Emergence of the New KPC-49 Variant Conferring an ESBL Phenotype with Resistance to Ceftazidime-Avibactam in the ST131-H30R1 Escherichia coli High-Risk Clone

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
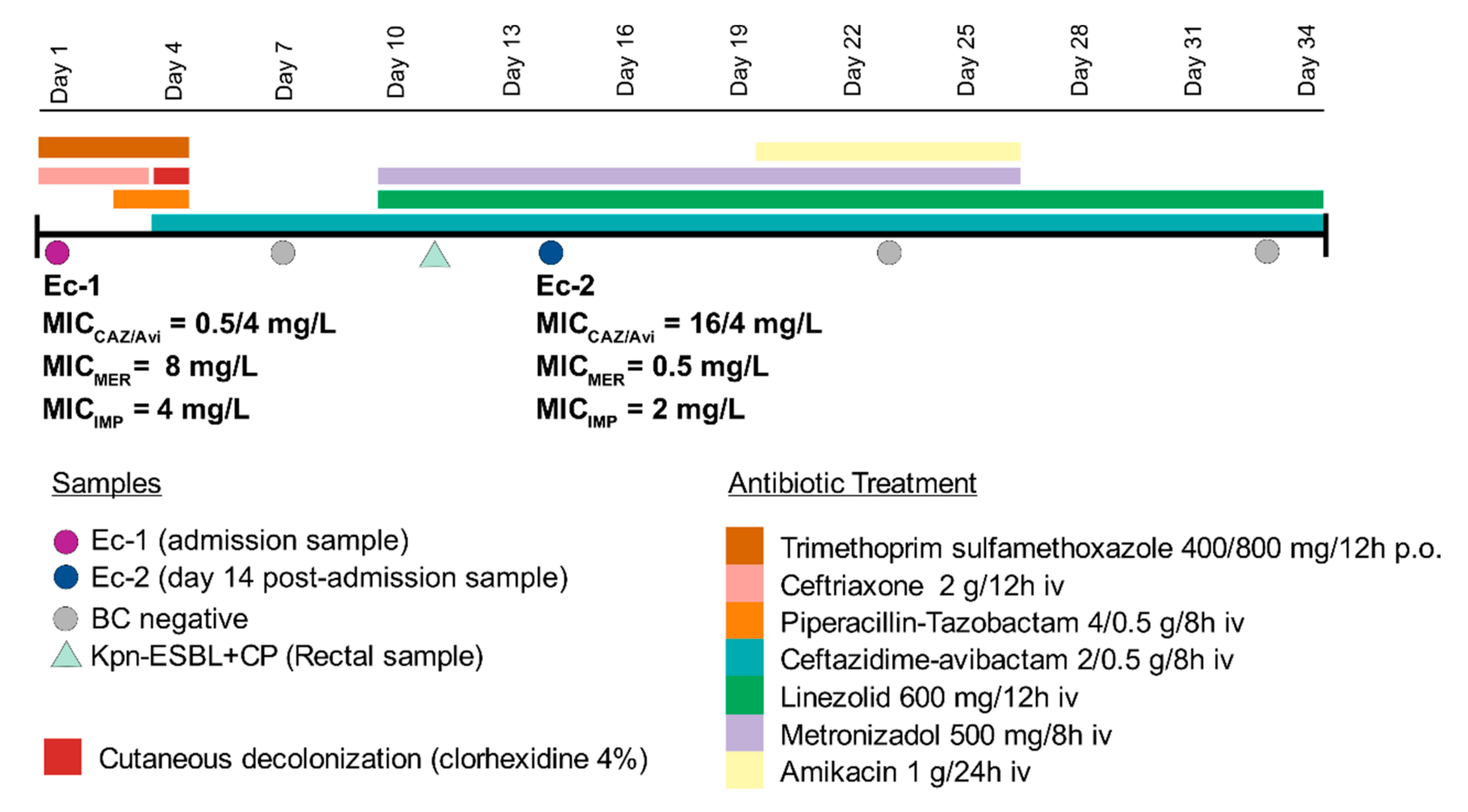
2.1. Case Report
2.2. Genome Characteristics and E. coli Typing
2.3. Resistance and Virulence Profile
2.4. blaKPC Genetic Environment
2.5. Resistance Phenotype Conferred by blaKPC-49 Gene
3. Discussion
4. Material and Methods
4.1. Bacterial Isolates, Susceptibility Testing, and Patient’s Data
4.2. Resistance Genes Characterization
4.3. DNA Extraction, Whole Genome Sequencing, and Bioinformatics Analysis
4.4. Validation of blaKPC-49 Variant
4.5. Sequence Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Ethics Committee Approval
Conflicts of Interest
References
- Johnson, J.R.; Johnston, B.; Clabots, C.; Kuskowski, M.A.; Castanheira, M. Escherichia coli Sequence Type ST131 as the Major Cause of Serious Multidrug-Resistant, E. coli Infections in the United States. Clin. Infect. Dis. 2010, 51, 286–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Menard, M.; Johnston, B.; Kuskowski, M.A.; Nichol, K.; Zhanel, G.G. Epidemic clonal groups of Escherichia coli as a cause of antimicrobial-resistant urinary tract infections in Canada, 2002 to 2004. Antimicrob. Agents Chemother. 2009, 53, 2733–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolas-Chanoine, M.H.; Bertrand, X.; Madec, J.Y. Escherichia coli st131, an intriguing clonal group. Clin. Microbiol. Rev. 2014, 27, 543–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Tchesnokova, V.; Johnston, B.; Clabots, C.; Roberts, P.L.; Billig, M.; Riddell, K.; Rogers, P.; Qin, X.; Butler-Wu, S.; et al. Abrupt emergence of a single dominant multidrug-resistant strain of Escherichia coli. J. Infect. Dis. 2013, 207, 919–928. [Google Scholar] [CrossRef]
- Lance, B.P.; Johnson, J.R.; Aziz, M.; Clabots, C.; Johnston, B.; Tchesnokova, V.; Nordstrom, L.; Billig, M.; Chattopadhyay, S.; Stegger, M.; et al. The Epidemic of extended-spectrum-beta-lactamase-producing Escherichia coli ST131 is driven by a single highly pathogenic subclone, H30-Rx. Am. Soc. Microbiol. 2013, 4, 1–10. [Google Scholar] [CrossRef]
- Merino, I.; Hernández-García, M.; Turrientes, M.C.; Pérez-Viso, B.; López-Fresneña, N.; Díaz Agero, C.; Maechler, F.; Fankhauser-Rodriguez, C.; Kola, A.; Schrenzel, J.; et al. Emergence of ESBL-E. coli-ST131-C1-M27 clade colonizing patients in Europe. J. Antimicrob. Chemother. 2018, 73, 2973–2980. [Google Scholar] [CrossRef]
- Ortega, A.; Sáez, D.; Bautista, V.; Fernández-Romero, B.; Lara, N.; Aracil, B.; Pérez-Vázquez, M.; Campos, J.; Oteo, J. Carbapenemase-producing Escherichia coli is becoming more prevalent in Spain mainly because of the polyclonal dissemination of OXA-48. J. Antimicrob. Chemother. 2016, 71, 2131–2138. [Google Scholar] [CrossRef] [Green Version]
- Naas, T.; Cuzon, G.; Villegas, M.V.; Lartigue, M.F.; Quinn, J.P.; Nordmann, P. Genetic structures at the origin of acquisition of the β-lactamase blaKPC gene. Antimicrob. Agents Chemother. 2008, 52, 1257–1263. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Price, S.; Poirel, L.; Bonomo, R.; Schwaber, M.; Daikos, G.; Cormican, M.; Cornaglia, G.; Garau, J. Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect. Dis. 2013, 13, 785–796. [Google Scholar] [CrossRef] [Green Version]
- Suay-García, B.; Pérez-Gracia, M.T. Present and future of carbapenem-resistant Enterobacteriaceae (CRE) infections. Antibiotics 2019, 8, 122. [Google Scholar] [CrossRef] [Green Version]
- Carmeli, Y.; Armstrong, J.; Laud, P.J.; Newell, P.; Stone, G.; Wardman, A.; Gasink, L.B. Ceftazidime-avibactam or best available therapy in patients with ceftazidime-resistant Enterobacteriaceae and Pseudomonas aeruginosa complicated urinary tract infections or complicated intra-abdominal infections (REPRISE): A randomised, pathogen-directed. Lancet Infect. Dis. 2016, 16, 661–673. [Google Scholar] [CrossRef]
- Wu, G.; Abraham, T.; Lee, S. Ceftazidime-avibactam for treatment of carbapenem-resistant enterobacteriaceae bacteremia. Clin. Infect. Dis. 2016, 63, 1147–1148. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, A.; Caio, C.; Mezzatesta, M.L.; Rifici, C.; Bramanti, P.; Stefani, S.; Mazzon, E. Successful ceftazidime-avibactam treatment of MDR-KPC-positive Klebsiella pneumoniae infection in a patient with traumatic brain injury. Medicine 2017, 96, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Van Duin, D.; Lok, J.J.; Earley, M.; Cober, E.; Richter, S.S.; Perez, F.; Salata, R.A.; Kalayjian, R.C.; Watkins, R.R.; Doi, Y.; et al. Colistin versus veftazidime-avibactam in the treatment of infections due to carbapenem-resistant Enterobacteriaceae. Clin. Infect. Dis. 2018, 66, 163–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, B.D.; Thuras, P.; Porter, S.B.; Anacker, M.; VonBank, B.; Vagnone, P.S.; Witwer, M.; Castanheira, M.; Johnson, J.R. Activity of cefiderocol, ceftazidime-avibactam, and eravacycline against carbapenem-resistant Escherichia coli isolates from the United States and international sites in relation to clonal background, resistance genes, coresistance, and region. Antimicrob. Agent Chemother. 2020, 64, 1–10. [Google Scholar] [CrossRef]
- Shields, R.K.; Chen, L.; Cheng, S.; Chavda, K.D.; Press, E.G. Emergence of ceftazidime-avibactam mutations during treatment of carbapenem-resistant Klebsiella pneumoniae Infections. Antimicrob. Agents Chemother. 2016, 61, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Haidar, G.; Clancy, J.C.; Shields, R.K.; Hao, B.C.; Cheng, S.; Nguyen, M.H. Mutations in blaKPC-3 That confer ceftazidime-avibactam resistance encode novel KPC-3 variants that function as extended-spectrum β-lactamases. Antimicrob. Agents Chemother. 2017, 61, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giddins, M.J.; Macesic, N.; Medini, K.A.; Stump, S.; Khan, S.; McConville, T.H.; Mehta, M.; Gomez-Simmonds, A.; Uhlemann, A.-C. Successive emergence of ceftazidime-avibactam resistance through distinct genomic adaptations in blaKPC-2-harboring Klebsiella pneumoniae sequence type 307 Isolates. Antimicrob. Agents Chemother. 2018, 62, 1–8. [Google Scholar]
- Plan de Prevención y control frente a la infección por EPC en la Comunidad de Madrid. Versión 1-September 2013. Available online: https://www.comunidad.madrid/servicios/salud/prevencion-control-infecciones-epc (accessed on 13 January 2021).
- Ripabelli, G.; Sammarco, M.L.; Scutellà, M.; Felice, V.; Tamburro, M. Carbapenem-resistant KPC- and TEM-producing Escherichia coli ST131 isolated from a hospitalized patient with urinary tract infection: First isolation in molise region, central Italy, July 2018. Microb. Drug Resist. 2019, 26. [Google Scholar] [CrossRef]
- Piazza, A.; Caltagirone, M.; Bitar, I.; Nucleo, E.; Spalla, M.; Fogato, E.; D’Angelo, R.; Pagani, L.R.M. Emergence of Escherichia coli sequence type 131 (ST131) and ST3948 with KPC-2, KPC-3 and KPC-8 carbapenemases from a long-term care and rehabilitation Facility (LTCRF) in Northern Italy. Adv. Exp. Med. Biol. 2015. [Google Scholar] [CrossRef]
- Hernández-García, M.; Pérez-Viso, B.; Navarro-San Francisco, C.; Baquero, F.; Morosini, M.I.; Ruiz-Garbajosa, P.; Cantón, R. Intestinal co-colonization with different carbapenemase-producing Enterobacterales isolates is not a rare event in an OXA-48 endemic area. EClinicalMedicine 2019, 15, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Baquero, F.; Coque, T.M.; Martínez, J.L.; Aracil-Gisbert, S.; Lanza, V.F. Gene transmission in the one health microbiosphere and the channels of antimicrobial resistance. Front. Microbiol. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curiao, T.; Morosini, M.I.; Ruiz-Garbajosa, P.; Robustillo, A.; Baquero, F.; Coque, T.M.; Cantón, R. Emergence of blaKPC-3-Tn4401a associated with a pKPN3/4-like plasmid within ST384 and ST388 Klebsiella pneumoniae clones in Spain. J. Antimicrob. Chemother. 2010, 65, 1608–1614. [Google Scholar] [CrossRef] [PubMed]
- Gona, F.; Barbera, F.; Pasquariello, A.C.; Grossi, P.; Gridelli, B.; Mezzatesta, M.L.; Caio, C.; Stefani, S.; Conaldi, P.G. In Vivo multiclonal transfer of blaKPC-3 from Klebsiella pneumoniae to Escherichia coli in surgery patients. Clin. Microbiol. Infect. 2014, 20, 633–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanamori, H.; Parobek, C.M.; Juliano, J.J.; Johnson, J.R.; Johnston, B.D.; Johnson, T.J.; Weber, D.J.; Rutala, W.A.; Anderson, D.J. Genomic analysis of multidrug-resistant Escherichia coli from North Carolina community hospitals: Ongoing. Antimicrob. Agents Chemother. 2017, 61, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, T.J.; Danzeisen, J.L.; Youmans, B.; Case, K.; Llop, K.; Munoz-aguayo, J.; Flores-figueroa, C.; Aziz, M.; Stoesser, N.; Sokurenko, E.; et al. Separate F-type plasmids have shaped the evolution of the H 30 subclone of. mSphere 2016, 1, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoesser, N.; Sheppard, A.E.; Pankhurst, L.; de Maio, N.; Moore, C.E.; Sebra, R.; Turner, P.; Anson, L.W.; Kasarskis, A.; Batty, E.M.; et al. Evolutionary history of the global emergence of the Escherichia coli epidemic clone ST131. MBio 2016, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Díaz, M.A.; Hernández-Bello, J.R.; Rodríguez-Baño, J.; Martínez-Martínez, L.; Calvo, J.; Blanco, J.; Pascual, A. Diversity of Escherichia coli strains producing extended-spectrum β-lactamases in Spain: Second nationwide study. J. Clin. Microbiol. 2010, 48, 2840–2845. [Google Scholar] [CrossRef] [Green Version]
- Humphries, R.M.; Hemarajata, P. Resistance to ceftazidime-Avibactam in Klebsiella pneumoniae due to porin mutations & the increased expression of KPC-3. Antimicrob. Agents Chemother. 2017, 61, 10–11. [Google Scholar] [CrossRef] [Green Version]
- Nelson, K.; Hemarajata, P.; Sun, D.; Rubio-Aparicio, D.; Tsivkovski, R.; Yang, S.; Sebra, R.; Kasarskis, A.; Nguyen, H.; Hanson, B.M.; et al. Resistance to Ceftazidime–Avibactam is due to transposition of KPC in a Porin–Deficient strain of Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2017, 61, e00989-17. [Google Scholar] [CrossRef] [Green Version]
- García-Fernández, S.; Morosini, M.I.; Marco, F.; Gijón, D.; Vergara, A.; Vila, J.; Ruiz-Garbajosa, P.; Cantón, R. Evaluation of the eazyplex® SuperBug CRE system for rapid detection of carbapenemases and ESBLs in clinical Enterobacteriaceae isolates recovered at two Spanish hospitals. J. Antimicrob. Chemother. 2014, 70, 1047–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.O.N.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Beghain, J.; Bridier-Nahmias, A.; Nagard, H.L.; Denamur, E.; Clermont, O. ClermonTyping: An easy-to-use and accurate in silico method for Escherichia genus strain phylotyping. Microb. Genom. 2018, 4, 1–8. [Google Scholar] [CrossRef]
- Banerjee, R.; Robicsek, A.; Kuskowski, M.A.; Porter, S.; Johnston, B.D.; Sokurenko, E.; Tchesnokova, V.; Price, L.B.; Johnson, J.R. Molecular epidemiology of Escherichia coli sequence type 131 and its H30 and H30-Rx subclones among extended-spectrum-β-lactamase-positive and -negative E. coli clinical isolates from the Chicago region, 2007 to 2010. Antimicrob. Agents Chemother. 2013, 57, 6385–6388. [Google Scholar] [CrossRef] [Green Version]
- Antipov, D.; Hartwick, N.; Shen, M.; Raiko, M.; Lapidus, A.; Pevzner, P.A. Plasmidspades: Assembling plasmids from whole genome sequencing data. Bioinformatics 2016, 32, 3380–3387. [Google Scholar] [CrossRef] [Green Version]
- Ardanuy, C.; Liñares, J.; Domínguez, M.A.; Hernández-Allés, S.; Benedí, V.J.; Martínez-Martínez, L. Outer membrane profiles of clonally related Klebsiella pneumoniae isolates from clinical samples and activities of cephalosporins and carbapenems. Antimicrob. Agents Chemother. 1998, 42, 1636–1640. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Broth Microdilution (mg/L) (Interpretation) | Gradient Strips (mg/L) (Interpretation) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Antimicrobials | Ec-1 | Ec-2 | E. coli pKPC-3-TM | E. coli pKPC-49-TM | E. coli DH5α | K. pneumoniae pKPC-3-TM | K. pneumoniae pKPC-49-TM | K. pneumoniae CSUB10R | E. coli pKPC-3-TM | E. coli pKPC-49-TM | E. coli DH5α | K. pneumoniae pKPC-3-TM | K. pneumoniae pKPC-49-TM | K. pneumoniae CSUB10R |
| Piperacillin-tazobactam | >64/4 (R) | 16/4 (I) | ≤4/4 (S) | ≤4/4 (S) | ≤4/4 (S) | >64/4 (R) | >64/4 (R) | >64/4 (R) | - | - | - | - | - | - |
| Ceftazidime | >32 (R) | >32 (R) | 1 (S) | 2 (I) | ≤0.25 (S) | >32 (R) | >32 (R) | >32 (R) | 0.25 (S) | 1 (S) | 0.19 (S) | >256 (R) | >256 (R) | >256 (R) |
| Cefotaxime | >32 (R) | >32 (R) | ≤0.25 (S) | ≤0.25 (S) | ≤0.25 (S) | >32 (R) | >32 (R) | >32 (R) | 0.032 (S) | 0.047 (S) | 0.032 (S) | >256 (R) | >256 (R) | >256 (R) |
| Cefepime | >64 (R) | 64 (R) | ≤0.125 (S) | 0.25 (S) | ≤0.125 (S) | >64 (R) | >64 (R) | 64 (R) | 0.047 (S) | 0.047 (S) | 0.023 (S) | >256 (R) | >256 (R) | 32 (R) |
| Aztreonam | >32 (R) | >32 (R) | 4 (I) | ≤0.5 (S) | ≤0.5 (S) | >32 (R) | >32 (R) | >32 (R) | - | - | - | - | - | - |
| Ceftazidime-avibactam | 0.5/4 (S) | 16/4 (R) | ≤0.125/4 (S) | ≤0.125/4 (S) | ≤0.125/4 (S) | 4/4 (S) | 4/4 (S) | 2/4 (S) | 0.047/4 (S) | 0.094/4 (S) | 0.094/4 (S) | 2/4 (S) | 12/4 (R) | 2/4 (S) |
| Imipenem | 4 (I) | 2 (S) | 0.5 (S) | ≤0.25 (S) | ≤0.25 (S) | >16 (R) | >16 (R) | 2 (S) | 0.25 (S) | 0.25 (S) | 0.38 (S) | >32 (R) | 4 (I) | 0.75 (S) |
| Meropenem | 8 (I) | 0.5 (S) | ≤0.125 (S) | ≤0.125 (S) | ≤0.125 (S) | 16 (R) | 8 (I) | 2 (S) | 0.023 (S) | 0.023 (S) | 0.032 (S) | 6 (I) | 2 (S) | 2 (S) |
| Gentamicin | >8 (R) | >8 (R) | ≤8 (S) | ≤8 (S) | ≤8 (S) | ≤8 (S) | ≤8 (S) | ≤8 (S) | - | - | - | - | - | - |
| Amikacin | ≤8 (S) | ≤8 (S) | ≤8 (S) | ≤8 (S) | ≤8 (S) | ≤8 (S) | ≤8 (S) | ≤8 (S) | - | - | - | - | - | - |
| Tobramycin | >8 (R) | >8 (R) | ≤8 (S) | ≤8 (S) | ≤8 (S) | ≤8 (S) | ≤8 (S) | ≤8 (S) | - | - | - | - | - | - |
| Ciprofloxacin | >2 (R) | >2 (R) | ≤2 (S) | ≤2 (S) | ≤2 (S) | >2 (R) | >2 (R) | >2 (R) | - | - | - | - | - | - |
| Tigecycline | ≤0.5 (S) | ≤0.5 (S) | ≤0.5 (S) | ≤0.5 (S) | ≤0.5 (S) | ≤0.5 (S) | ≤0.5 (S) | ≤0.5 (S) | - | - | - | - | - | - |
| Colistin | ≤2 (S) | ≤2 (S) | ≤2 (S) | ≤2 (S) | ≤2 (S) | ≤2 (S) | ≤2 (S) | ≤2 (S) | - | - | - | - | - | - |
| Fosfomycin | ≤32 (S) | ≤32 (S) | ≤32 (S) | ≤32 (S) | ≤32 (S) | >32 (S) | >32 (S) | >32 (S) | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-García, M.; Sánchez-López, J.; Martínez-García, L.; Becerra-Aparicio, F.; Morosini, M.I.; Ruiz-Garbajosa, P.; Cantón, R. Emergence of the New KPC-49 Variant Conferring an ESBL Phenotype with Resistance to Ceftazidime-Avibactam in the ST131-H30R1 Escherichia coli High-Risk Clone. Pathogens 2021, 10, 67. https://doi.org/10.3390/pathogens10010067
Hernández-García M, Sánchez-López J, Martínez-García L, Becerra-Aparicio F, Morosini MI, Ruiz-Garbajosa P, Cantón R. Emergence of the New KPC-49 Variant Conferring an ESBL Phenotype with Resistance to Ceftazidime-Avibactam in the ST131-H30R1 Escherichia coli High-Risk Clone. Pathogens. 2021; 10(1):67. https://doi.org/10.3390/pathogens10010067
Chicago/Turabian StyleHernández-García, Marta, Javier Sánchez-López, Laura Martínez-García, Federico Becerra-Aparicio, María Isabel Morosini, Patricia Ruiz-Garbajosa, and Rafael Cantón. 2021. "Emergence of the New KPC-49 Variant Conferring an ESBL Phenotype with Resistance to Ceftazidime-Avibactam in the ST131-H30R1 Escherichia coli High-Risk Clone" Pathogens 10, no. 1: 67. https://doi.org/10.3390/pathogens10010067