Cryptic Diversity in Colombian Edible Leaf-Cutting Ants (Hymenoptera: Formicidae)

 , ,
, , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. DNA Extraction and PCR Amplification
2.3. Sequence Alignment
2.4. Phylogenetic Analyses
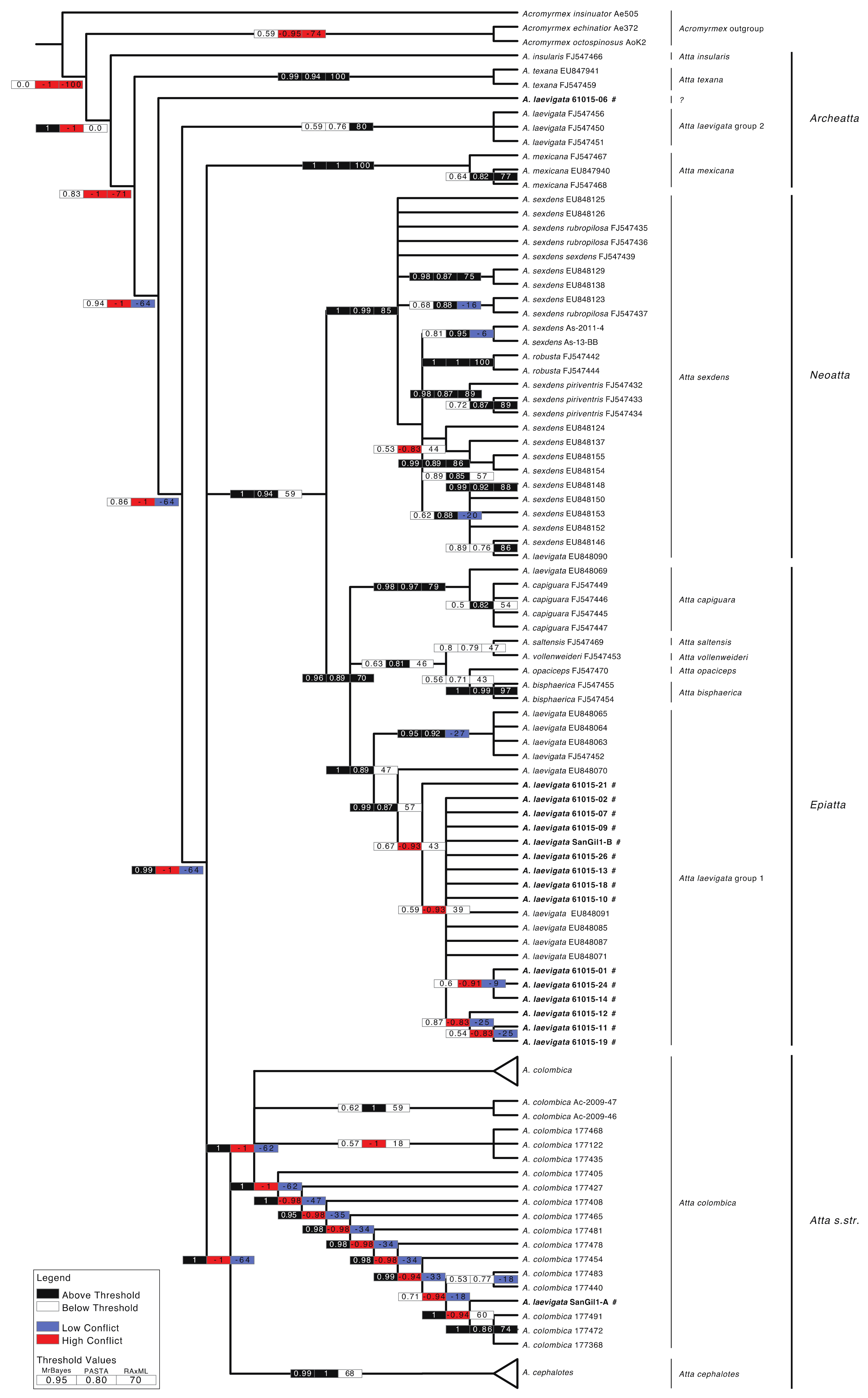
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weinbaum, K.Z.; Brashares, J.S.; Golden, C.D.; Getz, W.M. Searching for sustainability: Are assessments of wildlife harvests behind the times? Ecol. Lett. 2013, 16, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Marko, P.B.; Nance, H.A.; Guynn, K.D. Genetic detection of mislabeled fish from a certified sustainable fishery. Curr. Biol. 2011, 21, R621–R622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helyar, S.J.; Lloyd, H.A.D.; de Bruyn, M.; Leake, J.; Bennett, N.; Carvalho, G.R. Fish product mislabelling: Failings of traceability in the production chain and implications for illegal, unreported and unregulated (IUU) fishing. PLoS ONE 2014, 9, e98691. [Google Scholar] [CrossRef] [PubMed]
- Dentinger, B.T.M.; Suz, L.M. What’s for dinner? Undescribed species of porcini in a commercial packet. PeerJ 2014, 2, e570. [Google Scholar] [CrossRef]
- Sodhi, N.S.; Brook, B.W.; Bradshaw, C.J.A. Causes and consequences of species extinctions. In The Princeton Guide to Ecology; Levin, S.A., Carpenter, S.R., Godfray, H.C.J., Kinzig, A.P., Loreau, M., Losos, J.B., Walker, B., Wilcove, D.S., Eds.; Princeton University Press: Princeton, NJ, USA, 2009; pp. 514–520. [Google Scholar]
- Essington, T.E.; Moriarty, P.E.; Froehlich, H.E.; Hodgson, E.E.; Koehn, L.E.; Oken, K.L.; Siple, M.C.; Stawitz, C.C. Fishing amplifies forage fish population collapses. Proc. Natl. Acad. Sci. USA 2015, 112, 6648–6652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raubenheimer, D.; Rothman, J.M. Nutritional ecology of entomophagy in humans and other primates. Annu. Rev. Entomol. 2013, 58, 141–160. [Google Scholar] [CrossRef]
- Bolton, B. An Online Catalog of the Ants of the World. Available online: http://www.antcat.org/ (accessed on 22 October 2018).
- Weber, N.A. Fungus-growing ants. Science 1966, 153, 587–604. [Google Scholar] [CrossRef]
- Schultz, T.R.; Brady, S.G. Major evolutionary transitions in ant agriculture. Proc. Natl. Acad. Sci. USA 2008, 105, 5435–5440. [Google Scholar] [CrossRef] [Green Version]
- Nygaard, S.; Hu, H.; Li, C.; Schiøtt, M.; Chen, Z.; Yang, Z.; Xie, Q.; Ma, C.; Deng, Y.; Dikow, R.B.; et al. Reciprocal genomic evolution in the ant-fungus agricultural symbiosis. Nat. Commun. 2016, 7, 12233. [Google Scholar] [CrossRef]
- Branstetter, M.G.; Jesovnik, A.; Sosa-Calvo, J.; Lloyd, M.W.; Faircloth, B.C.; Brady, S.G.; Schultz, T.R. Dry habitats were crucibles of domestication in the evolution of agriculture in ants. Proc. R. Soc. Lond. B Biol. Sci. 2017, 284. [Google Scholar] [CrossRef]
- Hölldobler, B.; Wilson, E.O. The Leafcutter Ants; W. W. Norton & Company, Inc.: New York, NY, USA; London, UK, 2011; pp. 1–160. [Google Scholar]
- Fowler, H.G.; Pagani, M.I.; Da Silva, O.A.; Forti, L.C.; Da Silva, V.P.; De Vasconcelos, H.L. A pest is a pest is a pest? The dilemma of Neotropical leaf-cutting ants: Keystone taxa of natural ecosystems. Environ. Manag. 1989, 13, 671–675. [Google Scholar] [CrossRef]
- Leal, I.R.; Wirth, R.; Tabarelli, M. The multiple impacts of leaf-cutting ants and their novel ecological role in human-modified neotropical forests. Biotropica 2014, 46, 516–528. [Google Scholar] [CrossRef]
- Gonçalves, C.R. Contribuicão para o conhecimento do gênero Atta Fabr., das formigas saúvas. Boletim da Sociedade Brasileira de Agronomia 1942, 5, 333–358. [Google Scholar]
- Borgmeier, T. Estudos sôbre Atta (Hym. Formicidae). Mem I Oswaldo Cruz 1950, 48, 239–292. [Google Scholar] [CrossRef]
- Borgmeier, T. Revision der Gattung Atta Fabricius (Hymenoptera, Formicidae). Stud. Entomol. 1959, 2, 321–390. [Google Scholar]
- Bacci, M., Jr.; Solomon, S.E.; Mueller, U.G.; Martins, V.G.; Carvalho, A.O.R.; Vieira, L.G.E.; da Silva-Pinhati, A.C.O. Phylogeny of leafcutter ants in the genus Atta Fabricius (Formicidae: Attini) based on mitochondrial and nuclear DNA sequences. Mol. Phylogenet. Evol. 2009, 51, 427–437. [Google Scholar] [CrossRef]
- Bukkens, S.G.F. The nutritional value of edible insects. Ecol. Food Nutr. 1997, 36, 287–319. [Google Scholar] [CrossRef]
- Paoletti, M.G. Ecological Implications Minilivestock; Paoletti, M.G., Ed.; Science Publishers, Inc.: Enfield, NH, USA, 2005; pp. 1–677. [Google Scholar]
- Ramos-Elorduy, J. The importance of edible insects in the nutrition and economy of people of the rural areas of Mexico. Ecol. Food Nutr. 1997, 36, 347–366. [Google Scholar] [CrossRef]
- Ramos-Elorduy, J.; Pino Moreno, J.M. Edible insects of Chiapas, Mexico. Ecol. Food Nutr. 2002, 41, 271–299. [Google Scholar] [CrossRef]
- Onore, G. A brief note on edible insects in Ecuador. Ecol. Food Nutr. 1997, 36, 277–285. [Google Scholar] [CrossRef]
- Dufour, D.L. Insects as food: A case study from the Northwest Amazon. Am. Anthropol. 1987, 89, 383–397. [Google Scholar] [CrossRef]
- Ramos-Elorduy, J. Creepy Crawly Cuisine; Ramos-Elorduy, J., Ed.; Park Street Press: Rochester, VT, USA, 1998; 160p. [Google Scholar]
- Ramos-Elorduy, J. Insects: A sustainable source of food? Ecol. Food Nutr. 1997, 36, 247–276. [Google Scholar] [CrossRef]
- Frederickson, M.E. The reproductive phenology of an Amazonian ant species reflects the seasonal availability of its nest sites. Oecologia 2006, 149, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Baer, B.C.; Armitage, S.A.O.; Boomsma, J.J. Sperm storage induces an immunity cost in ants. Nature 2006, 441, 872–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sternberg, L.D.S.L.; Pinzon, M.C.; Moreira, M.Z.; Moutinho, P.; Rojas, E.I.; Herre, E.A. Plants use macronutrients accumulated in leaf-cutting ant nests. Proc. R. Soc. B 2007, 274, 315–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Hallwachs, W.; Janzen, D.H. Diversity and phylogenetic community structure of ants along a Costa Rican elevational gradient. Ecography 2014, 37, 720–731. [Google Scholar] [CrossRef] [Green Version]
- Solomon, S.E.; Bacci, M., Jr.; Martins, J., Jr.; Vinha, G.G.; Mueller, U.G. Paleodistributions and comparative molecular phylogeography of leafcutter ants (Atta spp.) provide new insight into the origins of Amazonian diversity. PLoS ONE 2008, 3, e2738. [Google Scholar] [CrossRef]
- Walsh, P.S.; Metzger, D.A.; Higuchi, R. Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques 1991, 10, 506–513. [Google Scholar] [CrossRef]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Moreau, C.S.; Bell, C.D.; Vila, R.; Archibald, S.B. Phylogeny of the ants: Diversification in the age of angiosperms. Science 2006, 312, 101–104. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Nygaard, S.; Zhang, G.; Schiøtt, M.; Li, C.; Wurm, Y.; Hu, H.; Zhou, J.; Ji, L.; Qiu, F.; Rasmussen, M.; et al. The genome of the leaf-cutting ant Acromyrmex echinatior suggests key adaptations to advanced social life and fungus farming. Genome Res. 2011, 21, 1339–1348. [Google Scholar] [CrossRef] [PubMed]
- Suen, G.; Teiling, C.; Li, L.; Holt, C.; Abouheif, E.; Bornberg-Bauer, E.; Bouffard, P.; Caldera, E.J.; Cash, E.; Cavanaugh, A.; et al. The genome sequence of the leaf-cutter ant Atta cephalotes reveals insights into its obligate symbiotic lifestyle. PLoS Genet. 2011, 7, e1002007. [Google Scholar] [CrossRef]
- Scientific, T.F. BigDye Terminator v3.1 Cycle Sequencing Kit User Guide (Pub. no. 4337035 Rev C). 2016, pp. 1–50. Available online: https://tools.thermofisher.com/content/sfs/manuals/cms_081527.pdf (accessed on 11 December 2018).
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Mirarab, S.; Nguyen, N.; Guo, S.; Wang, L.-S.; Kim, J.; Warnow, T. PASTA: Ultra-large multiple sequence alignment for nucleotide and amino-acid sequences. J. Comput. Biol. 2015, 22, 377–386. [Google Scholar] [CrossRef]
- Mirarab, S.; Warnow, T. FastSP: Linear time calculation of alignment accuracy. Bioinformatics 2011, 27, 3250–3258. [Google Scholar] [CrossRef]
- Crozier, R.H.; Crozier, Y.C. The mitochondrial genome of the honeybee Apis mellifera: Complete sequence and genome organization. Genetics 1993, 133, 97–117. [Google Scholar]
- Piel, W.H.; Auman, J.; Chan, L.; Dominus, M.J.; Gapeyev, V.; Guo, Y.; Lapp, H.; Ruan, J.; Tannen, V.; Vos, R.A. TreeBASE v. 2: A Database of Phylogenetic Knowledge. Available online: http://purl.org/phylo/treebase/phylows/study/TB2:S23442 (accessed on 11 December 2018).
- Nixon, K.C. WinClada 2002. Available online: http://evolution.genetics.washington.edu/phylip/software.pars.html (accessed on 11 December 2018).
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, F.; Oliver, J.L.; Marín, A.; Medina, J.R. The general stochastic model of nucleotide substitution. J. Theor. Biol. 1990, 142, 485–501. [Google Scholar] [CrossRef]
- Rambaut, A.; Suchard, M.A.; Xie, W.; Drummond, A.J. Tracer v 1.6 2014. Available online: http://beast.bio.ed.ac.uk/ (accessed on 11 December 2018).
- Huelsenbeck, J.P.; Ronquist, F.; Nielsen, R.; Bollback, J.P. Bayesian inference of phylogeny and Its impact on evolutionary biology. Science 2001, 294, 2310–2314. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.A.; Wilgenbusch, J.C.; Warren, D.L.; Swofford, D.L. AWTY (are we there yet?): A system for graphical exploration of MCMC convergence in Bayesian phylogenetics. Bioinformatics 2007, 24, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Alfaro, M.E. Bayes or bootstrap? A simulation study comparing the performance of Bayesian Markov chain Monte Carlo sampling and bootstrapping in assessing phylogenetic confidence. Mol. Biol. Evol. 2003, 20, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Lutzoni, F.M.; Kauff, F.; Cox, C.J.; McLaughlin, D.; Celio, G.; Dentinger, B.; Padamsee, M.; Hibbett, D.; James, T.Y.; Baloch, E.; et al. Assembling the fungal tree of life: Progress, classification, and evolution of subcellular traits. Am. J. Bot. 2004, 91, 1446–1480. [Google Scholar] [CrossRef]
- Oliver, J.C.; Miadlikowska, J.; Arnold, A.E.; Maddison, D.R.; Lutzoni, F.M. Hypha: A Mesquite Package for Support Value Integration 2013. Mesquite. Available online: http://mesquiteproject.org/packages/hypha (accessed on 11 December 2018).
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis 2011. Available online: http://mesquiteproject.org (accessed on 11 December 2018).
- Kooij, P.W. Scripts and Files for Hormigas Culonas Paper. Available online: https://github.com/pwkooij/Hormigas_culonas (accessed on 11 December 2018).
- Pergande, T. Mexican Formicidae. Proc. Calif. Acad. Sci. 1895, 5, 858–896. [Google Scholar]
- Fischer, G.; Azorsa, F.; Garcia, F.H.; Mikheyev, A.; Economo, E. Two new phragmotic ant species from Africa: Morphology and next-generation sequencing solve a caste association problem in the genus Carebara Westwood. ZooKeys 2015, 525, 77–105. [Google Scholar] [CrossRef] [PubMed]
- Blaimer, B.B.; Brady, S.G.; Schultz, T.R.; Lloyd, M.W.; Fisher, B.L.; Ward, P.R. Phylogenomic methods outperform traditional multi-locus approaches in resolving deep evolutionary history: A case study of formicine ants. BMC Evol. Biol. 2015, 15, 271. [Google Scholar] [CrossRef] [PubMed]
- Jesovnik, A.; Sosa-Calvo, J.; Lloyd, M.W.; Branstetter, M.G.; Fernández, F.; Schultz, T.R. Phylogenomic species delimitation and host-symbiont coevolution in the fungus-farming ant genus Sericomyrmex Mayr (Hymenoptera: Formicidae): Ultraconserved Elements (UCEs) resolve a recent radiation. Syst. Entomol. 2017, 42, 523–542. [Google Scholar] [CrossRef]
- Staab, M.; Kleineidam, C.J. Initiation of swarming behavior and synchronization of mating flights in the leaf-cutting ant Atta vollenweideri FOREL, 1893 (Hymenoptera: Formicidae). Myrmecol. News 2014, 19, 93–102. [Google Scholar]
- Keller, L.; Passera, L. Size and fat content of gynes in relation to the mode of colony founding in ants (Hymenoptera; Formicidae). Oecologia 1989, 80, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Costa-Neto, E.M. The use of insects in folk medicine in the state of Bahia, northeastern Brazil, with notes on insects reported elsewhere in Brazilian folk medicine. Hum. Ecol. 2002, 30, 245–263. [Google Scholar] [CrossRef]
- Den Boer, S.P.A.; Baer, B.C.; Dreier, S.; Aron, S.; Nash, D.R.; Boomsma, J.J. Prudent sperm use by leaf-cutter ant queens. Proc. R. Soc. B 2009, 276, 3945–3953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marti, H.E.; Carlson, A.L.; Brown, B.V.; Mueller, U.G. Foundress queen mortality and early colony growth of the leafcutter ant, Atta texana (Formicidae, Hymenoptera). Insect. Soc. 2015, 62, 357–363. [Google Scholar] [CrossRef]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kooij, P.W.; Dentinger, B.M.; Donoso, D.A.; Shik, J.Z.; Gaya, E. Cryptic Diversity in Colombian Edible Leaf-Cutting Ants (Hymenoptera: Formicidae). Insects 2018, 9, 191. https://doi.org/10.3390/insects9040191
Kooij PW, Dentinger BM, Donoso DA, Shik JZ, Gaya E. Cryptic Diversity in Colombian Edible Leaf-Cutting Ants (Hymenoptera: Formicidae). Insects. 2018; 9(4):191. https://doi.org/10.3390/insects9040191
Chicago/Turabian StyleKooij, Pepijn W., Bryn M. Dentinger, David A. Donoso, Jonathan Z. Shik, and Ester Gaya. 2018. "Cryptic Diversity in Colombian Edible Leaf-Cutting Ants (Hymenoptera: Formicidae)" Insects 9, no. 4: 191. https://doi.org/10.3390/insects9040191