
Modulation of Production of Th1/Th2 Cytokines in Peripheral Blood Mononuclear Cells and Neutrophils by Hepatitis C Virus Infection in Chronically Infected Patients

Abstract
:1. Introduction
2. Results
2.1. Subjects
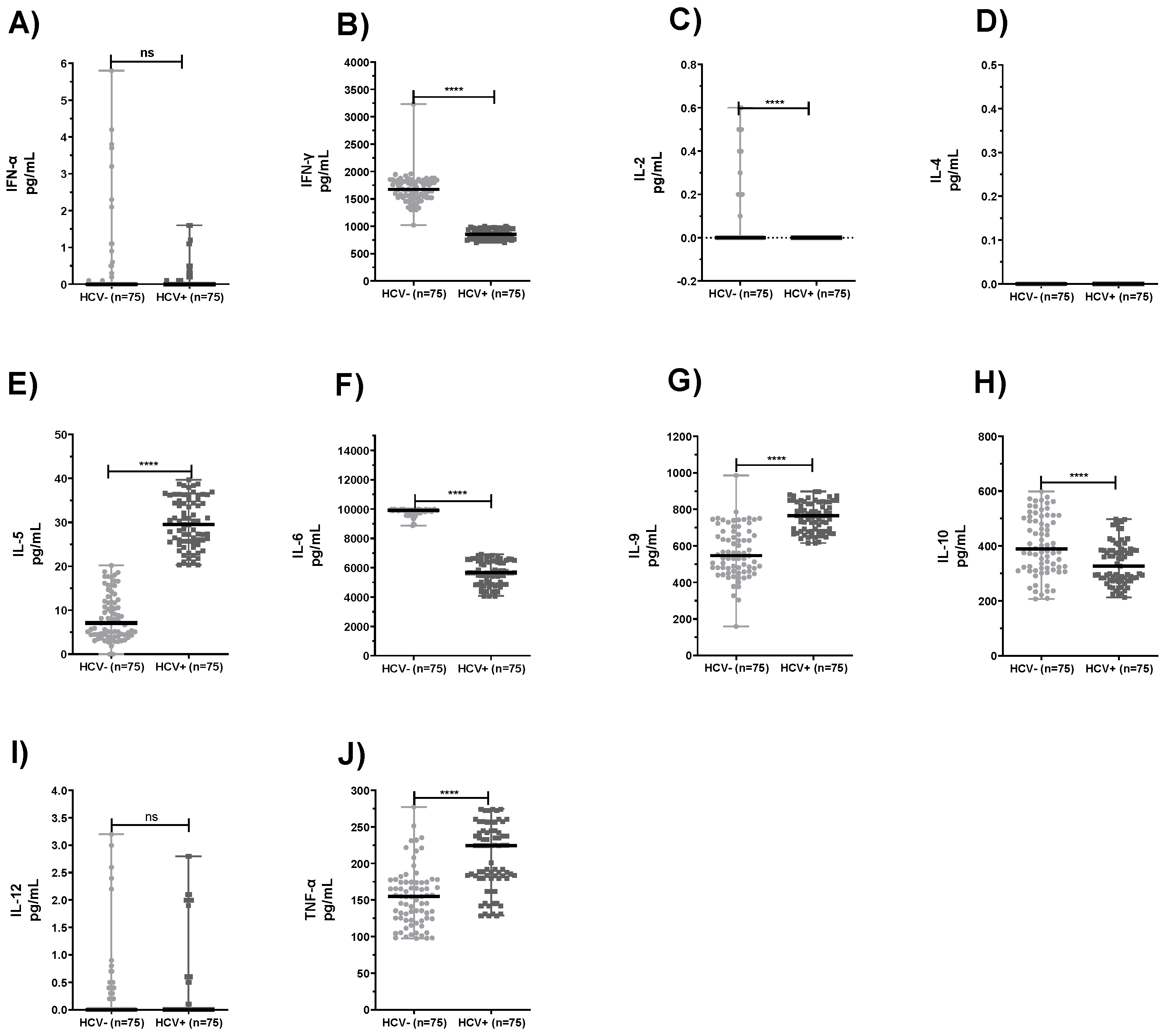
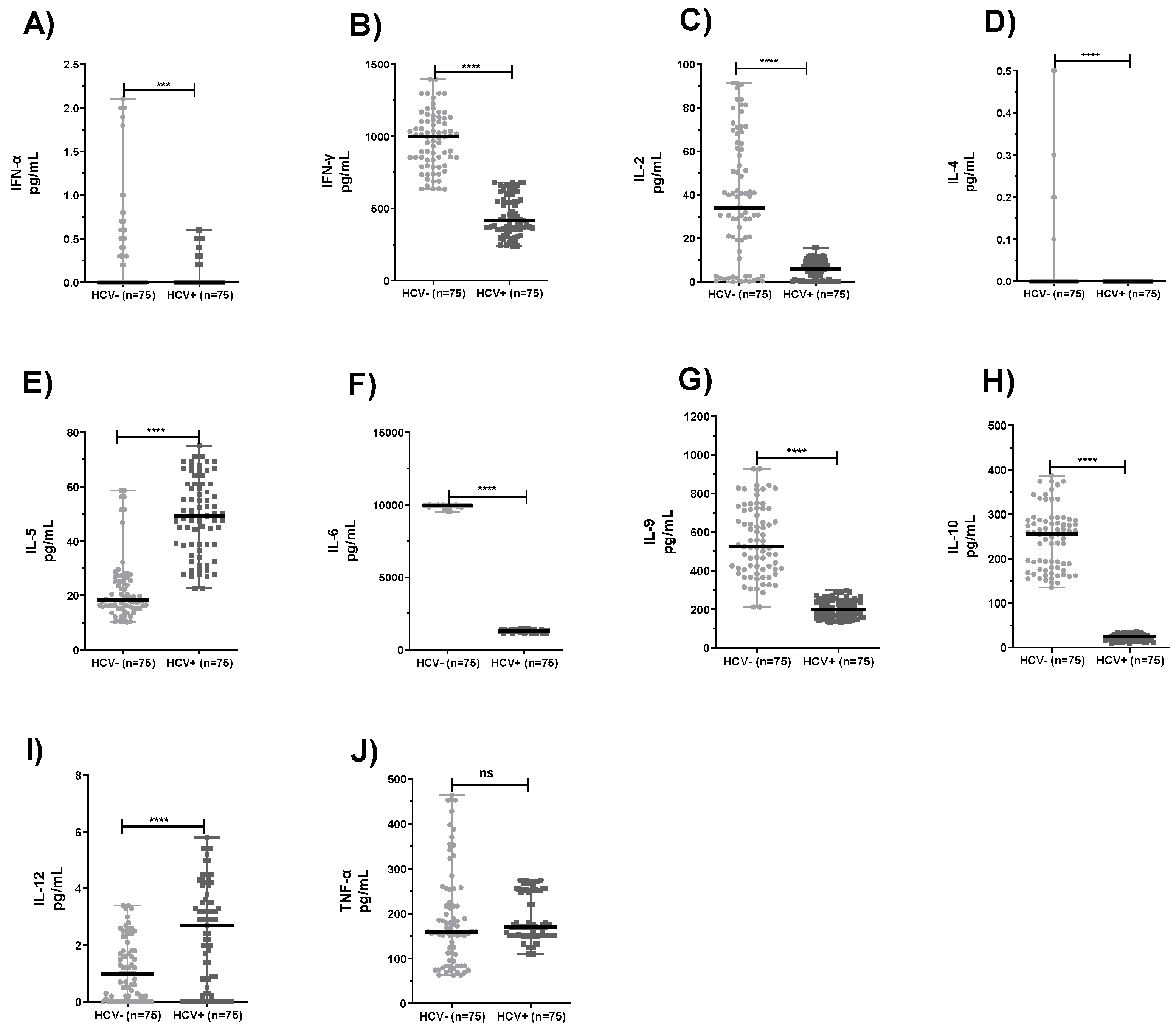
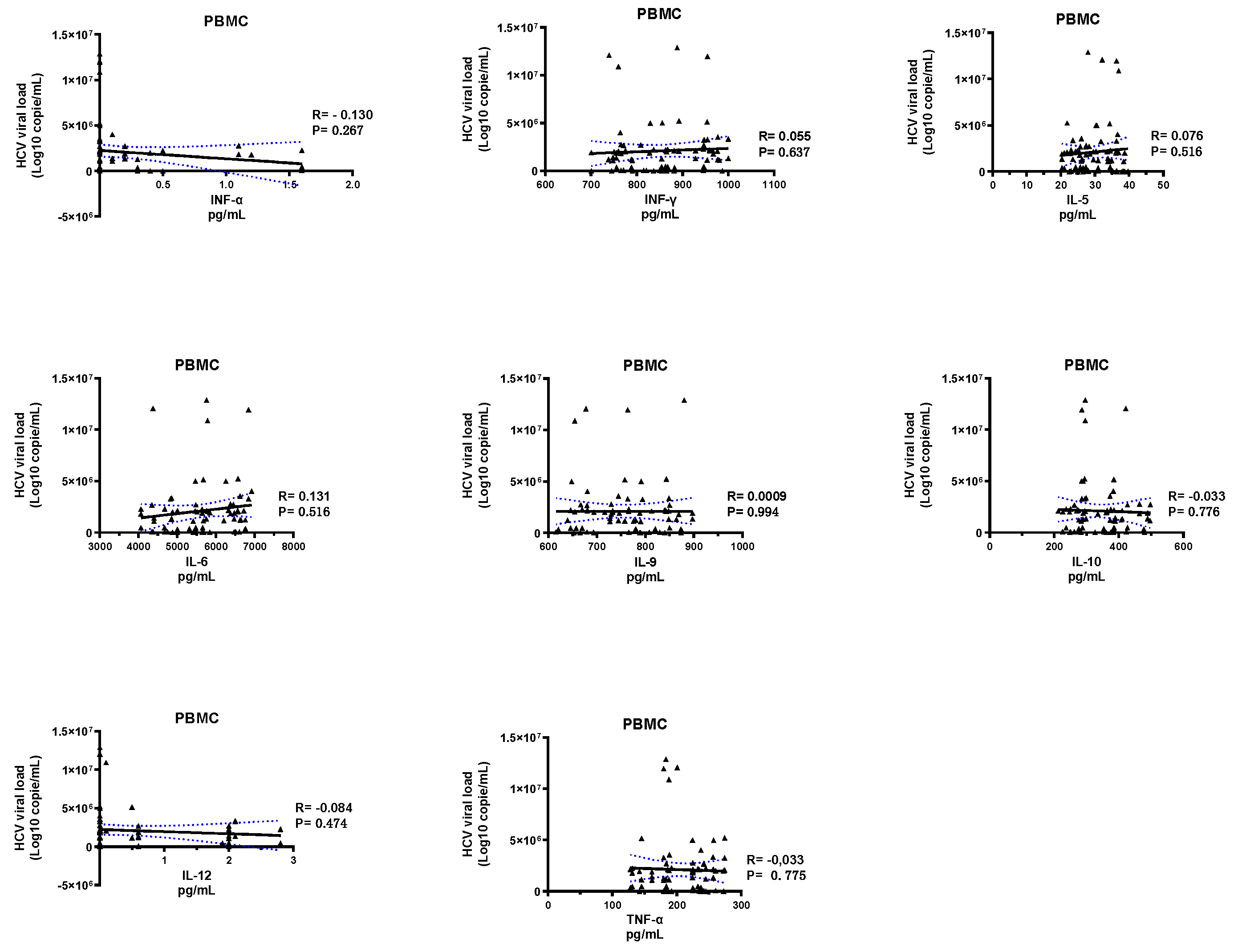
2.2. Comparison of Levels of Cytokines Produced by PBMC and Neutrophils from HCV+ Patients and Healthy Subjects
2.3. Th1–Th2 Cytokine Ratios
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Isolation of PBMC
4.3. Isolation of Neutrophils
4.4. Mitogen Stimulation
4.5. Evaluation of Cytokines
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kiermaier, E.; Sixt, M. Fragmented communication between immune cells. Science 2015, 349, 1055–1056. [Google Scholar] [CrossRef]
- Galani, I.E.; Andreakos, E. Neutrophils in viral infections: Current concepts and caveats. J. Leukoc. Biol. 2015, 98, 557–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naumenko, V.; Turk, M.; Jenne, C.N.; Kim, S.J. Neutrophils in viral infection. Cell Tissue Res. 2018, 371, 505–516. [Google Scholar] [CrossRef]
- Wisniewska-Ligier, M.; Wozniakowska-Gesicka, T.; Glowacka, E.; Lewkowicz, P.; Banasik, M.; Tchorzewski, H. Involvement of innate immunity in the pathogenesis of chronic hepatitis C in children. Scand. J. Immunol. 2006, 64, 425–432. [Google Scholar] [CrossRef]
- Tamassia, N.; Bianchetto-Aguilera, F.; Arruda-Silva, F.; Gardiman, E.; Gasperini, S.; Calzetti, F.; Cassatella, M.A. Cytokine production by human neutrophils: Revisiting the “dark side of the moon”. Eur. J. Clin. Investig. 2018, 48, e12952. [Google Scholar] [CrossRef]
- Rosen, H.R.; Golden-Mason, L. Control of HCV Infection by Natural Killer Cells and Macrophages. Cold Spring Harb. Perspect. Med. 2020, 10, a037101. [Google Scholar] [CrossRef]
- Norian, R.; Delirezh, N.; Azadmehr, A. Evaluation of proliferation and cytokines production by mitogen-stimulated bovine peripheral blood mononuclear cells. Vet. Res. Forum 2015, 6, 265–271. [Google Scholar]
- Sobue, S.; Nomura, T.; Ishikawa, T. Th1/Th2 cytokine profiles and their relationship to clinical features in patients with chronic hepatitis C virus infection. J. Gastroenterol. 2001, 36, 544–551. [Google Scholar] [CrossRef]
- Falasca, K.; Mancino, P.; Ucciferri, C.; Dalessandro, M.; Zingariello, P.; Lattanzio, F.M.; Petrarca, C.; Martinotti, S.; Pizzigallo, E.; Conti, P.; et al. Inflammatory cytokines and S-100b protein in patients with hepatitis C infection and cryoglobulinemias. Clin. Investig. Med. 2007, 30, 167–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gigi, E.; Raptopoulou-Gigi, M.; Kalogeridis, A.; Masiou, S.; Orphanou, E.; Vrettou, E.; Lalla, T.H.; Sinakos, E.; Tsapas, V. Cytokine mRNA expression in hepatitis C virus infection: TH1 predominance in patients with chronic hepatitis C and TH1-TH2 cytokine profile in subjects with self-limited disease. J. Viral Hepat. 2008, 15, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.G.; Golden-Mason, L.; Rutebemberwa, A.; Rosen, H.R. Interleukin (IL)-17/IL-22producing T cells enriched within the liver of patients with chronic hepatitis C viral (HCV) infection. Dig. Dis. Sci. 2012, 57, 381–389. [Google Scholar] [CrossRef]
- Gramenzi, A.; Andreone, P.; Loggi, E.; Foschi, F.G.; Cursaro, C.; Margotti, M.; Biselli, M.; Bernardi, M. Cytokine profile of peripheral blood mononuclear cells from patients with different outcomes of hepatitis C virus infection. J. Viral Hepat. 2005, 12, 525–530. [Google Scholar] [CrossRef]
- Day, C.L.; Lauer, G.M.; Robbins, G.K.; McGovern, B.; Wurcel, A.G.; Gandhi, R.T.; Walker, B.D. Broad specificity of virus specific CD4+ T-helper-cell responses in resolved hepatitis C virus infection. J. Virol. 2002, 76, 12584–12595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larrubia, J.R.; Benito-Martínez, S.; Miquel-Plaza, J.E.; Sanz-de-Villalobos, E.; González-Mateos, F.; Parra, T. Cytokines—their pathogenic and therapeutic role in chronic viral hepatitis. Rev. Esp Enferm Dig. 2009, 101, 343–351. [Google Scholar] [CrossRef]
- Cavalcante-Silva, L.H.A.; Carvalho, D.C.M.; Lima, E.A.; Galvão, J.G.F.M.; da Silva, J.S.F.; Sales-Neto, J.M.; Rodrigues-Mascarenhas, S. Neutrophils and COVID-19: The road so far. Int. Immunopharmacol. 2021, 90, 107233. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Stohlman, S.A.; Hinton, D.R.; Marten, N.W. Neutrophils promote mononuclear cell infiltration during viral-induced encephalitis. J. Immunol. 2003, 170, 3331–3336. [Google Scholar] [CrossRef] [PubMed]
- Tumpey, T.M.; Chen, S.H.; Oakes, J.E.; Lausch, R.N. Neutrophil-mediated suppression of virus replication after herpes simplex virus type 1 infection of the murine cornea. J. Virol. 1996, 70, 898–904. [Google Scholar] [CrossRef] [Green Version]
- Grist, J.J.; Marro, B.; Lane, T.E. Neutrophils and viral-induced neurologic disease. Clin. Immunol. 2018, 189, 52–56. [Google Scholar] [CrossRef]
- Niessen, A.; Heyder, P.; Krienke, S.; Blank, N.; Tykocinski, L.O.; Lorenz, H.M.; Schiller, M. Apoptoticcell-derived membrane microparticles and IFN-α induce an inflammatory immune response. J. Cell Sci. 2015, 128, 2443–2453. [Google Scholar]
- Maródi, L.; Káposzta, R.; Nemes, E. Survival of group B streptococcus type III in mononuclear phagocytes: Differential regulation of bacterial killing in cord macrophages by human recombinant gamma interferon and granulocyte-macrophage colony-stimulating factor. Infect. Immun. 2000, 68, 2167–2170. [Google Scholar] [CrossRef] [Green Version]
- Groepper, C.; Rufinatscha, K.; Schroder, N.; Stindt, S.; Ehlting, C.; Albrecht, U.; Bock, H.H.; Bartenschlager, R.; Häussinger, D.; Bode, J.G. HCV modifies EGF signaling and upregulates production of CXCR2 ligands: Role in inflammation and antiviral immune response. J. Hepatol. 2018, 69, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, Q.; Shao, X.; Wang, W.; Zhang, C.; Jin, Z. An immunosuppressive function of interleukin-35 in chronic hepatitis C virus infection. Int. Immunopharmacol. 2017, 50, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Kaukinen, P.; Sillanpää, M.; Kotenko, S.; Lin, R.; Hiscott, J.; Melen, K.; Julkunen, I. Hepatitis C virus NS2 and NS3/4A proteins are potent inhibitors of host cell cytokine/chemokine gene expression. Virol. J. 2006, 3, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andronescu, D.; Diaconu, S.; Tiuca, N.; Purcarea, R.M.; Andronescu, C.I. Hepatitis C treatment & management. J. Med. Life 2014, 7, 31–36. [Google Scholar] [PubMed]
- Fallahi, P.; Ferri, C.; Ferrari, S.M.; Corrado, A.; Sansonno, D.; Antonelli, A. Cytokines and HCV-related disorders. Clin. Dev. Immunol. 2012, 2012, 468107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, M.; Wan, M.; Guo, S.; Sun, R.; Yang, M.; Zhao, T.; Yan, Y.; Zhang, Y.; Huang, W.; Wu, X.; et al. An oligodeoxynucleotide capable of lessening acute lung inflammatory injury in mice infected by influenza virus. Biochem. Biophys. Res. Commun. 2011, 415, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Guidotti, L.G.; Chisari, F.V. Immunobiology and pathogenesis of viral hepatitis. Annu. Rev. Pathol. 2006, 1, 23–61. [Google Scholar] [CrossRef] [Green Version]
- Rehermann, B. Natural Killer Cells in Viral Hepatitis. Cell Mol. Gastroenterol. Hepatol. 2015, 1, 578–588. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, R.; Meyer, K.; Moriyama, M.; Kato, N.; Yokosuka, O.; Ray, R.B.; Aurora, R.; Ray, R.; Kanda, T. Rapid hepatitis C virus clearance by antivirals correlates with immune status of infected patients. J. Med. Virol. 2019, 91, 411–418. [Google Scholar] [CrossRef]
- Mourtzikoua, A.; Alepakia, M.; Stamoulic, M.; Pouliakisa, A.; Sklirisc, A.; Karakitsosa, P. Evaluation of serum levels of IL-6, TNF-α, IL-10, IL-2 and IL-4 in patients with chronic hepatitis. Immunología 2014, 33, 41–50. [Google Scholar] [CrossRef]
- Semmo, N.; Day, C.L.; Ward, S.M.; Lucas, M.; Harcourt, G.; Loughry, A.; Klenerman, P. Preferential loss of IL-2-secreting CD4+ T helper cells in chronic HCV infection. Hepatology 2005, 41, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Toumpanakis, D.; Vassilakkopoulos, T. Molecular mechanisms of action of interleukin-6 (IL-6). Pneumon 2007, 2, 154–167. [Google Scholar]
- Yoshida, H.; Hashizume, M.; Suzuki, M.; Mihara, M. Anti-IL-6 receptor antibody suppressed T cell activation by inhibiting IL-2 production and inducing regulatory T cells. Eur. J. Pharmacol. 2010, 634, 178–183. [Google Scholar] [CrossRef]
- Naseem, S.; Manzoor, S.; Javed, A.; Abbas, S. Interleukin-6 Rescues Lymphocyte from Apoptosis and Exhaustion Induced by Chronic Hepatitis C Virus Infection. Viral. Immunol. 2018, 31, 624–631. [Google Scholar] [CrossRef]
- Yoneda, S.; Umemura, T.; Katsuyama, Y.; Kamijo, A.; Joshita, S.; Komatsu, M. Association of serum cytokine levels with treatment response to pegylated interferon and ribavirin therapy in genotype 1 chronic hepatitis C patients. J. Infect. Dis. 2011, 203, 1087–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heymann, F.; Tacke, F. Immunology in the liver-from homeostasis to disease. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 88–110. [Google Scholar] [CrossRef]
- Ali, M.E.; El-Badawy, O.; Afifi, N.A.; Eldin, A.S.; Hassan, E.A.; Halby, H.M.; El-Mokhtar, M.A. Role of T-Helper 9 Cells in Chronic Hepatitis C-Infected Patients. Viruses 2018, 10, 341. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Fulgencio, M.; Jiménez, J.L.; Berenguer, J.; Fernández-Rodríguez, A.; López, J.C.; Cosín, J.; Miralles, P.; Micheloud, D.; Munoz-Fernández, M.A.; Resino, S. Plasma Il-6 and Il-9 predict the failure of interferon-α plus ribavirin therapy in HIV/HCV-coinfected patients. J. Antimicrob. Chemother. 2012, 67, 1238–1245. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, T.; Hirahara, K.; Onodera, A.; Endo, Y.; Hosokawa, H.; Shinoda, K.; Tumes, D.J.; Okamoto, Y. Th2 Cells in Health and Disease. Annu. Rev. Immunol. 2017, 35, 53–84. [Google Scholar] [CrossRef]
- Zhang, L.; Hao, C.Q.; Miao, L.; Dou, X.G. Role of Th1/Th2 cytokines in serum on the pathogenesis of chronic hepatitis C and the outcome of interferon therapy. Genet. Mol. Res. 2014, 13, 9747–9755. [Google Scholar] [CrossRef]
- Radkowski, M.; Bednarska, A.; Horban, A.; Stanczak, J.; Wilkinson, J.; Adair, D.M.; Nowicki, M.; Rakela, J.; Laskus, T. Infection of primary human macrophages with hepatitis C virus in vitro: Induction of tumour necrosis factor-alpha and interleukin 8. J. Gen. Virol. 2004, 85, 47–59. [Google Scholar] [CrossRef]
- Hammerstad, S.S.; Blackard, J.T.; Lombardi, A.; Owen, R.P.; Concepcion, E.; Yi, Z.; Zhang, W.; Tomer, Y. Hepatitis C Virus Infection of Human Thyrocytes: Metabolic, Hormonal, and Immunological Implications. J. Clin. Endocrinol. Metab. 2020, 105, 1157–1168. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Zhang, H.F.; Chen, H.; Li, J.; Yang, X.J.; Zhu, S.S.; Cheng, Y. Level of the cytokine secreted by peripheral blood mononuclear cells in patients with chronic hepatitis C before antiviral therapy. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi 2008, 22, 364–366. [Google Scholar] [PubMed]
- Fan, X.G.; Tang, F.Q.; Yi, H.; Liu, W.E.; Houghton, M.; Hu, G.L. Effect of IL-12 on T-cell immune responses in patients with chronic HCV infection. APMIS 2000, 108, 531–538. [Google Scholar] [CrossRef]
- Ahmed, F.; Ibrahim, A.; Cooper, C.L.; Kumar, A.; Crawley, A.M. Chronic Hepatitis C Virus Infection Impairs M1 Macrophage Differentiation and Contributes to CD8(+) T-Cell Dysfunction. Cells 2019, 8, 374. [Google Scholar] [CrossRef] [Green Version]
- Yue, M.; Deng, X.; Zhai, X.; Xu, J.; Kong, K.; Zhang, J. Th1 and Th2 cytokine profiles induced by hepatitis C virus F protein in peripheral blood mononuclear cells from chronic hepatitis C patients. Immunol. Lett. 2013, 152, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Cornberg, M.; Wedemeyer, H. Hepatitis C virus infection from the perspective of heterologous immunity. Curr. Opin. Virol. 2016, 16, 41–48. [Google Scholar] [CrossRef]
- Essa, S.; Raghupathy, R.; Pacsa, A.S.; El-Shazly, A.; Said, T. Changes in cell-mediated immunity in kidney transplant recipients with active CMV infection. FEMS Immunol. Med. Microbiol. 2002, 32, 199–204. [Google Scholar] [CrossRef]
- Oh, H.; Siano, B.; Diamond, S. Neutrophil Isolation Protocol. J. Vis. Exp. 2008, 17, 745–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spirig, R.; Schaub, A.; Kropf, A.; Miescher, S.; Spycher, M.O.; Rieben, R. Reconstituted high-density lipoprotein modulates activation of human leukocytes. PLoS ONE 2013, 8, e71235. [Google Scholar]
- Dallegri, F.; Frumento, G.; Maggi, A.; Patrone, F. PHA-induced neutrophil-mediated cytotoxicity. Clin. Lab. Immunol. 1983, 11, 203–206. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | HCV− | HCV+ |
|---|---|---|
| No. | 75 | 86 |
| Gender (M/F) | 39/36 | 45/40 |
| Age (mean ± SD) [range], years | 32 ± 10.35 (32–55) | 39 ± 9.21 (32–55) |
| Viral load mean ± SD [range], IU/mL | ND | 2.1 × 106 ± 2.7 × 106 (325–11,948,083) |
| HCV genotype (1/3/4) | ND | 4/7/75 |
| AST (mean ± SD, IU/mL) | ND | 64.58 ± 33.45 |
| ALT (mean ± SD, IU/mL) | ND | 82.78 ± 55.45 |
| ALP mean ± SD, IU/mL | ND | 152.65 ± 39.77 |
| Th1/Th2 | PBMC | Neutrophils | ||
|---|---|---|---|---|
| HCV− | HCV+ | HCV− | HCV+ | |
| IL-2/IL-4 | 13 | 100 | 10 | 580 |
| IL-2/IL-5 | 2.3 | 0.003 | 0.004 | 0.12 |
| IL-2/IL-9 | 0.03 | 0.0001 | 0.000003 | 0.03 |
| IL-2/IL-10 | 0.05 | 0.0003 | 0.0002 | 0.2 |
| TNF/IL-4 | 300 | 224,600 | 85,500 | 16,980 |
| TNF /IL-5 | 53.8 | 7.6 | 32.9 | 3.4 |
| TNF/IL-9 | 0.8 | 0.3 | 0.2 | 0.9 |
| TNF/IL-10 | 1.1 | 0.7 | 1.9 | 6.7 |
| IL-6/IL-4 | 7065.3 | 5,654,200 | 9,964,700 | 132,510 |
| IL-6/IL-5 | 1268.1 | 191.7 | 3832.6 | 26.9 |
| IL-6/IL-9 | 18.5 | 7.4 | 26.6 | 6.7 |
| IL-6/IL-10 | 25.4 | 17.3 | 219 | 52.2 |
| IL-12/IL-4 | 0.00007 | 0.1 | 300 | 270 |
| IL-12 /IL-5 | 0.00001 | 0.000003 | 0.1 | 0.05 |
| IL-12/IL-9 | 0.0000002 | 0.0000001 | 0.0008 | 0.01 |
| IL-12/IL-10 | 0.0000003 | 0.0000003 | 0.007 | 0.1 |
| IFN-α/IL-4 | 0.7 | 0.1 | 0.1 | 0.01 |
| IFN-α/IL-5 | 0.13 | 0.000003 | 0.00004 | 0.000002 |
| IFN-α/IL-9 | 0.0019 | 0.0000001 | 0.0000003 | 0.0000005 |
| IFN-α/IL-10 | 0.003 | 0.0000003 | 0.000002 | 0.000004 |
| IFN-γ/IL-4 | 1196 | 854,500 | 451,200 | 41,740 |
| IFN-γ/IL-5 | 215 | 29 | 173.5 | 8.5 |
| IFN-γ/IL-9 | 3.1 | 1.1 | 1.2 | 2.1 |
| IFN-γ/IL-10 | 4.3 | 2.6 | 9.9 | 16.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Essa, S.; Siddique, I.; Saad, M.; Raghupathy, R. Modulation of Production of Th1/Th2 Cytokines in Peripheral Blood Mononuclear Cells and Neutrophils by Hepatitis C Virus Infection in Chronically Infected Patients. Pathogens 2021, 10, 1519. https://doi.org/10.3390/pathogens10111519
Essa S, Siddique I, Saad M, Raghupathy R. Modulation of Production of Th1/Th2 Cytokines in Peripheral Blood Mononuclear Cells and Neutrophils by Hepatitis C Virus Infection in Chronically Infected Patients. Pathogens. 2021; 10(11):1519. https://doi.org/10.3390/pathogens10111519
Chicago/Turabian StyleEssa, Sahar, Iqbal Siddique, Motaz Saad, and Raj Raghupathy. 2021. "Modulation of Production of Th1/Th2 Cytokines in Peripheral Blood Mononuclear Cells and Neutrophils by Hepatitis C Virus Infection in Chronically Infected Patients" Pathogens 10, no. 11: 1519. https://doi.org/10.3390/pathogens10111519