudomonad strain F113 with the potato cyst nematode Globodera rostochiensis. Appl. Environ. Microb. 1997, 63, 1357–1361. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, I.A.; Shahid Shaukat, S. Suppression of root-knot disease by Pseudomonas fluorescens CHA0 in tomato: Importance of bacterial secondary metabolite, 2, 4-diacetylpholo- roglucinol. Soil Biol. Biochem. 2003, 35, 1615–1623. [Google Scholar] [CrossRef]
- Niu, Q.; Huang, X.; Tian, B.; Yang, J.; Liu, J.; Zhang, L.; Zhang, K. Bacillus sp. B16 kills nematodes with a serine protease identified as a pathogenic factor. Appl. Microbiol. Biot. 2005, 69, 722–730. [Google Scholar] [CrossRef]
- Niu, Q.; Huang, X.; Zhang, L.; Xu, J.; Yang, D.; Wei, K.; Niu, X.; An, Z.; Bennett, J.; Zou, C.; et al. A Trojan horse mechanism of bacterial pathogenesis against nematodes. Proc. Natl. Acad. Sci. USA 2010, 107, 16631–16636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, Q.; Huang, X.; Zhang, L.; Li, Y.; Li, J.; Yang, J.; Zhang, K. A neutral protease from Bacillus nematocida, another potential virulence factor in the infection against nematodes. Arch. Microbiol. 2006, 185, 439–448. [Google Scholar] [CrossRef]
- Bravo, A.; Gómez, I.; Porta, H.; García-Gómez, B.I.; Rodriguez-Almazan, C.; Pardo, L.; Soberón, M. Evolution of Bacillus thuringiensis Cry toxins insecticidal activity. Microb. Biotechnol. 2012, 6, 17–26. [Google Scholar] [CrossRef]
- Reitz, M.; Rudolph, K.; Schroder, I.; Hoffmann-Hergarten, S.; Hallmann, J.; Sikora, R.A. Lipopolysaccharides of Rhizobium etli strain G12 act in potato roots as an inducing agent of systemic resistance to infection by the cyst nematode Globodera pallida. Appl. Environ. Microb. 2000, 66, 3515–3518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.; Li, S.; Liu, X.; Zhang, C.; Xu, J.; Chen, Y. Bacillus halotolerans strain LYSX1-induced systemic resistance against the root-knot nematode Meloidogyne javanica in tomato. Ann. Microbiol. 2019, 69, 1227–1233. [Google Scholar] [CrossRef]
- Samuel, B.S.; Rowedder, H.; Braendle, C.; Félix, M.-A.; Ruvkun, G. Caenorhabditis elegans responses to bacteria from its natural habitats. Proc. Natl. Acad. Sci. USA 2016, 113, E3941–E3949. [Google Scholar] [CrossRef] [Green Version]
- Anyanful, A.; Dolan-Livengood, J.M.; Lewis, T.; Sheth, S.; DeZalia, M.N.; Sherman, M.A.; Kalman, L.V.; Benian, G.M.; Kalman, D. Paralysis and killing of Caenorhabditis elegans by enteropathogenic Escherichia coli requires the bacterial tryptophanase gene. Mol. Microbiol. 2005, 57, 988–1007. [Google Scholar] [CrossRef]
- Chou, T.-C.; Chiu, H.-C.; Kuo, C.-J.; Wu, C.-M.; Syu, W.-J.; Chiu, W.-T.; Chen, C.-S. Enterohaemorrhagic Escherichia coli O157:H7 Shiga-like toxin 1 is required for full pathogenicity and activation of the p38 mitogen-activated protein kinase pathway in Caenorhabditis elegans. Cell Microbiol. 2013, 15, 82–97. [Google Scholar] [CrossRef]
- Maghodia, A.B.; Spiegel, Y.; Sela, S. Interactions between Escherichia coli and the plant-parasitic nematode Meloidogyne javanica. J. Appl. Microbiol. 2008, 105, 1810–1816. [Google Scholar] [CrossRef]
- Studier, F.W.; Moffatt, B.A. Use of bacteriophage T7 RNA polymerase to direct selective high-level expression of cloned genes. J. Mol. Biol. 1986, 189, 113–130. [Google Scholar] [CrossRef]
- Ito, M.; Kim, Y.G.; Tsuji, H.; Kiwaki, M.; Nomoto, K.; Tanaka, R.; Okada, N.; Danbara, H. A practical random mutagenesis system for probiotic Lactobacillus casei using Tn5 transposition complexes. J. Appl. Microbiol. 2010, 109, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Zhang, W.; Lei, L.; Liu, X.; Wei, H. Genome-wide investigation of the genes involved in nicotine metabolism in Pseudomonas putida J5 by Tn5 transposon mutagenesis. Appl. Microbiol. Biot. 2015, 99, 6503–6514. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Yuan, L.; Guo, W.; Li, Y.; Che, Y.; Zou, L.; Chen, G. Construction of a Tn5-tagged mutant library of Xanthomonas oryzae pv. oryzicola as an invaluable resource for functional genomics. Curr. Microbiol. 2011, 62, 908–916. [Google Scholar] [CrossRef] [PubMed]
- Holden, H.M.; Thoden, J.B.; Raushel, F.M. Carbamoyl phosphate synthetase: An amazing biochemical odyssey from substrate to product. Cell Mol. Life Sci. 1999, 56, 507–522. [Google Scholar] [CrossRef]
- Ali, N.I.; Siddiqui, I.A.; Shahid Shaukat, S.; Zaki, M.J. Nematicidal activity of some strains of Pseudomonas spp. Soil Biol. Biochem. 2002, 34, 1051–1058. [Google Scholar] [CrossRef]
- Mendoza, A.R.; Kiewnick, S.; Sikora, R.A. In vitro activity of Bacillus firmus against the burrowing nematode Radopholus similis, the root-knot nematode Meloidogyne incognita and the stem nematode Ditylenchus dipsaci. Biocontrol Sci. Techn. 2008, 18, 377–389. [Google Scholar] [CrossRef]
- Xia, Y.; Xie, S.; Ma, X.; Wu, H.; Wang, X.; Gao, X. The purL gene of Bacillus subtilis is associated with nematicidal activity. FEMS Microbiol. Lett. 2011, 322, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Wang, X.; Wang, K.; Su, L.; Li, H.; Li, R.; Shen, Q. The nematicidal effect of Camellia seed cake on root-knot nematode Meloidogyne javanica of banana. PLoS ONE 2015, 10, e0119700. [Google Scholar] [CrossRef] [Green Version]
- Meyer, S.L.F.; Halbrendt, J.M.; Carta, L.K.; Skantar, A.M.; Liu, T.; Abdelnabby, H.M.E.; Vinyard, B.T. Toxicity of 2,4-diacetylphloroglucinol (DAPG) to plant-parasitic and bacterial-feeding nematodes. J. Nematol. 2009, 41, 274–280. [Google Scholar]
- Aballay, E.; Prodan, S.; Zamorano, A.; Castaneda-Alvarez, C. Nematicidal effect of rhizobacteria on plant-parasitic nematodes associated with vineyards. World J. Microb. Biot. 2017, 33, 131. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Qi, G.; Yin, R.; Zhang, H.; Li, C.; Zhao, X. Bacillus cereus strain S2 shows high nematicidal activity against Meloidogyne incognita by producing sphingosine. Sci. Rep. 2016, 6, 28756. [Google Scholar] [CrossRef]
- Liu, Z.; Budiharjo, A.; Wang, P.; Shi, H.; Fang, J.; Borriss, R.; Zhang, K.; Huang, X. The highly modified microcin peptide plantazolicin is associated with nematicidal activity of Bacillus amyloliquefaciens FZB42. Appl. Microbiol. Biot. 2013, 97, 10081–10090. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Wang, J.; Song, Z.; Cheng, J.; Zhang, D.; Liu, Y. Nematicidal effects of 5-aminolevulinic acid on plant-parasitic nematodes. J. Nematol. 2017, 49, 295–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, T.; Tan, S.; Shen, Q.; Ran, W. Bacillus cereus X5 suppresses root-knot nematode of tomato by colonizing in roots and soil. Afr. J. Microbiol. Res. 2012, 6, 2321–2327. [Google Scholar] [CrossRef]
- Engelbrecht, G.; Horak, I.; Jansen van Rensburg, P.J.; Claassens, S. Bacillus-based bionematicides: Development, modes of action and commercialisation. Biocontrol Sci. Techn. 2018, 28, 629–653. [Google Scholar] [CrossRef]
- Aleshkin, G.I.; Kadzhaev, K.V.; Markov, A.P. High and low UV-dose responses in SOS-induction of the precise excision of transposons Tn1, Tn5 and Tn10 in Escherichia coli. Mutat. Res. Fund Mol. M. 1998, 401, 179–191. [Google Scholar] [CrossRef]
- Song, X.; Guo, J.; Ma, W.; Ji, Z.; Zou, L.; Chen, G.; Zou, H. Identification of seven novel virulence genes from Xanthomonas citri subsp. citri by Tn5-based random mutagenesis. J. Microbiol. 2015, 53, 330–336. [Google Scholar] [CrossRef]
- Kevany, B.M.; Rasko, D.A.; Thomas, M.G. Characterization of the complete zwittermicin a biosynthesis gene cluster from Bacillus cereus. Appl. Environ. Microb. 2008, 75, 1144–1155. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.; Caldovic, L.; Tuchman, M. Sources and fates of carbamyl phosphate: A labile energy-rich molecule with multiple facets. Biology 2018, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Song, X.; Zou, L.; Zou, H.; Chen, G. The small and large subunits of carbamoyl- phosphate synthase exhibit diverse contributions to pathogenicity in Xanthomonas citri subsp. citri. J. Integr. Agr. 2015, 14, 1338–1347. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, T.; Rou, W.; Song, X.; Guo, J.; Fan, X.J.; Kamau, G.G.; Zou, H. Molecular study on the carAB operon reveals that carB gene is required for swimming and biofilm formation in Xanthomonas citri subsp. citri. Bmc Microbiol. 2015, 15, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, B.A.; Bzik, D.J. De novo pyrimidine biosynthesis is required for virulence of Toxoplasma gondii. Nature 2002, 415, 926–929. [Google Scholar] [CrossRef]
- Garvis, S.; Munder, A.; Ball, G.; de Bentzmann, S.; Wiehlmann, L.; Ewbank, J.J.; Tümmler, B.; Filloux, A. Caenorhabditis elegans semi-automated liquid screen reveals a specialized role for the chemotaxis gene cheB2 in Pseudomonas aeruginosa virulence. PLoS Pathog. 2009, 5, e1000540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yorgey, P.; Rahme, L.G.; Tan, M.-W.; Ausubel, F.M. The roles of mucD and alginate in the virulence of Pseudomonas aeruginosa in plants, nematodes and mice. Mol. Microbiol. 2001, 41, 1063–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, C.; Nie, X.; Tang, Z.; Zhang, Y.; Lin, J.; Sun, M.; Peng, D. A novel serine protease, Sep1, from Bacillus firmus DS-1 has nematicidal activity and degrades multiple intestinal-associated nematode proteins. Sci. Rep. 2016, 6, 25012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, I.A.; Haas, D.; Heeb, S. Extracellular protease of Pseudomonas fluorescens CHA0, a biocontrol factor with activity against the root-knot nematode Meloidogyne incognita. Appl. Environ. Microb. 2005, 71, 5646–5649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landy, M.; Warren, G.H.; RosenmanM, S.B.; Colio, L.G. Bacillomycin: An antibiotic from Bacillus subtilis active against pathogenic fungi. Exp. Biol. Med. 1948, 67, 539–541. [Google Scholar] [CrossRef]
- Gray, N.F. Ecology of nematophagous fungi: Comparison of the soil sprinkling method with the Baermann funnel technique in the isolation of endoparasites. Soil Biol. Biochem. 1984, 16, 81–83. [Google Scholar] [CrossRef]
- Terefe, M.; Tefera, T.; Sakhuja, P.K. Effect of a formulation of Bacillus firmus on root-knot nematode Meloidogyne incognita infestation and the growth of tomato plants in the greenhouse and nursery. J. Invertebr. Pathol. 2009, 100, 94–99. [Google Scholar] [CrossRef]
- Sambrook, J.; Russel, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Plainview, TX, USA, 2001; pp. 116–126. [Google Scholar]
- Liu, Y.; Huang, N. Efficient amplification of insert end sequences from bacterial artificial chromosome clones by thermal asymmetric interlaced PCR. Plant. Mol. Biol. Rep. 1998, 16, 175. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | No. of Juveniles Hatched | Hatching Inhibition Rate (%) |
|---|---|---|
| 100% | 28.67 ± 7.37 a | 98.83 |
| 50% | 133.33 ± 18.61 b | 94.56 |
| 25% | 305.67 ± 34.39 c | 87.53 |
| Control | 2453 ± 121.20 d |
| Material | Relevant Genotype or Characteristics * | Source or Reference |
|---|---|---|
| Strains | ||
| Escherichia coli | ||
| DH5α | F-, φ80d/lacZΔM15, Δ(lacZYA-argF)U169,deoR | This lab |
| BL21(DE3) | F-, ompT, hsdSB(rB- mB-), gal dcm(DE3) | This lab |
| MB12 | Mutant of BL21(DE3), carB::Tn5; Kmr | This study |
| MB12-1 | Complementation of MB12 mutant with expression vector pColdcarB; Apr, Kmr | This study |
| Plasmids | ||
| pUC18 | E. coli clone vector; lacZ; Apr | This lab |
| pColdII | cspA promoter-based expression vector; Apr | This lab |
| pUCcarB | pUC18 derivative carrying carB; Apr | This study |
| pColdcarB | pCold II derivative carrying carB; cspA promoter-based expression vector; Apr | This study |
| Primers | Sequence (5′-3′; Restriction Sites Underlined if Present) | Use of Primers |
|---|---|---|
| carB-F | GGATCCATGCCAAAACGTACAGATAT (BamHI) | carB gene amplification |
| carB-R | AAGCTTTTATTTGATCTGTGCGTGCA (HindIII) | |
| pColdII-F | ACGCCATATCGCCGAAAGG | carB gene identification |
| pColdII-R | GGCAGGGATCTTAGATTCTG | |
| SP1 | GTCTTCGGTTTCCGTGTTTCG | Tn5 transposon left flanking sequences Pre-amplification |
| DP1 | NGTCGASWGANAWGAA | |
| SP2 | AAATGGCATCCGGATCTGCATC | Tn5 transposon left flanking sequences Primary amplification |
| DP2 | AGWGNAGWANCAWAGG | |
| SP3 | TACCCTGTGGAACACCTACATCTG | Tn5 transposon left flanking sequences Second amplification |
| DP3 | CAWCGICNGAIASGAA | |
| SP4 | GGTTGTAACACTGGCAGAGCATT | Tn5 transposon right flanking sequences Pre-amplification |
| DP4 | TCSTICGNACITWGGA | |
| SP5 | CGCATCTTCCCGACAACGCAG | Tn5 transposon right flanking sequences Primary amplification |
| DP5 | STTGNTASTNCTNTGC | |
| SP6 | AACTGGTCCACCTACAACAAAG | Tn5 transposon right flanking sequences Second amplification |
| DP6 | WCAGNTGWTNGTNCTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, Y.; Li, S.; Xu, G.; Xie, S.; Liu, X.; Lin, X.; Wu, H.; Gao, X. The carB Gene of Escherichia coli BL21(DE3) is Associated with Nematicidal Activity against the Root-Knot Nematode Meloidogyne javanica. Pathogens 2021, 10, 222. https://doi.org/10.3390/pathogens10020222
Xia Y, Li S, Xu G, Xie S, Liu X, Lin X, Wu H, Gao X. The carB Gene of Escherichia coli BL21(DE3) is Associated with Nematicidal Activity against the Root-Knot Nematode Meloidogyne javanica. Pathogens. 2021; 10(2):222. https://doi.org/10.3390/pathogens10020222
Chicago/Turabian StyleXia, Yanfei, Shen Li, Guohui Xu, Shanshan Xie, Xueting Liu, Xiaomin Lin, Huijun Wu, and Xuewen Gao. 2021. "The carB Gene of Escherichia coli BL21(DE3) is Associated with Nematicidal Activity against the Root-Knot Nematode Meloidogyne javanica" Pathogens 10, no. 2: 222. https://doi.org/10.3390/pathogens10020222