
Antimicrobial and Anti-Biofilm Activity of Polymyxin E Alone and in Combination with Probiotic Strains of Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895 against Clinical Isolates of Selected Acinetobacter spp.: A Preliminary Study

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Bacterial Isolation and Identification
2.2. Bacterial Identification and Antibiotic Susceptibility Using VITEK 2 System
2.3. Probiotic Strains Were Tolerant to Polymyxin E
2.4. Minimum Inhibitory Concentrations (MICs)
2.5. The Bacilli Strains Auto-Aggregated and Co-Aggregated with Isolated Pathogens
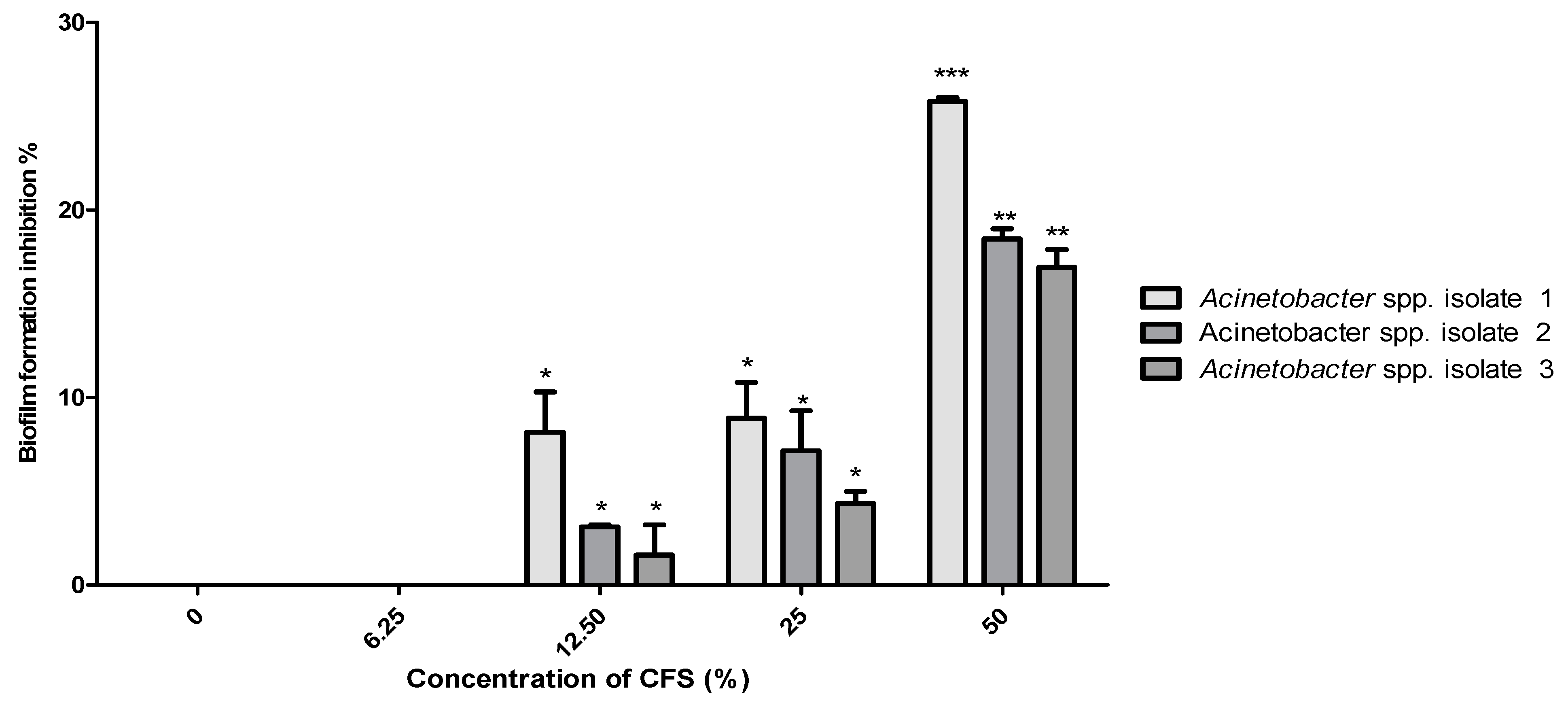
2.6. Minimum Biofilm Inhibitory Concentrations (MBIC)
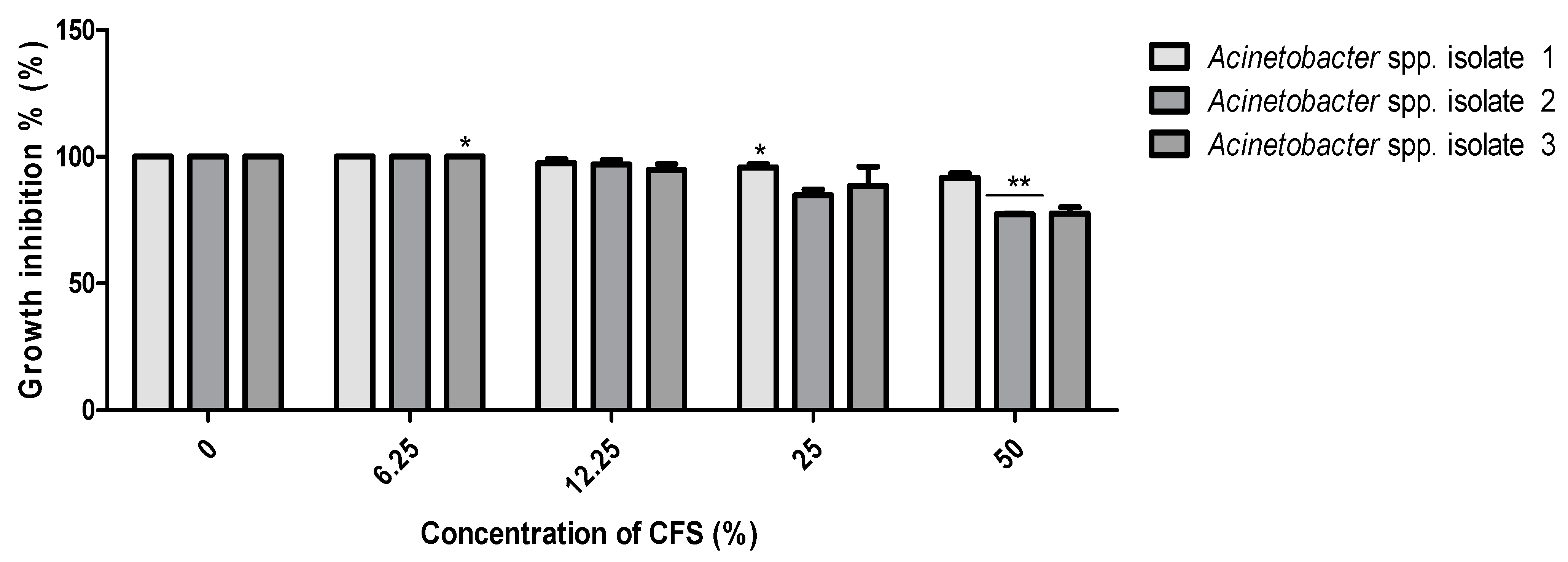
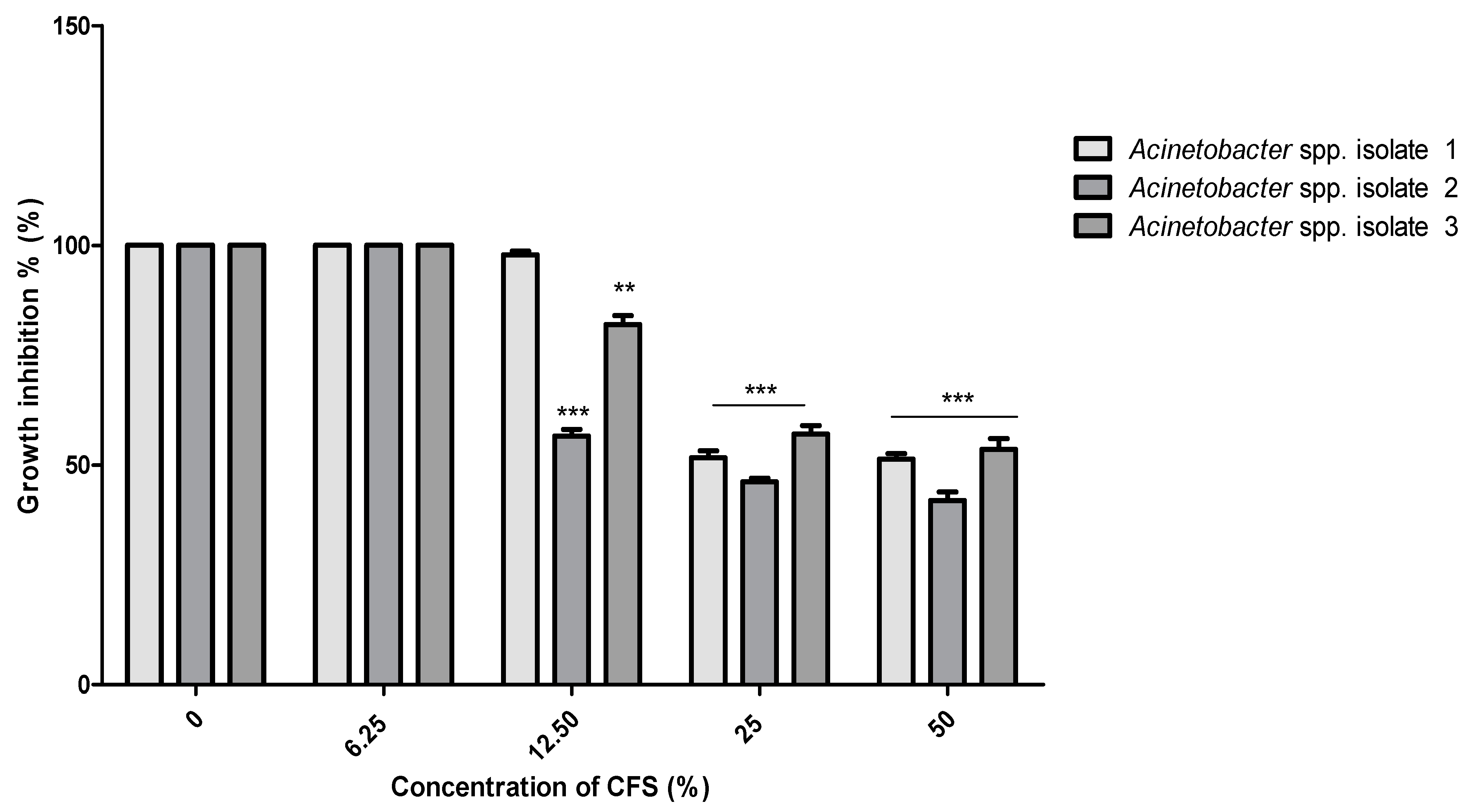
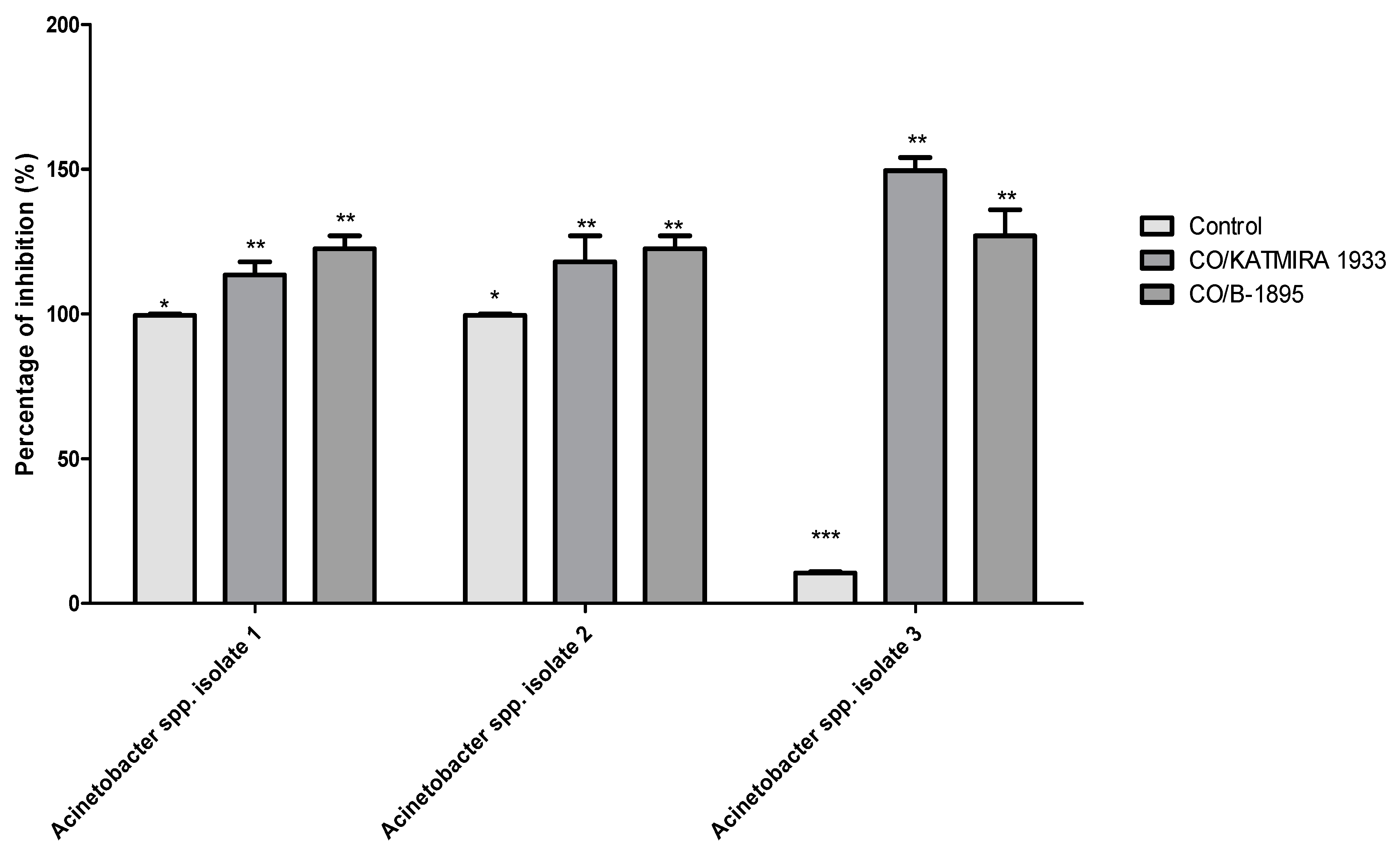
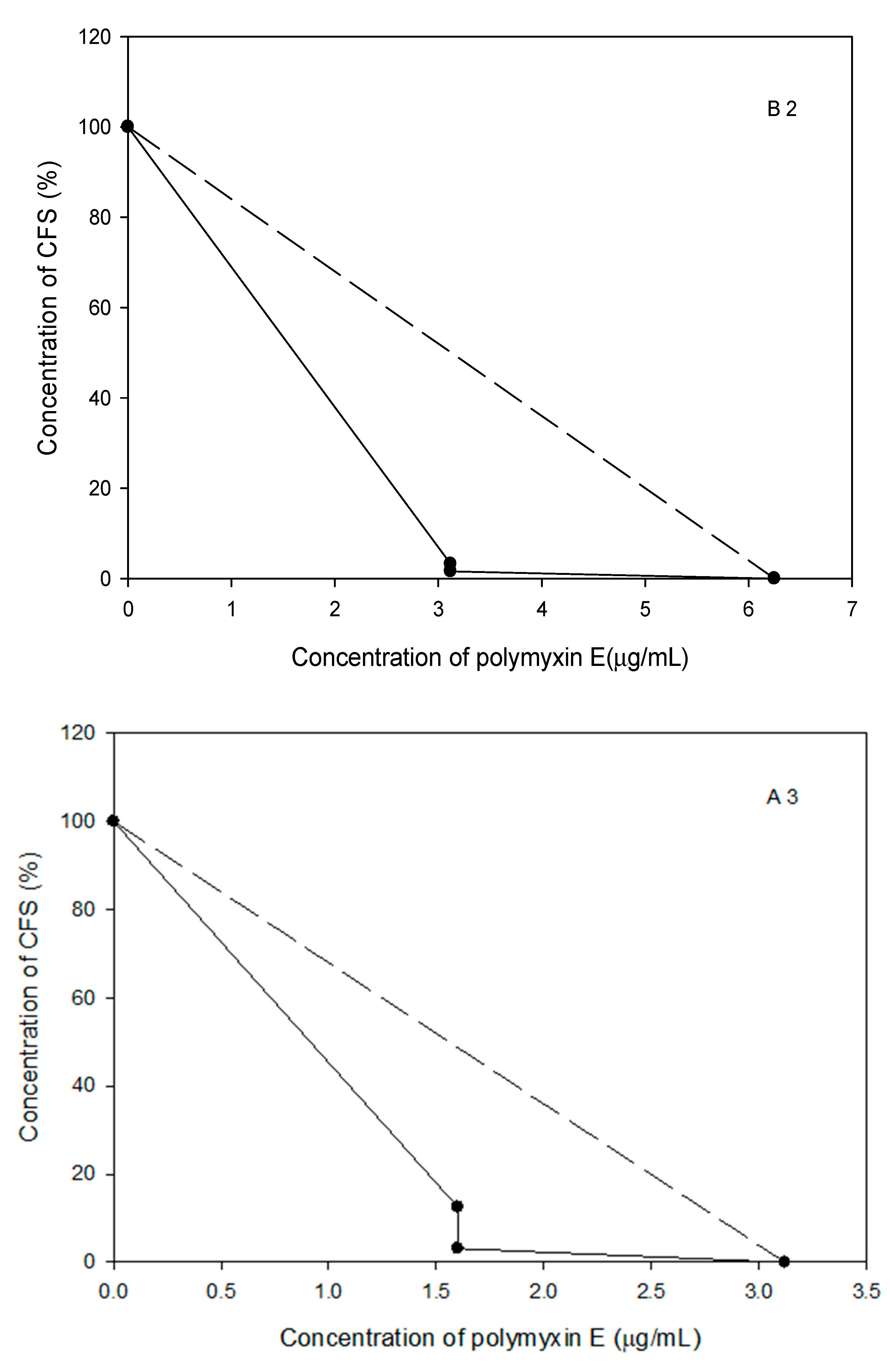
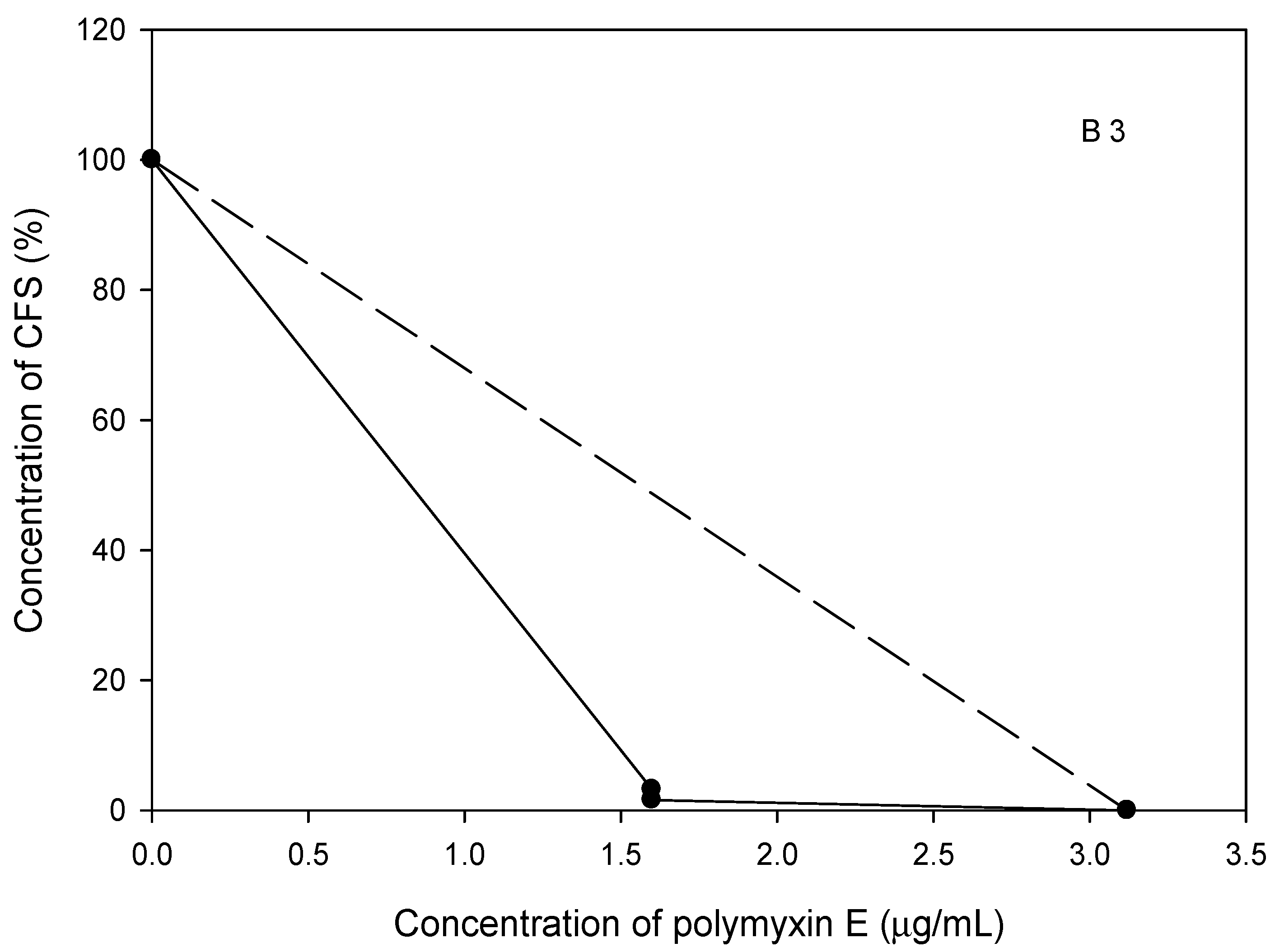
2.7. The Polymyxin E Synergizes with Bacilli CFS
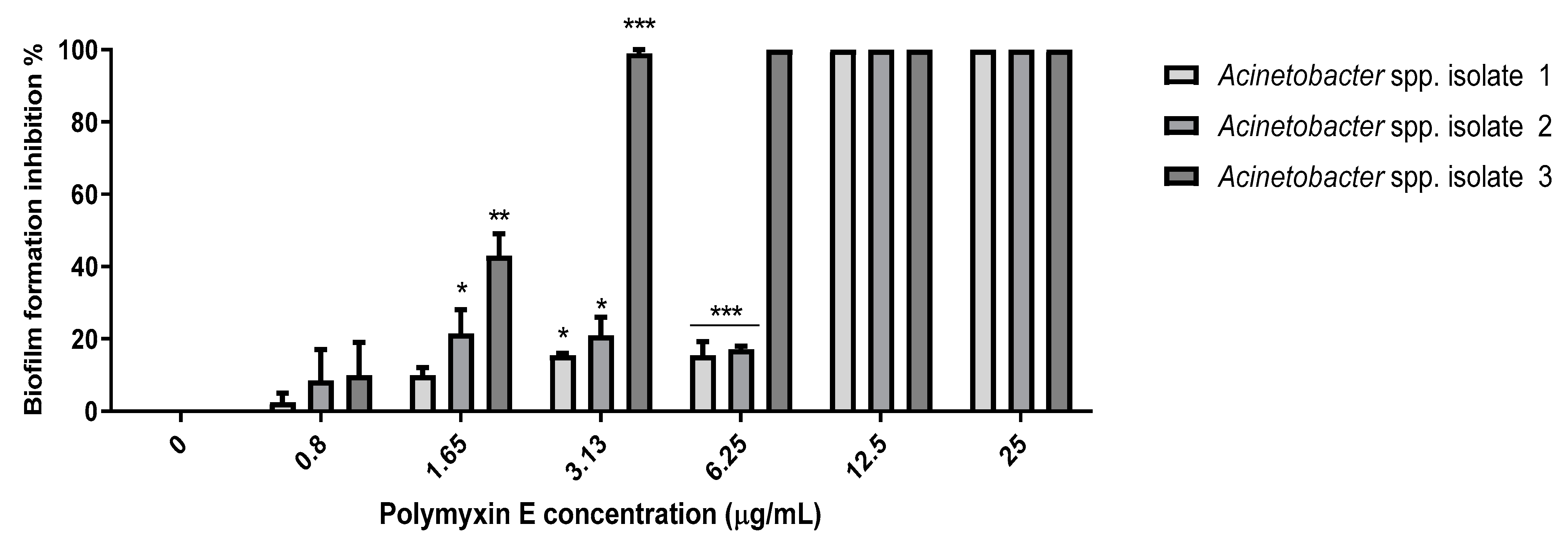
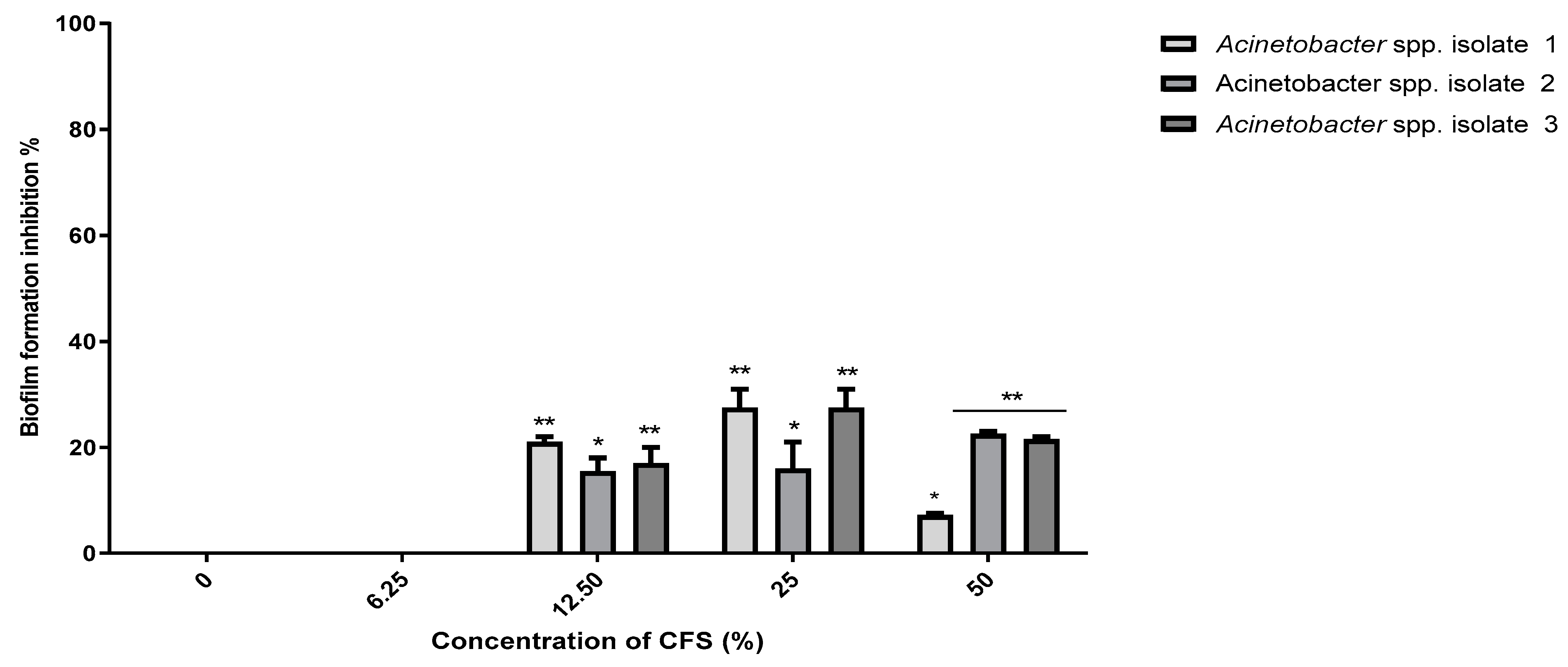
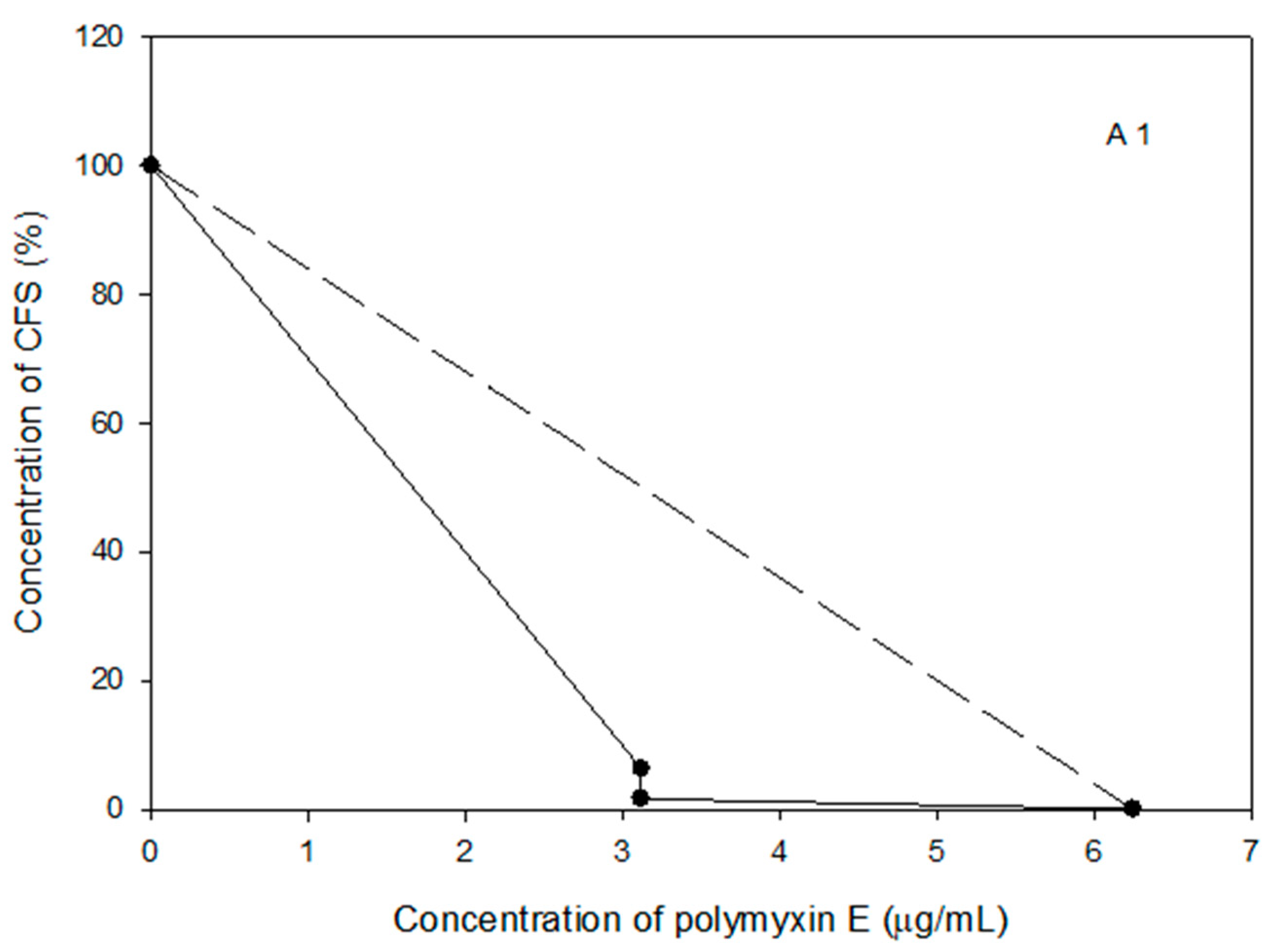
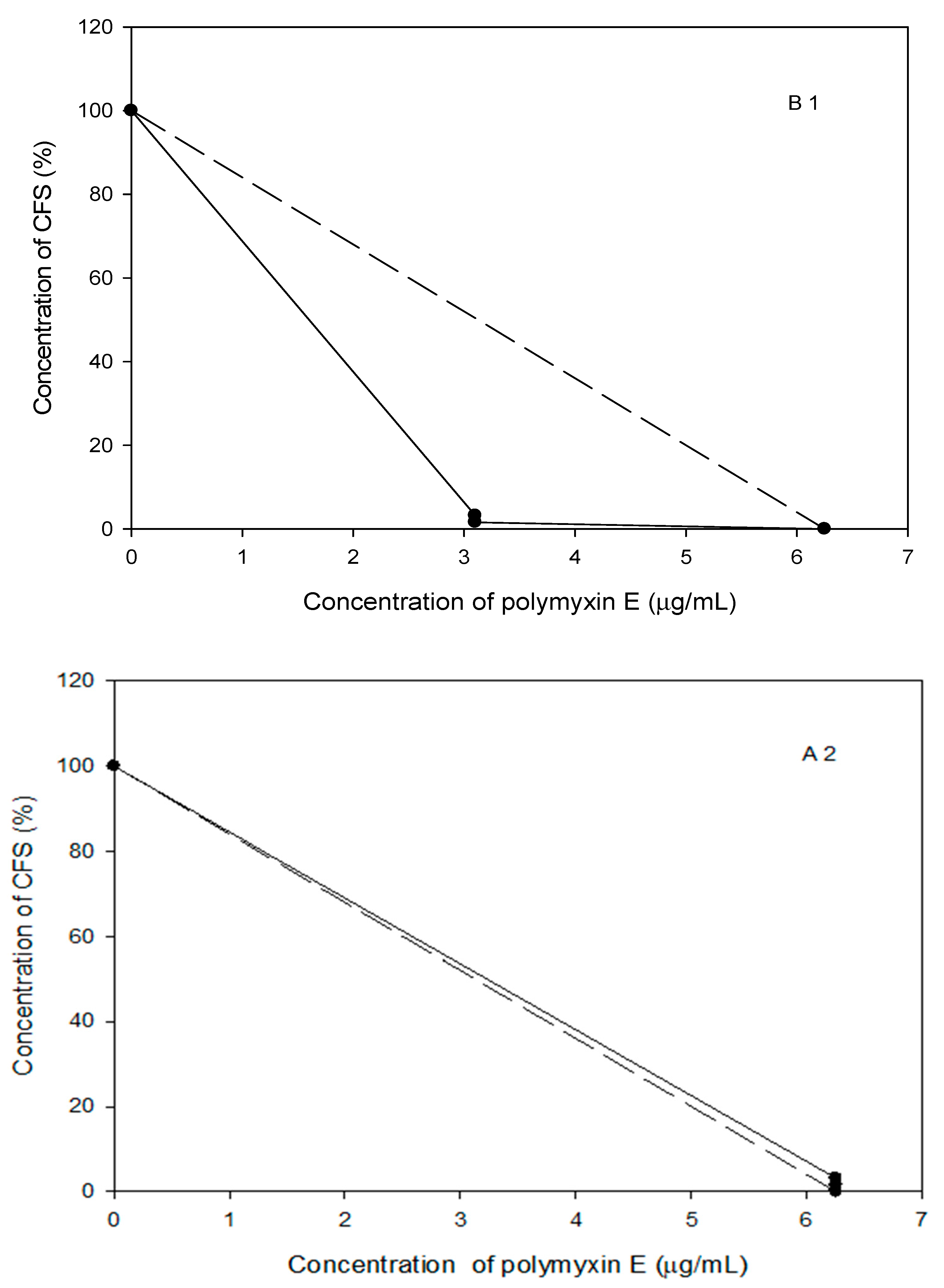
2.8. Probiotic Strains Synergized with Polymyxin E against the Planktonic and Biofilm Cells of the Selected Acinetobacter spp. Isolates
3. Discussion
4. Materials and Methods
4.1. Bacterial Growth Conditions, Isolation, and Identification of Acinetobacter spp. Isolates
4.2. Ethical Statement and Consent
4.3. Antibiogram Assay of the Selected Acinetobacter spp. Isolates and the Probiotic Bacillus Strains
4.4. Biofilm Formation Assay
4.5. Preparation of CFS of the Tested Probiotics
4.6. Antibiotics Combination with CFS of Probiotics Using Disc Diffusion Method
4.7. Coaggregation Test
4.8. Minimum Inhibitory Concentration (MIC)
4.9. Determination of Minimal Biofilm Inhibitory Concentration (MBIC)
4.10. Checkerboard Assay for Antimicrobial Combinations
4.11. Checkerboard Assay, Data Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martín-Aspas, A.; Guerrero-Sánchez, F.M.; García-Colchero, F.; Rodríguez-Roca, S.; Girón-González, J.-A. Differential characteristics of Acinetobacter baumannii colonization and infection: Risk factors, clinical picture, and mortality. Infect. Drug Resist. 2018, 11, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Morris, F.; Dexter, C.; Kostoulias, X.; Uddin, M.I.; Peleg, A.Y. The Mechanisms of Disease Caused by Acinetobacter baumannii. Front. Microbiol. 2019, 10, 1601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, D.-S.; Eom, Y.-B. Antimicrobial and anti-biofilm activities of Clostridium butyricum supernatant against Acinetobacter baumannii. Arch. Microbiol. 2020, 202, 1059–1068. [Google Scholar] [CrossRef]
- Rezaee, P.; Hamzeh, A.; Mohammadi, M. Acinetobacter baumannii antibiotics resistance in Iran. J. Bacteriol. Mycol. Open Access 2019, 7, 159–162. [Google Scholar] [CrossRef]
- Raheem, A.; Hussain, S.S.; Essa, R.H.; Atia, A.-J. The effect of new hydantoin derivative (compound) on Acinetobacter baumannii biofilm formation isolated from clinical sources. J. Univ. Babylon Pure App. Sci. 2018, 26, 71–79. [Google Scholar]
- Asif, M.; Alvi, I.A.; Rehman, S.U. Insight into Acinetobacter baumannii: Pathogenesis, global resistance, mechanisms of resistance, treatment options, and alternative modalities. Infect. Drug Resist. 2018, 11, 1249–1260. [Google Scholar] [CrossRef] [Green Version]
- Pacheco, T.; Bustos, R.-H.; González, D.; Garzón, V.; García, J.-C.; Ramírez, D. An Approach to Measuring Colistin Plasma Levels Regarding the Treatment of Multidrug-Resistant Bacterial Infection. Antibiotics 2019, 8, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farshadzadeh, Z.; Taheri, B.; Rahimi, S.; Shoja, S.; Pourhajibagher, M.; Haghighi, M.A.; Bahador, A. Growth Rate and Biofilm Formation Ability of Clinical and Laboratory-Evolved Colistin-Resistant Strains of Acinetobacter baumannii. Front. Microbiol. 2018, 9, 153. [Google Scholar] [CrossRef]
- Mosca, A.; Dalfino, L.; Romanelli, F.; Stolfa, S.; Prete, R.D.; Santacroce, L. Effectiveness of colistin with rifampicin and meropenem against colistin-resistant Acinetobacter baumannii strains: An in vitro study. Pharmacophore 2020, 11, 1–6. [Google Scholar]
- Isayenko, O.Y.; Knysh, O.V.; Kotsar, O.V.; Ryzhkova, T.N.; Dyukareva, G.I. Simultaneous and sequential influence of metabolite complexes of Lactobacillus rhamnosus and Saccharomyces boulardii and antibiotics against poly-resistant Gram-negative bacteria. Regul. Mech. Biosyst. 2020, 11, 139–145. [Google Scholar] [CrossRef]
- Food and Agriculture Organization/World Health Organization Expert Consultation (FAO/WHO). Evaluation of Health and Nutritional Properties of Probiotics in Food including Powder Milk with Live Lactic Acid Bacteria; [cited 2005 September 8]; Food and Agriculture Organization of the United Nations and World Health Organization: Córdoba, Argentina, 2001; Available online: https://www.fao.org/3/a0512e/a0512e.pdf (accessed on 1 October 2021).
- Hager, C.L.; Isham, N.; Schrom, K.P.; Chandra, J.; McCormick, T.; Miyagi, M.; Ghannoum, M.A. Effects of a Novel Probiotic Combination on Pathogenic Bacterial-Fungal Polymicrobial Biofilms. mBio 2019, 10, e00338-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karacaer, F.; Hamed, I.; Özogul, F.; Glew, R.H.; Özcengiz, D. The function of probiotics on the treatment of ventilator-associated pneumonia (VAP): Facts and gaps. J. Med. Microbiol. 2017, 66, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, L. Acinetobacter baumannii biofilm resistance mechanisms and prevention and control of progress. New Century Sci. Press. 2016, 3, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Hadid, M.A. Evaluating the Effects of Probiotics and Vinegars on Bacteria Isolated from Burns and Wounds Infections. Master’s Thesis, College of Science, Al-Nahrain University, Baghdad, Iraq, 2015. [Google Scholar]
- AL-Dahlaki, S.M.M. Molecular Detection and Gene Expression for Hcpandblaoxa-51 genes in Acinetobacter baumannii Isolated from Different Clinical Sources. Master’s Thesis, College of Science, University of Diyala, Baghdad, Iraq, 2020. [Google Scholar]
- Raut, S.; Rijal, K.R.; Khatiwada, S.; Karna, S.; Khanal, R.; Adhikari, J.; Adhikari, B. Trend and characteristics of Acinetobacter baumannii infections in patients attending universal college of medical sciences, Bhairahawa, Western Nepal: A Longitudinal Study of 2018. Infect. Drug Resist. 2020, 13, 1631–1641. [Google Scholar] [CrossRef]
- Pal, N.; Sujatha, R.; Kumar, A. Phenotypic and genotypic identification of Acinetobacter baumannii with special reference to blaoxa-51 Like gene and its antimicrobial susceptibility pattern from intensive care uniteS in Kanpur. Int. J. Contemp. Med. Res. 2017, 4, 1154–1158. [Google Scholar]
- Rahimi, S.; Farshadzadeh, Z.; Taheri, B.; Mohammadi, M.; Haghighi, M.-A.; Bahador, A. The Relationship Between Antibiotic Resistance Phenotypes and Biofilm Formation Capacity in Clinical Isolates of Acinetobacter baumannii. Jundishapur J. Microbiol. 2018, 11, e74315. [Google Scholar] [CrossRef] [Green Version]
- Nhu, N.T.K.; Riordan, D.W.; Nhu, T.D.H.; Thanh, D.P.; Thwaites, G.; Lan, N.P.H.; Wren, B.W.; Baker, S.; A Stabler, R. The induction and identification of novel Colistin resistance mutations in Acinetobacter baumannii and their implications. Sci. Rep. 2016, 6, 28291. [Google Scholar] [CrossRef]
- Maleki, M.H.; Sekawi, Z.; Soroush, S.; Azizi-Jalilian, F.; Asadollahi, K.; Mohammadi, S.; Emaneini, M.; Taherikalani, M. Phenotypic and genotypic characteristics of tetracycline resistant Acinetobacter baumannii isolates from nosocomial infections at Tehran hospitals. Iranian J. Basic Med. Sci. 2014, 17, 21–26. [Google Scholar] [CrossRef]
- Almasaudi, S.B. Acinetobacter spp. as nosocomial pathogens: Epidemiology and resistance features. Saudi J. Biol. Sci. 2016, 25, 586–596. [Google Scholar] [CrossRef] [Green Version]
- Taitt, C.R.; Leski, T.; Stockelman, M.G.; Craft, D.W.; Zurawski, D.V.; Kirkup, B.; Vora, G.J. Antimicrobial Resistance Determinants in Acinetobacter baumannii Isolates Taken from Military Treatment Facilities. Antimicrob. Agents Chemother. 2013, 58, 767–781. [Google Scholar] [CrossRef] [Green Version]
- Ugolotti, E.; Di Marco, E.; Bandettini, R.; Tripodi, G.; Biassoni, R. The whole genome sequencing of Acinetobacter-calcoaceticus-baumannii complex strains involved in suspected outbreak in an intensive care unit of a pediatric hospital. J. Hosp. Adm. 2016, 5, 81. [Google Scholar] [CrossRef] [Green Version]
- Moubareck, C.; Halat, D. Insights into Acinetobacterbaumannii: A review of microbiological, virulence, and resistance traits in a threatening nosocomial pathogen. Antibiotics 2020, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug resistant, extensively drug resistant, and pan drug resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2011, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef]
- Bahador, A.; Raoo An, R.; Farshadzadeh, Z.; Beitollahi, L.; Khaledi, A.; Rahimi, S.; Mokhtaran, M.; Tavana, A.M.; Esmaeili, D. The prevalence of IS Aba 1 and IS Aba 4 in Acinetobacter baumannii species of different international clone lineages among patients with burning in Tehran, Iran. Jundishapur J. Microbiol. 2015, 8, e17167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Runci, F.; Bonchi, C.; Frangipani, E.; Visaggio, D.; Visca, P. Acinetobacter baumannii Biofilm Formation in Human Serum and Disruption by Gallium. Antimicrob. Agents Chemother. 2017, 61, e01563-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eze, E.C.; Chenia, H.Y.; El Zowalaty, M.E. Acinetobacter baumannii biofilms: Effects of physicochemical factors, virulence, antibiotic resistance determinants, gene regulation, and future antimicrobial treatments. Infect. Drug Resist. 2018, 11, 2277–2299. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Unno, Y.; Ubagai, T.; Ono, Y. Sub-minimum inhibitory concentrations of colistin and polymyxin B promote Acinetobacter baumannii biofilm formation. PLoS ONE 2018, 13, e0194556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.-F.; Lin, Y.-Y.; Lan, C.-Y. Characterization of biofilm production in different strains of Acinetobacter baumannii and the effects of chemical compounds on biofilm formation. PeerJ 2020, 8, e9020. [Google Scholar] [CrossRef]
- Efremenkova, O.; Gabrielyan, N.; Malanicheva, I.; Demiankova, M.; Efimenko, T.; Rogozhin, E.; Sharapchenko, S.; Krupenio, T.; Davydov, D.; Kornilov, M. Antimicrobial properties of the probiotic strain Bacillus subtilis 534. Int. Arch. Med. Microbiol. 2019, 2, 119. [Google Scholar]
- Soltan, M.M.; Davoodabadi, A.; Abdi, M.; Hajiabdolbaghi, M.; SharifiYazdi, M.K.; Douraghi, M.; TabatabaeiBafghi, S.M. Inhibitory effect of Lactobacillus plantarum and Lb. fermentum isolated from the faeces of healthy infants against non fermentative bacteria causing nosocomial infections. New Microb. New Infect. 2017, 15, 9–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lashin, G.M.A.; Tohamy, E.Y.; Askora, A.A.; El-Zahraa Mahmoud, F. Use of probiotic acid bacteria for the control of multidrug resistant bacterial isolated from clinical infections. Bull. Fac. Sci. Zagazig Univ. 2017, 39, 61–81. [Google Scholar] [CrossRef]
- Kumariya, R.; Garsa, A.K.; Rajput, Y.; Sood, S.; Akhtar, N.; Patel, S. Bacteriocins: Classification, synthesis, mechanism of action and resistance development in food spoilage causing bacteria. Microb. Pathog. 2019, 128, 171–177. [Google Scholar] [CrossRef] [PubMed]
- AlGburi, A.; Alazzawi, S.A.; Al-Ezzy, A.I.A.; Weeks, R.; Chistyakov, V.; Chikindas, M.L. Potential Probiotics Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895 Co-Aggregate with Clinical Isolates of Proteus mirabilis and Prevent Biofilm Formation. Probiotics Antimicrob. Proteins 2020, 12, 1471–1483. [Google Scholar] [CrossRef] [PubMed]
- Zamani, H.; Rahbar, S.; Garakoui, S.R.; Sahebi, A.A.; Jafari, H.; Microbiology, S.O.P.C.S.O. Antibiofilm potential of Lactobacillus plantarum spp. cell free supernatant (CFS) against multidrug resistant bacterial pathogens. Pharm. Biomed. Res. 2017, 3, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Chapot-Chartier, M.P.; Kulakauskas, S. Cell wall structure and function in lactic acid bacteria. Microb. Cell Factories 2014, 13, S9. [Google Scholar] [CrossRef] [Green Version]
- Algburi, A.; Al-Hasani, H.M.; Ismael, T.K.; Abdelhameed, A.; Weeks, R.; Ermakov, A.M.; Chikindas, M.L. Antimicrobial Activity of Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895 Against Staphylococcus aureus Biofilms Isolated from Wound Infection. Probiotics Antimicrob. Proteins 2020, 13, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Mathur, H.; Field, D.; Rea, M.C.; Cotter, P.; Hill, C.; Ross, R.P. Fighting biofilms with lantibiotics and other groups of bacteriocins. Npj Biofilms Microbiomes 2018, 4, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, M.S.M.; Lourenço, M.L.M.C.; Vasconcelos, B.M.; Carneiro, V.A. Probiotics Lactobacillus strains: A promising alternative therapy against to biofilm-forming enteropathogenicbacteria? Afr. J. Microbiol. Res. 2019, 13, 544–551. [Google Scholar]
- Isayenko, O.Y. Synergistic activity of filtrates of Lactobacillus rhamnosus and Saccharomyces boulardii and antibacterial preparations against Corynebacterium spp. Regul. Mech. Biosyst. 2019, 10, 445–456. [Google Scholar] [CrossRef]
- Cavera, V.L.; Volski, A.; Chikindas, M.L. The Natural Antimicrobial Subtilosin A Synergizes with Lauramide Arginine Ethyl Ester (LAE), ε-Poly-l-lysine (Polylysine), Clindamycin Phosphate and Metronidazole, Against the Vaginal Pathogen Gardnerella vaginalis. Probiotics Antimicrob. Proteins 2015, 7, 164–171. [Google Scholar] [CrossRef]
- Al-Gburi, A.; Volski, A.; Chikindas, M.L. Natural antimicrobials subtilosin and lauramide arginine ethyl ester synergize with conventional antibiotics clindamycin and metronidazole against biofilms of Gardnerella vaginalis but not against biofilms of healthy vaginal lactobacilli. Pathog. Dis. 2015, 73, ftv018. [Google Scholar] [CrossRef]
- Karlyshev, A.V.; Melnikov, V.G.; Chistyakov, V.A.; Masood, N.; Jackson, E.; Moore, K.; Farbos, A.; Paszkiewicz, K.; Dickins, B.; McNally, A.; et al. Draft Genome Sequence of Bacillus amyloliquefaciens B-1895. Genome Announc. 2014, 2, e00585-14. [Google Scholar] [CrossRef] [Green Version]
- Rowles, H.L. How are Probiotics Affected by Antibiotics? Ann. Clin. Lab. Res. 2017, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Cappuccino, J.G.; Welsh, C.T. Microbiology: A Laboratory Manual, 11th ed.; Pearson Education: London, UK, 2018. [Google Scholar]
- Lotfi, G.; Hassaine, H.; Klouche, N.; Khadir, A.; Aissaoui, N.; Nas, F.; Zingg, W. Detection of biofilm formation of a collection of fifty strains of Staphylococcus aureus isolated in Algeria at the University Hospital of Tlemcen. J. Bacteriol. Res. 2014, 6, 1–6. [Google Scholar] [CrossRef]
- Tang, J.; Kang, M.; Chen, H.; Shi, X.; Zhou, R.; Chen, J.; Du, Y. The staphylococcal nuclease prevents biofilm formation in Staphylococcus aureus and other biofilm-forming bacteria. Sci. China Life Sci. 2011, 54, 863–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Gburi, A.; Volski, A.; Cugini, C.; Walsh, E.M.; Chistyakov, V.A.; Mazanko, M.S.; Bren, A.B.; Dicks, L.M.T.; Chikindas, M.L. Safety Properties and Probiotic Potential of Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895. Adv. Microbiol. 2016, 6, 432–452. [Google Scholar] [CrossRef] [Green Version]
- Ledder, R.G.; Timperley, A.S.; Friswell, M.K.; Macfarlane, S.; McBain, A.J. Coaggregation between and among human intestinal and oral bacteria. FEMS Microbiol. Ecol. 2008, 66, 630–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Algburi, A.; Zhang, Y.; Weeks, R.; Comito, N.; Zehm, S.; Pinto, J.; Uhrich, K.E.; Chikindas, M.L. Gemini Cationic Amphiphiles Control Biofilm Formation by Bacterial Vaginosis Pathogens. Antimicrob. Agents Chemother. 2017, 61, e00650-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turovskiy, Y.; Chikindas, M.L. Zinc Lactate and Sapindin Act Synergistically with Lactocin 160 Against Gardnerella vaginalis. Probiotics Antimicrob. Proteins 2011, 3, 144–149. [Google Scholar] [CrossRef] [Green Version]
- European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Diseases (ESCMID). Terminology relating to methods for the determination of susceptibility of bacteria to antimicrobial agents. Clin. Microbiol. Infect. 2000, 6, 503–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics Family | Antibiotics Type | Resistant Isolates No. and % |
|---|---|---|
| Penicillins | Ampicillin | Acinetobacter spp. isolates 1, 2, and 3 |
| B-lactam inhibitors | Piperacillin/Tazobactam | Acinetobacter spp. isolates 1, 2, and 3 |
| Cephalosporins | Cefazolin | Acinetobacter spp. isolates 1, 2, and 3 |
| Cefoxitin | Acinetobacter spp. isolates 1, 2, and 3 | |
| Ceftazidime | Acinetobacter spp. isolates 1, 2, and 3 | |
| Ceftriaxone | Acinetobacter spp. isolates 1, 2, and 3 | |
| Cefepime | Acinetobacter spp. isolates 1, 2, and 3 | |
| Carbapenems | Imipenem | Acinetobacter spp. isolates 1, 2, and 3 |
| Meropenem | Acinetobacter spp. isolates 1, 2, and 3 | |
| Aminoglycosides | Amikacin | Acinetobacter spp. isolates 1 and 2 |
| Gentamicin | Acinetobacter spp. isolates 1 and 2 | |
| Tobramycin | Acinetobacter spp. isolates 1 and 2 | |
| Tetracyclines | Tigecycline | Acinetobacter spp. isolate 1 |
| Minocycline | Acinetobacter spp. isolate 1 | |
| Fluoroquinolones | Ciprofloxacin | Acinetobacter spp. isolates 1, 2, and 3 |
| Levofloxacin | Acinetobacter spp. isolates 1, 2, and 3 | |
| Folate pathway antagonists | Trimethoprim /sulfamethoxazole | Acinetobacter spp. isolates 1 and 2 |
| Lipopeptides | Polymyxin E | None |
| Probiotic Strains | The Average Diameters of Inhibition Zone around Antibiotic Discs (mm) | |||||
|---|---|---|---|---|---|---|
| AK * | PME * | FOX * | CTX * | MEM * | TS * | |
| Bacillus subtilis KATMIRA1933 | 22 | zero | 35 | 40 | 37 | 37 |
| Bacillus amyloliquefaciens B-1895 | 14 | zero | 28 | 28 | 35 | 30 |
| Bacterial Strains | Auto- and Co-Aggregation % after 4 h | Auto- and Co-Aggregation % after 24 h |
|---|---|---|
| B.amyloliquefaciens B-1895 | 25.5% | 95.7% |
| B. subtilis KATMIRA1933 | 22.76% | 82.4% |
| Acinetobacter spp. isolates1, 2, and 3 | (24.5%, 20.58%, 28.97%) | (0.48%, 8.4%, 30.5%) |
| B.amyloliquefaciens B-1895 + Acinetobacter spp. isolate 1 | 23.98% | 50.57%, |
| B.amyloliquefaciens B-1895 + Acinetobacter spp. isolate 2 | 29.39% | 55.64% |
| B.amyloliquefaciens B-1895 + Acinetobacter spp. isolate 3 | 17.15% | 50.67% |
| B. subtilis KATMIRA1933 + Acinetobacter spp. isolate 1 | 33.43% | 60.1%, |
| B. subtilis KATMIRA1933 + Acinetobacter spp. isolate 2 | 31.89% | 53.16% |
| B. subtilis KATMIRA1933 + Acinetobacter spp. isolate 3 | 34% | 62.8% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
AL-Dulaimi, M.; Algburi, A.; Abdelhameed, A.; Mazanko, M.S.; Rudoy, D.V.; Ermakov, A.M.; Chikindas, M.L. Antimicrobial and Anti-Biofilm Activity of Polymyxin E Alone and in Combination with Probiotic Strains of Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895 against Clinical Isolates of Selected Acinetobacter spp.: A Preliminary Study. Pathogens 2021, 10, 1574. https://doi.org/10.3390/pathogens10121574
AL-Dulaimi M, Algburi A, Abdelhameed A, Mazanko MS, Rudoy DV, Ermakov AM, Chikindas ML. Antimicrobial and Anti-Biofilm Activity of Polymyxin E Alone and in Combination with Probiotic Strains of Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895 against Clinical Isolates of Selected Acinetobacter spp.: A Preliminary Study. Pathogens. 2021; 10(12):1574. https://doi.org/10.3390/pathogens10121574
Chicago/Turabian StyleAL-Dulaimi, Munaf, Ammar Algburi, Alyaa Abdelhameed, Maria S. Mazanko, Dmitry V. Rudoy, Alexey M. Ermakov, and Michael L. Chikindas. 2021. "Antimicrobial and Anti-Biofilm Activity of Polymyxin E Alone and in Combination with Probiotic Strains of Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895 against Clinical Isolates of Selected Acinetobacter spp.: A Preliminary Study" Pathogens 10, no. 12: 1574. https://doi.org/10.3390/pathogens10121574