Risk Assessment of Two Insecticides on Encarsia formosa, Parasitoid of Whitefly Bemisia tabaci

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants, Insects and Insecticides
2.2. Toxicity to Parasitoid Adults
2.3. Toxicity to Parasitoid Pre-Pupae and Pupae
2.4. Sublethal Effects on Parasitoid Searching Behaviour
2.5. Data Analysis
3. Results
3.1. Toxicity to Parasitoid Adults
3.2. Toxicity to Parasitoid Pre-Pupae and Pupae
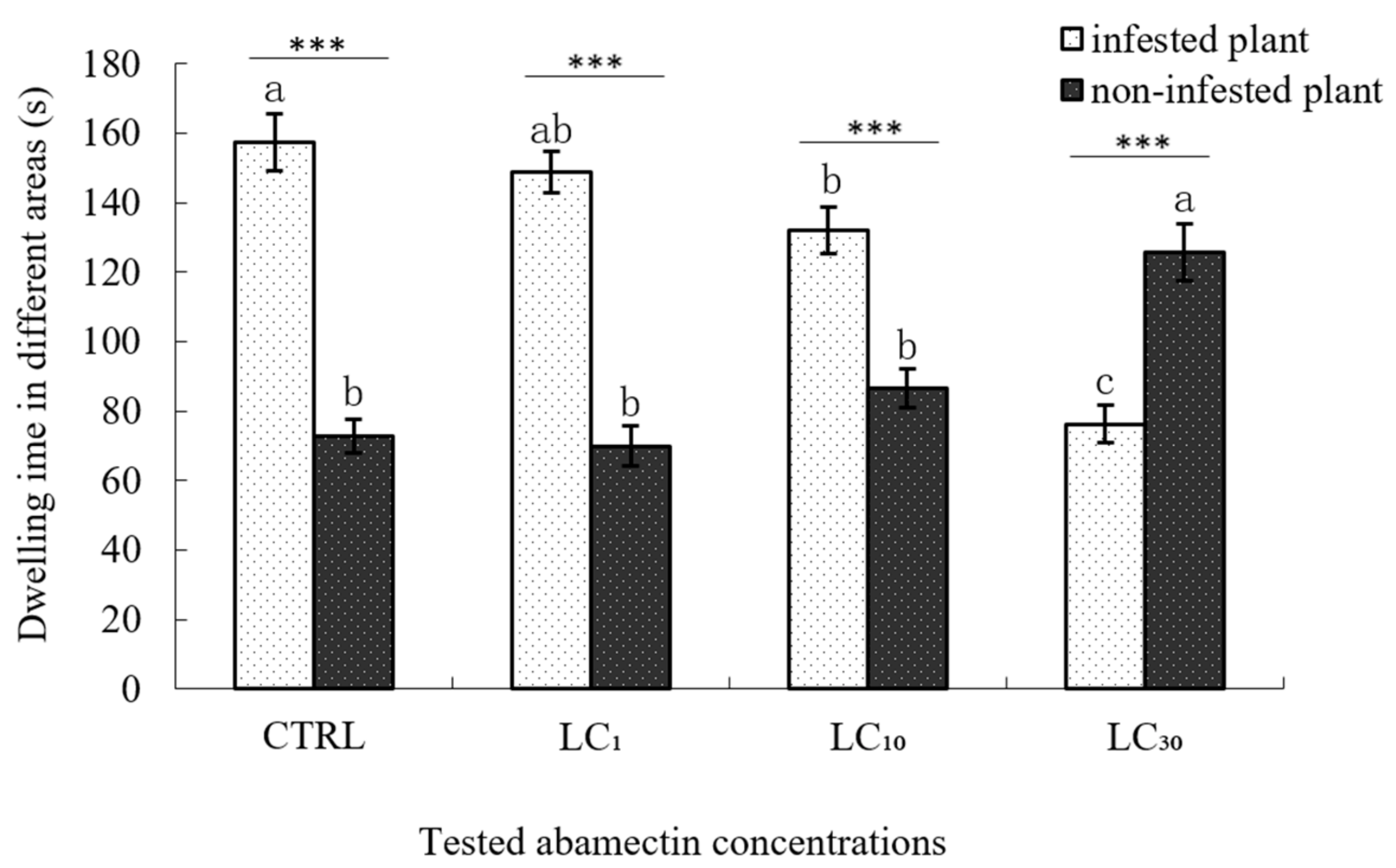
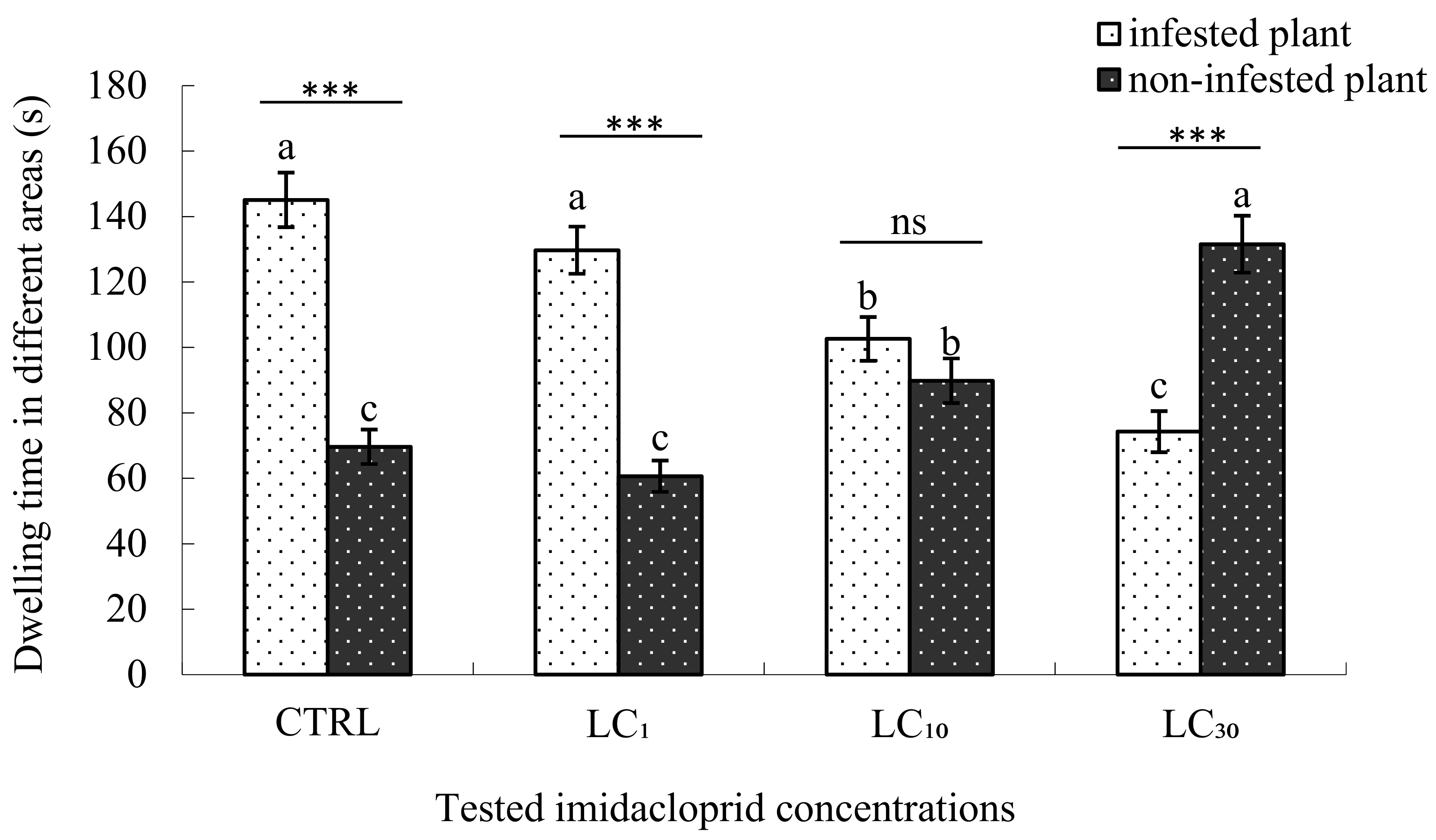
3.3. Sublethal Effects on Parasitoid Searching Behaviour
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brown, J.K.; Frohlich, D.R.; Rosell, R.C. The sweetpotato or silverleaf whiteflies: Biotypes of Bemisia tabaci oraspeciesco complex. Annu. Rev. Entomol. 1995, 40, 511–534. [Google Scholar] [CrossRef]
- De Barro, P.J.; Liu, S.S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A statement of species status. Annu. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Berlinger, M.J. Host plant resistance to Bemisia tabaci. Agric. Ecosyst. Environ. 1986, 17, 69–82. [Google Scholar] [CrossRef]
- Jones, D.R. Plant viruses transmitted by whiteflies. Eur. J. Plant Pathol. 2003, 109, 195–219. [Google Scholar] [CrossRef]
- Qiu, B.L.; Dang, F.; Li, S.J.; Ahmed, M.Z.; Jin, F.L.; Ren, S.X.; Cuthbertson, A.G.S. Comparison of biological parameters between the invasive B biotype and a new defined Cv biotype of Bemisia tabaci (Hemiptera: Aleyrodidae) in China. J. Pest Sci. 2011, 84, 419–427. [Google Scholar] [CrossRef]
- Su, Q.; Pan, H.P.; Liu, B.M.; Chu, D.; Xie, W.; Wu, Q.J.; Wang, S.L.; Xu, B.Y.; Zhang, Y.J. Insect symbiont facilitates vector acquisition, retention, and transmission of plant virus. Sci. Rep. 2013, 3, srep01367. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, A.G.S.; Buxton, J.H.; Blackburn, L.F.; Mathers, J.J.; Robinson, K.A.; Powell, M.E.; Fleming, D.A.; Bell, H.A. Eradicating Bemisia tabaci Q biotype on poinsettia plants in the UK. Crop Prot. 2012, 42, 42–48. [Google Scholar] [CrossRef]
- Basit, M.; Saeed, S.; Ahmad, M.; Sayyed, A.H. Can resistance in Bemisia tabaci (Homoptera: Aleyrodidae) be overcome with mixtures of neonicotinoids and insect growth regulators? Crop Prot. 2013, 44, 135–141. [Google Scholar] [CrossRef]
- Gerling, D.; Alomar, Ò.; Arnò, J. Biological control of Bemisia tabaci using predators and parasitoids. Crop Prot. 2001, 20, 779–799. [Google Scholar] [CrossRef]
- Bacci, L.; Crespo, A.L.B.; Galvan, T.L.; Pereira, E.J.G.; Picanco, M.C.; Silva, G.A.; Chediak, M. Toxicity of insecticides to the sweetpotato whitefly (Hemiptera: Aleyrodidae) and its natural enemies. Pest Manag. Sci. 2007, 63, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Li, S.J.; Xue, X.; Ahmed, M.Z.; Ren, S.X.; Du, Y.Z.; Wu, J.H.; Cuthbertson, A.G.S.; Qiu, B.L. Host plants and natural enemies of Bemisia tabaci (Homoptera: Aleyrodidae) in China. Insect Sci. 2011, 18, 101–120. [Google Scholar] [CrossRef]
- Van Lenteren, J.C.; Van Roermund, H.J.W.; Sutterlin, S. Biological control of greenhouse whitefly (Trialeurodes vaporariorum) with the parasitoid Encarsia formosa: How does it work? Biol. Control 1996, 6, 1–10. [Google Scholar] [CrossRef]
- Van Lenteren, J.C. A greenhouse without pesticides: Fact or fantasy. Crop Prot. 2000, 19, 375–384. [Google Scholar] [CrossRef]
- Sugiyama, K.; Katayama, H.; Saito, T. Effect of insecticides on the mortalities of three whitefly parasitoid species, Eretmocerus mundus, Eretmocerus eremicus and Encarsia formosa (Hymenoptera: Aphelinidae). Appl. Entomol. Zool. 2011, 46, 311–317. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Murchie, A.K. European red spider mite—An environmental consequence of persistent chemical pesticide application. Int. J. Environ. Sci. Technol. 2005, 2, 287–290. [Google Scholar] [CrossRef]
- Prabhaker, N.; Morse, J.G.; Castle, S.J.; Naranjo, S.E.; Henneberry, T.J.; Toscano, N.C. Toxicity of seven foliar insecticides to four insect parasitoids attacking citrus and cotton pests. J. Econ. Entomol. 2007, 100, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Bueno, A.F.; Batistela, M.J.; Bueno, R.C.O.F.; Franca-Neto, J.B.; Nishikawa, M.A.N.; Filho, A.L. Effects of integrated pest management, biological control and prophylactic use of insecticides on the management and sustainability of soybean. Crop Prot. 2011, 30, 937–945. [Google Scholar] [CrossRef]
- Biondi, A.; Desneux, N.; Siscaro, G.; Zappalà, L. Using organic-certified rather than synthetic pesticides may not be safer for biological control agents: Selectivity and side effects of 14 pesticides on the predator Orius laevigatus. Chemosphere 2012, 87, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, A.G.S.; Murchie, A.K. The environmental impact of an orchard winter wash and early season pesticide applications on both a beneficial and a pest mite species in Bramley apple orchards. Int. J. Environ. Sci. Technol. 2006, 3, 333–339. [Google Scholar] [CrossRef]
- Cloyd, R.A.; Bethke, J.A. Impact of neonicotinoid insecticides on natural enemies in greenhouse and interiorscape environments. Pest Manag. Sci. 2011, 67, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Walthall, W.K.; Stark, J.D. A comparison of acute mortality and population growth rate as endpoints of toxicological effect. Ecotoxicol. Environ. Saf. 1997, 37, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Mahdavi, V.; Saber, M.; Rafiee, D.H.; Mehrvar, A. Comparative study of the population level effects of carbaryl and abamectin on larval ectoparasitoid Habrobracon hebetor Say (Hymenoptera: Braconidae). BioControl 2011, 56, 823–830. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.S.; He, Y.R.; Guo, X.L.; Luo, Y.L. Acute toxicities and sublethal effects of some conventional insecticides on Trichogramma chilonis (Hymenoptera: Trichogrammatidae). J. Econ. Entomol. 2012, 105, 1157–1163. [Google Scholar] [CrossRef] [PubMed]
- Moura, R.; Garcia, P.; Cabral, S.; Soares, A.O. Does pirimicarb affect the voracity of the euriphagous predator, Coccinella undecimpunctata L. (Coleoptera: Coccinellidae)? Biol. Control 2006, 38, 363–368. [Google Scholar] [CrossRef]
- Palumbo, J.C.; Horowitzb, A.R.; Prabhaker, N. Insecticidal control and resistance management for Bemisia tabaci. Crop Prot. 2001, 20, 739–765. [Google Scholar] [CrossRef]
- Tomizawa, M.; Casida, J.E. Selective toxicity of neonicotinoids attributable to specificity of insect and mammalian nicotinic receptors. Annu. Rev. Entomol. 2003, 48, 339–364. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, P.; Nauen, R. Neonicotinoids—From zero to hero in insecticide chemistry. Pest Manag. Sci. 2008, 64, 1084–1098. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.P.; Wu, C.X.; Wang, Y.H.; Cang, T.; Chen, L.P.; Yu, R.X.; Wang, Q. Assessment of toxicity risk of insecticides used in rice ecosystem on Trichogramma japonicum, an egg parasitoid of rice lepidopterans. J. Econ. Entomol. 2012, 105, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Sohrabi, F.; Shishehbor, P.; Saber, M.; Mosaddegh, M.S. Lethal and sublethal effects of buprofezin and imidacloprid on the whitefly parasitoid Encarsia inaron (Hymenoptera: Aphelinidae). Crop Prot. 2012, 32, 83–89. [Google Scholar] [CrossRef]
- Sohrabi, F.; Shishehbor, P.; Saber, M.; Mosaddegh, M.S. Lethal and sublethal effects of imidacloprid and buprofezin on the sweetpotato whitefly parasitoid Eretmocerus mundus (Hymenoptera: Aphelinidae). Crop Prot. 2013, 45, 98–103. [Google Scholar] [CrossRef]
- Biondi, A.; Zappala, L.; Stark, J.D.; Desneux, N. Do biopesticides affect the demographic traits of a parasitoid wasp and its biocontrol services through sublethal effects? PLoS ONE 2013, 8, e76548. [Google Scholar] [CrossRef] [PubMed]
- Ohta, I.; Takeda, M. Acute toxicities of 42 pesticides used for green peppers to an aphid parasitoid, Aphidius gifuensis (Hymenoptera: Braconidae), in adult and mummy stages. Appl. Entomol. Zool. 2015, 50, 207–212. [Google Scholar] [CrossRef]
- Chitgar, M.G.; Ghadamyari, M. Effects of Amitraz on the parasitoid Encarsia formosa (Gahan) (Hymenoptera: Aphelinidae) for Control of Trialeurodes vaporariorum Westwood (Homoptera: Aleyrodidae): IOBC Methods. J. Entomol. Res. Soc. 2012, 14, 61–69. [Google Scholar]
- Desneux, N.; Pham-Delègue, M.H.; Kaiser, L. Effects of sub-lethal and lethal doses of lambda-cyhalothrin on oviposition experience and host-searching behaviour of a parasitic wasp, Aphidius ervi. Pest Manag. Sci. 2004, 60, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, A.G.S.; Blackburn, L.F.; Northing, P.; Luo, W.; Cannon, R.J.C.; Walters, K.F.A. Leaf dipping as an environmental screening measure to test chemical efficacy against Bemisia tabaci on poinsettia plants. Int. J. Environ. Sci. Technol. 2009, 6, 347–352. [Google Scholar] [CrossRef]
- General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China and Standardization Administration of the People’s Republic of China. Test Guidelines on Environmental Safety Assessment for Chemical Pesticides―Part 17: Trichogramma Acute Toxicity Test; China Zhijian Publishing House & Standards Press of China: Beijing, China, 2014; pp. 1–12. [Google Scholar]
- De Morais, M.R.; Zanardi, O.Z.; Rugno, G.R.; Yamamoto, P.T. Impact of five insecticides used to control citrus pests on the parasitoid Ageniaspis citricola Longvinovskaya (Hymenoptera: Encyrtidae). Ecotoxicology 2016, 25, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Vanaclocha, P.; Vidal-Quist, C.; Oheix, S.; Monton, H.; Planes, L.; Catalan, J.; Tena, A.; Verdu, M.J.; Urbaneja, A. Acute toxicity in laboratory tests of fresh and aged residues of pesticides used in citrus on the parasitoid Aphytis melinus. J. Pest Sci. 2013, 86, 329–336. [Google Scholar] [CrossRef]
- Prabhaker, N.; Castle, S.J.; Naranjo, S.E.; Toscano, N.C.; Morse, J.G. Compatibility of two systemic neonicotinoids, imidacloprid and thiamethoxam, with various natural enemies of agricultural pests. J. Econ. Entomol. 2011, 104, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Saber, M. Acute and population level toxicity of imidacloprid and fenpyroximate on an important egg parasitoid, Trichogramma cacoeciae (Hymenoptera: Trichogrammatidae). Ecotoxicology 2011, 20, 1476–1484. [Google Scholar] [CrossRef] [PubMed]
- Suh, C.P.C.; Orr, D.B.; van Duyn, J.W. Effect of insecticides on Trichogramma exiguum (Trichogrammatidae: Hymenoptera) preimaginal development and adult survival. J. Econ. Entomol. 2000, 93, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Preetha, G.; Stanley, J.; Suresh, S.; Kuttalam, S.; Samiyappan, R. Toxicity of selected insecticides to Trichogramma chilonis: Assessing their safety in the rice ecosystem. Phytoparasitica 2009, 37, 209–215. [Google Scholar] [CrossRef]
- Wang, H.Y.; Yang, Y.; Su, J.Y.; Shen, J.L.; Gao, C.F.; Zhu, Y.C. Assessment of the impact of insecticides on Anagrus nilaparvatae (Pang et Wang) (Hymenoptera: Mymanidae), an egg parasitoid of the rice planthopper, Nilaparvata lugens (Hemiptera: Delphacidae). Crop Prot. 2008, 27, 514–522. [Google Scholar] [CrossRef]
- Hidrayani; Purnomo; Rauf, A.; Ridland, P.M.; Hoffmann, A.A. Pesticide applications on Java potato fields are ineffective in controlling leafminers, and have antagonistic effects on natural enemies of leafminers. Int. J. Pest Manag. 2005, 51, 181–187. [Google Scholar] [CrossRef]
- Mohammed, A.A.H.; Desneux, N.; Fan, Y.J.; Han, P.; Ali, A.; Song, D.L.; Gao, X.W. Impact of imidacloprid and natural enemies on cereal aphids: Integration or ecosystem service disruption? Entomol. Gen. 2017, 1, 47–61. [Google Scholar] [CrossRef]
- Varenhorst, A.J.; O’Neal, M.E. The response of natural enemies to selective insecticides applied to soybean. Environ. Entomol. 2012, 41, 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Tran, D.H.; Takagi, M.; Takasu, K. Effects of selective insecticides on host searching and oviposition behavior of Neochrysocharis formosa (Westwood) (Hymenoptera: Eulophidae), a larval parasitoid of the American serpentine leafminer. Appl. Entomol. Zool. 2004, 39, 435–441. [Google Scholar] [CrossRef]
- Liu, F.; Bao, S.W.; Song, Y.; Lu, H.Y.; Xu, J.X. Effects of imidacloprid on the orientation behavior and parasitizing capacity of Anagrus nilaparvatae, an egg parasitoid of Nilaparvata lugens. BioControl 2010, 55, 473–483. [Google Scholar] [CrossRef]
- Bethke, J.A.; Redak, R.A. Effect of imidacloprid on the silverleaf whitefly, Bemisia argentifolii Bellows and Perring (Homoptera: Aleyrodidae), and whitefly parasitism. Ann. Appl. Biol. 1997, 130, 397–407. [Google Scholar] [CrossRef]
- Desneux, N.; Rafalimanana, H.; Kaiser, L. Dose-response relationship in lethal and behavioural effects of different insecticides on the parasitic wasp Aphidius ervi. Chemosphere 2004, 54, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Bayram, A.; Salerno, G.; Onofri, A.; Conti, E. Sublethal effects of two pyrethroids on biological parameters and behavioral responses to host cues in the egg parasitoid Telenomus busseolae. Biol. Control 2010, 53, 153–160. [Google Scholar] [CrossRef]
- Abbes, K.; Biondi, A.; Kurtulus, A.; Ricupero, M.; Russo, A.; Siscaro, G.; Chermiti, B.; Zappala, L. Combined non-target effects of insecticide and high temperature on the parasitoid Bracon nigricans. PLoS ONE 2015, 10, e0138411. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxicity Risk Grade | Safety Factor (SF) |
|---|---|
| extreme high risk | SF ≤ 0.05 |
| high risk | 0.05 < SF ≤ 0.5 |
| medium risk | 0.5 < SF ≤ 5 |
| low risk | SF > 5 |
| Parasitoid Age (h) | Regression Equation of Toxicity | Correlation Coefficient (r) | LC30 (mg/L) (95% Confidence Interval) | LC50 (mg/L) (95% Confidence Interval) |
|---|---|---|---|---|
| 12 | Y = −0.396 + 1.472X | 0.966 | 0.818 (0.255–1.456) | 1.857 (0.891–2.771) |
| 24 | Y = −1.829 + 2.541X | 0.928 | 3.262 (0.443–5.839) | 5.246 (1.894–11.081) |
| 48 | Y = −1.770 + 2.162X | 0.973 | 3.768 (2.805–4.707) | 6.587 (5.330–8.078) |
| 72 | Y = −2.092 + 2.499X | 0.987 | 4.239 (3.312–5.150) | 6.872 (5.695–8.266) |
| 96 | Y = −2.439 + 2.560X | 0.991 | 5.598 (4.457–6.740) | 8.972 (7.490–10.804) |
| Parasitoid Age (h) | Regression Equation of Toxicity | Correlation Coefficient (r) | LC30 (mg/L) (95% Confidence Interval) | LC50 (mg/L) (95% Confidence Interval) |
|---|---|---|---|---|
| 12 | Y = −2.854 + 1.740X | 0.976 | 21.818 (12.965–30.399) | 43.669 (31.555–56.3) |
| 24 | Y = −3.789 + 2.034X | 0.974 | 40.262 (29.717–50.668) | 72.892 (58.593–90.425) |
| 48 | Y = −4.441 + 2.301X | 0.981 | 43.818 (30.749–56.726) | 81.296 (66.387–99.827) |
| 72 | Y = −3.392 + 1.747X | 0.983 | 46.943 (35.828–57.980) | 87.470 (68.605–112.343) |
| 96 | Y = −3.399 + 1.688X | 0.986 | 50.445 (35.926–65.201) | 103.138 (80.635–135.647) |
| Pesticide | Parasitoid Age (h) | Field Recommended Concentration (mg/L) | Safety Factor (SF) | Toxicity Risk Grade |
|---|---|---|---|---|
| abamectin | 12 | 20 | 0.093 | high risk |
| 24 | 20 | 0.262 | high risk | |
| 48 | 20 | 0.329 | high risk | |
| 72 | 20 | 0.344 | high risk | |
| 96 | 20 | 0.449 | high risk | |
| imidacloprid | 12 | 33.3 | 1.311 | medium risk |
| 24 | 33.3 | 2.189 | medium risk | |
| 48 | 33.3 | 2.441 | medium risk |
| Parasitoid Stage | Regression Equation of Toxicity | Correlation Coefficient (r) | LC30 (mg/L) (95% Confidence Interval) | LC50 (mg/L) (95% Confidence Interval) |
|---|---|---|---|---|
| pre-pupa | Y = −2.086 + 3.244X | 0.951 | 3.029 (0.386–5.167) | 4.395 (1.447–8.493) |
| pupa | Y = −1.372 + 2.109X | 0.990 | 2.524 (1.730–3.278) | 4.474 (3.473–5.532) |
| Pesticide | Parasitoid Stage | Field Recommended Concentration (mg/L) | Safety Factor (SF) | Toxicity Risk Grade |
|---|---|---|---|---|
| abamectin | pre-pupa | 20 | 0.220 | high risk |
| pupa | 20 | 0.224 | high risk | |
| imidacloprid | pre-pupa | 33.3 | 0.667 | medium risk |
| pupa | 33.3 | 0.993 | medium risk |
| Parasitoid Stage | Regression Equation of Toxicity | Correlation Coefficient (r) | LC30 (mg/L) (95% Confidence Interval) | LC50 (mg/L) (95% Confidence Interval) |
|---|---|---|---|---|
| pre-pupa | Y = −1.773 + 1.317X | 0.931 | 8.877 (2.822–15.911) | 22.211 (11.107–33.026) |
| pupa | Y = −3.024 + 2.032X | 0.951 | 16.990 (1.210–32.911) | 30.781 (6.680–54.553) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Z.; Liu, Y.; Wang, L.; Guo, Q.; Ali, S.; Chen, X.-S.; Qiu, B.-L. Risk Assessment of Two Insecticides on Encarsia formosa, Parasitoid of Whitefly Bemisia tabaci. Insects 2018, 9, 116. https://doi.org/10.3390/insects9030116
He Z, Liu Y, Wang L, Guo Q, Ali S, Chen X-S, Qiu B-L. Risk Assessment of Two Insecticides on Encarsia formosa, Parasitoid of Whitefly Bemisia tabaci. Insects. 2018; 9(3):116. https://doi.org/10.3390/insects9030116
Chicago/Turabian StyleHe, Zhan, Yuan Liu, Lei Wang, Qiu Guo, Shaukat Ali, Xiao-Sheng Chen, and Bao-Li Qiu. 2018. "Risk Assessment of Two Insecticides on Encarsia formosa, Parasitoid of Whitefly Bemisia tabaci" Insects 9, no. 3: 116. https://doi.org/10.3390/insects9030116