
Molecular Epidemiology of Anisakis spp. in Wedge Sole, Dicologlossa cuneata (Moreau, 1881), from Fishmarkets in Granada (Southern Spain), Caught in Two Adjacent NE and CE Atlantic Areas

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Host
2.2. Morphological and Molecular Identification. Epidemiological Parameters
3. Discussion
4. Materials and Methods
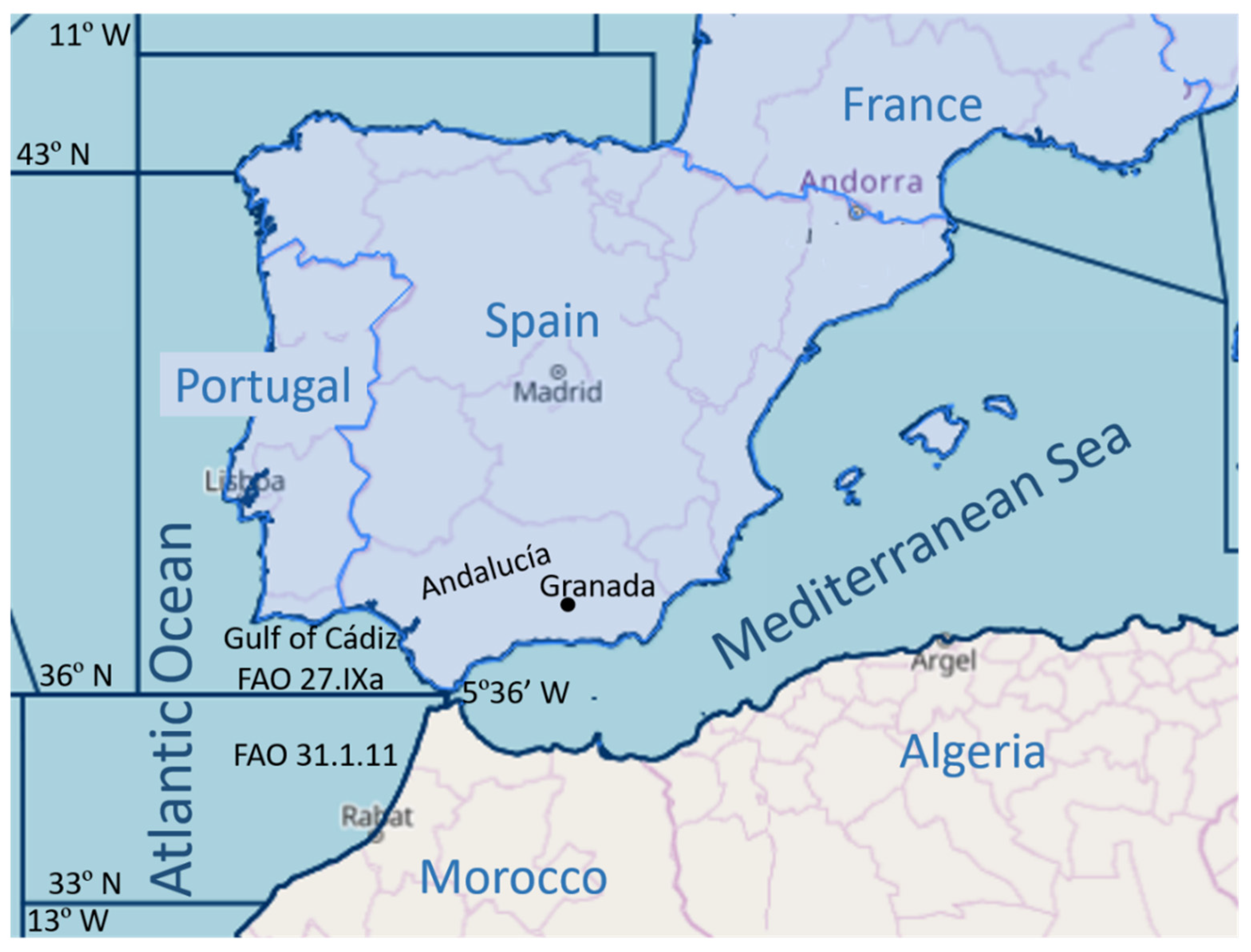
4.1. Hosts and Search for Parasites
4.2. Genetic Identification of Anisakis spp. Larvae
4.3. Epidemiological Parameters and Statistical Comparisons
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rello Yubero, F.J.; Adroher Auroux, F.J.; Valero-López, A. Anisákidos parásitos de peces comerciales. Riesgos asociados a la salud pública. An. Real Acad. Cienc. Vet. Andal. Orient. 2004, 17, 173–198. [Google Scholar]
- Ishikura, H.; Kikuchi, K.; Nagasawa, K.; Ooiwa, T.; Takamiya, H.; Sato, N.; Sugane, K. Anisakidae and anisakidosis. In Progress in Clinical Parasitology; Sun, T., Ed.; Springer-Verlag Inc.: New York, NY, USA, 1993; Volume 3, pp. 43–102. ISBN 978-1-4612-7646-3. [Google Scholar]
- Adroher-Auroux, F.J.; Benítez-Rodríguez, R. Anisakiasis and Anisakis: An underdiagnosed emerging disease and its main etiological agents. Res. Vet. Sci. 2020, 132, 535–545. [Google Scholar] [CrossRef] [PubMed]
- McClelland, G.; Misra, R.K.; Martell, D.J. Variations in abundance of larval anisakines, sealworm (Pseudoterranova decipiens) and related species, in Eastern Canadian cod and flatfish. Can. Tech. Rep. Fish. Aquat. Sci. 1985, 1392, xi + 57. [Google Scholar]
- Marques, J.F.; Santos, M.J.; Cabral, H.N. Zoogeographical patterns of flatfish (Pleuronectiformes) parasites in the Northeast Atlantic and the importance of the Portuguese coast as a transitional area. Sci. Mar. 2009, 73, 461–471. [Google Scholar] [CrossRef] [Green Version]
- McClelland, G.; Misra, R.K.; Marcogliese, D.J. Variations in abundance of larval anisakines, sealworm (Phocanema decipiens) and related species in cod and flatfish from the southern Gulf of St. Lawrence (4T) and the Breton Shelf (4Vn). Can. Tech. Rep. Fish. Aquat. Sci. 1983, 1201, ix+51. [Google Scholar]
- Marques, J.F.; Santos, M.J.; Teixeira, C.M.; Batista, M.I.; Cabral, H.N. Host-parasite relationships in flatfish (Pleuronectiformes)—The relative importance of host biology, ecology and phylogeny. Parasitology 2011, 138, 107–121. [Google Scholar] [CrossRef]
- Oliva, M.E.; Castro, R.E.; Burgos, R. Parasites of the flatfish Paralichthys adspersus (Steindachner, 1867) (Pleuronectiformes) from northern Chile. Mem. Inst. Oswaldo Cruz 1996, 91, 301–306. [Google Scholar] [CrossRef] [Green Version]
- González, M.T.; Acuña, E.; Oliva, M.E. Metazoan parasite fauna of the bigeye flounder, Hippoglossina macrops, from northern Chile. Influence of host age and sex. Mem. Inst. Oswaldo Cruz 2001, 96, 1049–1054. [Google Scholar] [CrossRef] [Green Version]
- Alarcos, A.J.; Pereira, A.N.; Taborda, N.L.; Luque, J.L.; Timi, J.T. Parasitological evidence of stocks of Paralichthys isosceles (Pleuronectiformes: Paralichthyidae) at small and large geographical scales in South American Atlantic coasts. Fish. Res. 2016, 173, 221–228. [Google Scholar] [CrossRef]
- Álvarez, F.; Iglesias, R.; Paramá, A.I.; Leiro, J.; Sanmartín, M. Abdominal macroparasites of commercially important flatfishes (Teleostei: Scophthalmidae, Pleuronectidae, Soleidae) in northwest Spain (ICES IXa). Aquaculture 2002, 213, 31–53. [Google Scholar] [CrossRef]
- Marques, J.F.; Santos, M.J.; Cabral, H.N. Soleidae macroparasites along the Portuguese coast: Latitudinal variation and host–parasite associations. Mar. Biol. 2006, 150, 285–298. [Google Scholar] [CrossRef]
- Marques, J.F.; Cabral, H.N.; Busi, M.; D’Amelio, S. Molecular identification of Anisakis species from Pleuronectiformes off the Portuguese coast. J. Helminthol. 2006, 80, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Seesao, Y. Caractérisation des Anisakidae Dans les Poissons Marins: Développement D’une Méthode D’identification Par Séquençage à Haut-Débit et Étude de Prévalence. Ph.D. Thesis, Université du Droit et de la Santé-Lille II, Lille, France, 2015. [Google Scholar]
- Silva, M.E.R.; Eiras, J.C. Occurrence of Anisakis sp. in fishes off the Portuguese West coast and evaluation of its zoonotic potential. Bull. Eur. Assoc. Fish Pathol. 2003, 23, 13–17. [Google Scholar]
- IDAPES Consulta estadísticas pesqueras. Primera Venta de Pesca Fresca en Lonja. Available online: http://www.juntadeandalucia.es/agriculturaypesca/idapes/ (accessed on 9 September 2021).
- Fulton, T.W. The rate of growth of fishes. In Proceedings of the 22nd Annual Report of the Fishery Board of Scotland; Edinburgh, UK; 1904, Volume 3, pp. 141–241.
- Nash, R.D.M.; Valencia, A.H.; Geffen, A.J. The origin of Fulton’s condition factor—Setting the record straight. Fisheries 2006, 31, 236–238. [Google Scholar]
- Berland, B. Nematodes from some Norwegian marine fishes. Sarsia 1961, 2, 1–50. [Google Scholar] [CrossRef]
- Quéro, J.-C.; Desoutter, M.; Lagardère, F. Soleidae. In Fishes of the North-Eastern Atlantic and the Mediterranean; Whitehead, P.J.P., Bauchot, M.-L., Hureau, J.-C., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, France, 1986; Volume 3, pp. 1308–1324. [Google Scholar]
- Belghyti, D.; Berrada-rkhami, O.; Boy, V.; Aguesse, P.; Gabrion, C. Population biology of two helminth parasites of flatfishes from the Atlantic coast of Morocco. J. Fish Biol. 1994, 44, 1005–1021. [Google Scholar] [CrossRef]
- Belghyti, D.; Aguesse, P.; Gabrion, C. Éthologie alimentaire de Citharus linguatula et Dicologoglossa cuneata sur les côtes atlantiques du Maroc. Vie Milieu/Life Environ. 1993, 43, 95–108. [Google Scholar]
- Adroher-Auroux, F.J.; Benítez-Rodríguez, R. Hysterothylacium aduncum. In Fish Parasites: A Handbook of Protocols for Their Isolation, Culture and Transmission; Sitjà-Bobadilla, A., Bron, J.E., Wiegertjes, G., Piazzon, M.C., Eds.; European Association of Fish Pathologists (EAFP)/5m Books Series; 5M Books Ltd.: Great Easton, UK, 2021; pp. 311–329. ISBN 9781789181333. [Google Scholar]
- Rello, F.J.; Adroher, F.J.; Benítez, R.; Valero, A. The fishing area as a possible indicator of the infection by anisakids in anchovies (Engraulis encrasicolus) from southwestern Europe. Int. J. Food Microbiol. 2009, 129, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Molina-Fernández, D.; Malagón, D.; Gómez-Mateos, M.; Benítez, R.; Martín-Sánchez, J.; Adroher, F.J. Fishing area and fish size as risk factors of Anisakis infection in sardines (Sardina pilchardus) from Iberian waters, southwestern Europe. Int. J. Food Microbiol. 2015, 203, 27–34. [Google Scholar] [CrossRef]
- Adroher, F.J.; Valero, A.; Ruiz-Valero, J.; Iglesias, L. Larval anisakids (Nematoda: Ascaridoidea) in horse mackerel (Trachurus trachurus) from the fish market in Granada (Spain). Parasitol. Res. 1996, 82, 253–256. [Google Scholar] [CrossRef]
- Levsen, A.; Karl, H. Anisakissimplex (s.l.) in grey gurnard (Eutrigla gurnardus) from the North Sea: Food safety considerations in relation to fishing ground and distribution in the flesh. Food Control 2014, 36, 15–19. [Google Scholar] [CrossRef]
- Brattey, J.; Bishop, C.A. Larval Anisakis simplex (Nematoda: Ascaridoidea) infection in the musculature of Atlantic cod, Gadus morhua, from Newfoundland and Labrador. Can. J. Fish. Aquat. Sci. 1992, 49, 2635–2647. [Google Scholar] [CrossRef]
- Molina-Fernández, D.; Rubio-Calvo, D.; Adroher, F.J.; Benítez, R. Molecular epidemiology of Anisakis spp. in blue whiting Micromesistius poutassou in eastern waters of Spain, western Mediterranean Sea. Int. J. Food Microbiol. 2018, 282, 49–56. [Google Scholar] [CrossRef]
- Rello, F.J.; Valero, A.; Adroher, F.J. Anisakid parasites of the pouting (Trisopterus luscus) from the Cantabrian Sea coast, Bay of Biscay, Spain. J. Helminthol. 2008, 82, 287–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bussmann, B.; Ehrich, S. Investigations on infestation of blue whiting (Micromesistius poutassou) with larval Anisakis sp. (Nematoda: Ascaridida). Arch. Fischereiwiss. 1979, 29, 155–165. [Google Scholar]
- Davey, J.T. The incidence of Anisakis sp. larvae (Nematoda: Ascaridata) in the commercially exploited stocks of herring (Clupea harengus L., 1758,) (Pisces: Clupeidae) in British and adjacent waters. J. Fish Biol. 1972, 4, 535–554. [Google Scholar] [CrossRef]
- Valero, A.; Martín-Sánchez, J.; Reyes-Muelas, E.; Adroher, F.J. Larval anisakids parasitizing the blue whiting, Micromesistius poutassou, from Motril Bay in the Mediterranean region of southern Spain. J. Helminthol. 2000, 74, 361–364. [Google Scholar] [CrossRef] [Green Version]
- Eltink, A. Anisakis larvae (Nematoda: Ascaridida) in mackerel, (Scomber scombrus L.) in lCES sub-areas IV, VI, VII and VIII in 1970-1971 and 1982–1984. Int. Counc. Explor. Sea 1988, CM1988/H23, 29p. [Google Scholar] [CrossRef]
- Smith, J.W. Anisakissimplex (Rudolphi, 1809, det. Krabbe, 1878): Length distribution and viability of L3 of known minimum age from herring Clupea harengus L. J. Helminthol. 1984, 58, 337–340. [Google Scholar] [CrossRef]
- Monstad, T. Some aspects of mortality, condition factors and liver state with Anisakis-infection in blue whiting in the North-East Atlantic. In Proceedings of the Fourth Soviet-Norwegian Symposium, Bergen, Norway, 12–16 June 1990; Monstad, T., Ed.; Institute of Marine Research: Bergen, Norway, 1990; pp. 319–339. [Google Scholar]
- Serrat, A.; Lloret, J.; Frigola-Tepe, X.; Muñoz, M. Trade-offs between life-history traits in a coldwater fish in the Mediterranean Sea: The case of blue whiting Micromesistius poutassou. J. Fish Biol. 2019, 95, 428–443. [Google Scholar] [CrossRef]
- Richards, J. Preliminary results of the 1977 blue whiting surveys of the west of Scotland. Int. Counc. Explor. Sea 1977, CM1977/H33, 15. [Google Scholar]
- Rohde, K. Ecology of marine parasites. Helgoländer Meeresunters. 1984, 37, 5–33. [Google Scholar] [CrossRef] [Green Version]
- Mouritsen, K.N.; Hedeholm, R.; Schack, H.B.; Møller, L.N.; Storr-Paulsen, M.; Dzido, J.; Rokicki, J. Occurrence of anisakid nematodes in Atlantic cod (Gadus morhua) and Greenland cod (Gadus ogac), West Greenland. Acta Parasitol. 2010, 55, 81–89. [Google Scholar] [CrossRef]
- De la Torre Molina, R.; Pérez Aparicio, J.; Hernández Bienes, M.; Jurado Pérez, R.; Martínez Ruso, A.; Morales Franco, E. Anisakiasis en pescados frescos comercializados en el norte de Córdoba. Rev. Esp. Salud Pública 2000, 74, 517–526. [Google Scholar] [CrossRef] [Green Version]
- Abattouy, N.; Valero, A.; Benajiba, M.H.; Lozano, J.; Martín-Sánchez, J. Anisakissimplex s.l. parasitization in mackerel (Scomber japonicus) caught in the North of Morocco—Prevalence and analysis of risk factors. Int. J. Food Microbiol. 2011, 150, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Biary, A.; Berrouch, S.; Dehhani, O.; Maarouf, A.; Sasal, P.; Mimouni, B. Prevalence and identification of Anisakis nematodes in fish consumed in Marrakesh, Morocco. Mol. Biol. Rep. 2021, 48, 3417–3422. [Google Scholar] [CrossRef] [PubMed]
- Abattouy, N.; Valero-López, A.; Lozano-Maldonado, J.; Benajiba, M.H.; Martín-Sánchez, J. Epidemiology and molecular identification of Anisakis pegreffii (Nematoda: Anisakidae) in the horse mackerel Trachurus trachurus from northern Morocco. J. Helminthol. 2014, 88, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Cipriani, P.; Acerra, V.; Bellisario, B.; Sbaraglia, G.L.; Cheleschi, R.; Nascetti, G.; Mattiucci, S. Larval migration of the zoonotic parasite Anisakis pegreffii (Nematoda: Anisakidae) in European anchovy, Engraulis encrasicolus: Implications to seafood safety. Food Control 2016, 59, 148–157. [Google Scholar] [CrossRef] [Green Version]
- Farjallah, S.; Busi, M.; Mahjoub, M.O.; Slimane, B.B.; Paggi, L.; Said, K.; D’Amelio, S. Molecular characterization of larval anisakid nematodes from marine fishes off the Moroccan and Mauritanian coasts. Parasitol. Int. 2008, 57, 430–436. [Google Scholar] [CrossRef]
- Mattiucci, S.; Nascetti, G.; Cianchi, R.; Paggi, L.; Arduino, P.; Margolis, L.; Brattey, J.; Webb, S.; D’Amelio, S.; Orecchia, P.; et al. Genetic and ecological data on the Anisakis simplex complex, with evidence for a new species (Nematoda, Ascaridoidea, Anisakidae). J. Parasitol. 1997, 83, 401. [Google Scholar] [CrossRef]
- Abollo, E.; Gestal, C.; Pascual, S. Anisakis infestation in marine fish and cephalopods from Galician waters: An updated perspective. Parasitol. Res. 2001, 87, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Mattiucci, S.; Cipriani, P.; Levsen, A.; Paoletti, M.; Nascetti, G. Molecular epidemiology of Anisakis and anisakiasis: An ecological and evolutionary road map. Adv. Parasitol. 2018, 99, 93–263. [Google Scholar] [CrossRef] [PubMed]
- Hartwich, G. Keys to genera of the Ascaridoidea. In CIH Keys to the Nematode Parasites of Vertebrates. No. 2; Anderson, R., Chabaud, A., Willmott, S., Eds.; CAB: Slough, UK, 1974; p. iv + 15. [Google Scholar]
- Yoshinaga, T.; Ogawa, K.; Wakabayashi, H. Experimental life cycle of Hysterothylacium aduncum (Nematoda: Anisakidae) in fresh water. Fish Pathol. 1987, 22, 243–251. [Google Scholar] [CrossRef]
- Petter, A.J.; Maillard, C. Larves d’ascarides parasites de poissons en Méditerranée occidentale. Bull. Muséum Natl. D’histoire Nat. 1988, 10A, 347–369. [Google Scholar]
- Zhu, X.-Q.; Gasser, R.B.; Podolska, M.; Chilton, N.B. Characterisation of anisakid nematodes with zoonotic potential by nuclear ribosomal DNA sequences. Int. J. Parasitol. 1998, 28, 1911–1921. [Google Scholar] [CrossRef]
- D’Amelio, S.; Mathiopoulos, K.D.; Santos, C.P.; Pugachev, O.N.; Webb, S.C.; Picanço, M.; Paggi, L. Genetic markers in ribosomal DNA for the identification of members of the genus Anisakis (Nematoda: Ascaridoidea) defined by polymerase chain reaction-based restriction fragment length polymorphism. Int. J. Parasitol. 2000, 30, 223–226. [Google Scholar] [CrossRef]
- Pontes, T.; D’Amelio, S.; Costa, G.; Paggi, L. Molecular characterization of larval anisakid nematodes from marine fishes of Madeira by a PCR-based approach, with evidence for a new species. J. Parasitol. 2005, 91, 1430–1434. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Reiczigel, J.; Rózsa, L. Quantitative Parasitology 3.0. Software 2005. Available online: https://www.zoologia.hu/qp/ (accessed on 15 January 2021).
- Rózsa, L.; Reiczigel, J.; Majoros, G. Quantifying parasites in samples of hosts. J. Parasitol. 2000, 86, 228–232. [Google Scholar] [CrossRef]
- Reiczigel, J.; Marozzi, M.; Fábián, I.; Rózsa, L. Biostatistics for parasitologists—A primer to Quantitative Parasitology. Trends Parasitol. 2019, 35, 277–281. [Google Scholar] [CrossRef]
- Luna, S.M. Dicologlossa cuneata (Moreau 1881) Wedge Sole. Available online: http://fishbase.org/summary/Dicologlossa-cuneata.html (accessed on 17 June 2021).


{kind=link}
| Parameters | Both Zones | Zone 27.IXa | Zone 34.1.11 | |
|---|---|---|---|---|
| Host | ||||
| Wedge sole | No. of fish (F) | 110 | 70 | 40 |
| Mean weight ± SD (range) | 57.9 ± 38.0 (21.8–170.0) | 34.2 ± 6.8 (21.8–56.5) | 99.4 ± 34.4 *** (48.4–170.0) | |
| Mean length ± SD (range) | 19.2 ± 3.1 (15.0–26.6) | 17.7 ± 1.1 (15.0–19.6) | 22.8 ± 2.0 *** (18.5–26.6) | |
| Condition factor ± SD (range) | 0.72 ± 0.11 (0.56–1.11) | 0.67 ± 0.07 (0.56–0.94) | 0.81 ± 0.11 *** (0.60–1.11) | |
| Parasites | ||||
| Ascaridoids | Prevalence (%) CI 95% | 12.7 0.08–0.20 | 5.7 1.6–14.0 | 25.0 ** 12.7–41.2 |
| Mean intensity (range) CI 95% | 1.07 (1–2) 1.00–1.21 | 1 (1) uncertain | 1.10 (1–2) ns 1.00–1.30 | |
| Mean abundance CI 95% | 0.14 0.07–0.21 | 0.06 0.01–0.11 | 0.28 * 0.13–0.43 | |
| Anisakis spp. | Prevalence (%) CI 95% | 4.5 0.02–0.10 | 0 0.0–5.1 | 12.5 ** 4.2–26.8 |
| Mean intensity (range) CI 95% | 1 uncertain | 0 nd | 1 (1) ns uncertain | |
| Mean abundance CI 95% | 0.05 0.01–0.08 | 0 uncertain | 0.13 * 0.03–0.22 | |
| Hysterothylacium spp. | Prevalence (%) CI 95% | 8.2 0.04–0.15 | 5.7 1.6–14.0 | 12.5 ns 4.2–26.8 |
| Mean intensity (range) CI 95% | 1.11 (1–2) 1.00–1.33 | 1 (1) uncertain | 1.2 (1–2) ns 1.00–1.40 | |
| Mean abundance CI 95% | 0.09 0.04–0.15 | 0.06 0.01–0.11 | 0.15 ns 0.05–0.30 |
| Length (cm) | <17.0 | 17.0–19.9 | ≥20.0 | |
|---|---|---|---|---|
| Parasites | Fish number (F) | 29 | 45 | 36 |
| Ascaridoids | Prevalence (%) ns CI 95% | 3.4 0.1–17.8 | 8.9 2.5–21.2 | 25.0 12.1–42.2 |
| MI (range) CI 95% | 1 uncertain | 1 uncertain | 1.11 (1–2) 1.00–1.33 | |
| MA CI 95% | 0.03 0.00–0.10 | 0.09 0.02–0.18 | 0.28 0.11–0.44 | |
| Anisakis spp. | Prevalence (%) * CI 95% | 0 0.0–11.9 | 0 0.0–7.9 | 13.9 4.7–29.5 |
| MI CI 95% | 0 | 0 | 1 uncertain | |
| MA CI 95% | 0 | 0 | 0.14 0.03–0.25 | |
| Hysterothylacium spp. | Prevalence (%) ns CI 95% | 3.4 0.1–17.8 | 8.9 2.5–21.2 | 11.1 3.1–26.1 |
| MI (range) CI 95% | 1 uncertain | 1 uncertain | 1.25 (1–2) 1.00–1.50 | |
| MA CI 95% | 0.03 0.00–0.10 | 0.09 0.02–0.18 | 0.14 0.03–0.31 |
| FAO Area | Parameters | Ascaridoids # | Hysterothylacium spp. | Anisakis spp. | |
|---|---|---|---|---|---|
| 27.IXa, NE Atlantic, coast of SW Spain | Fish weight ± SD | U I | 33.7 ± 6.2 42.2 ± 12.0 ns | 33.7 ± 6.2 42.2 ± 12.0 ns | ND |
| Fish length ± SD | U I | 17.1 ± 1.0 18.5 ± 1.4 ** | 17.1 ± 1.0 18.5 ± 1.4 ** | ND | |
| Condition factor ± SD | U I | 0.67 ± 0.07 0.65 ± 0.06 ns | 0.67 ± 0.07 0.65 ± 0.06 ns | ND | |
| 34.1.11, CE Atlantic, coast of NW Morocco | Fish weight ± SD | U I | 96.2 ± 33.5 109.1 ± 37.2 ns | 100.7 ± 34.6 90.8 ± 35.6 ns | 95.4 ± 33.3 127.3 ± 32.0 * |
| Fish length ± SD | U I | 22.5 ± 1.9 23.5 ± 2.3 ns | 22.9 ± 1.9 22.3 ± 2.6 ns | 22.5 ± 1.9 24.8 ± 1.2 ** | |
| Condition factor ± SD | U I | 0.81 ± 0.13 0.81 ± 0.12 ns | 0.81 ± 0.13 0.78 ± 0.11 ns | 0.81 ± 0.13 0.83 ± 0.13 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buzo-Domínguez, S.; Morales-Yuste, M.; Domingo-Hernández, A.M.; Benítez, R.; Adroher, F.J. Molecular Epidemiology of Anisakis spp. in Wedge Sole, Dicologlossa cuneata (Moreau, 1881), from Fishmarkets in Granada (Southern Spain), Caught in Two Adjacent NE and CE Atlantic Areas. Pathogens 2021, 10, 1302. https://doi.org/10.3390/pathogens10101302
Buzo-Domínguez S, Morales-Yuste M, Domingo-Hernández AM, Benítez R, Adroher FJ. Molecular Epidemiology of Anisakis spp. in Wedge Sole, Dicologlossa cuneata (Moreau, 1881), from Fishmarkets in Granada (Southern Spain), Caught in Two Adjacent NE and CE Atlantic Areas. Pathogens. 2021; 10(10):1302. https://doi.org/10.3390/pathogens10101302
Chicago/Turabian StyleBuzo-Domínguez, Sara, Manuel Morales-Yuste, Ana María Domingo-Hernández, Rocío Benítez, and Francisco Javier Adroher. 2021. "Molecular Epidemiology of Anisakis spp. in Wedge Sole, Dicologlossa cuneata (Moreau, 1881), from Fishmarkets in Granada (Southern Spain), Caught in Two Adjacent NE and CE Atlantic Areas" Pathogens 10, no. 10: 1302. https://doi.org/10.3390/pathogens10101302