
Sublethal Effects of Spirotetramat, Cyantraniliprole, and Pymetrozine on Aphis gossypii (Hemiptera: Aphididae)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect
2.2. Chemicals
2.3. Insecticidal Toxicity Experiment
2.4. Development and Reproduction of A. gossypii at Sublethal Concentrations
2.5. Measurement of the Growth Parameters of A. gossypii after Treatment with Sublethal Concentrations of the Three Pesticides
2.6. Data Analysis
3. Results
3.1. Toxicity to A. gossypii at the Recommended Concentrations of the Three Insecticides
3.2. Sublethal Response of A. gossypii to the Three Insecticides
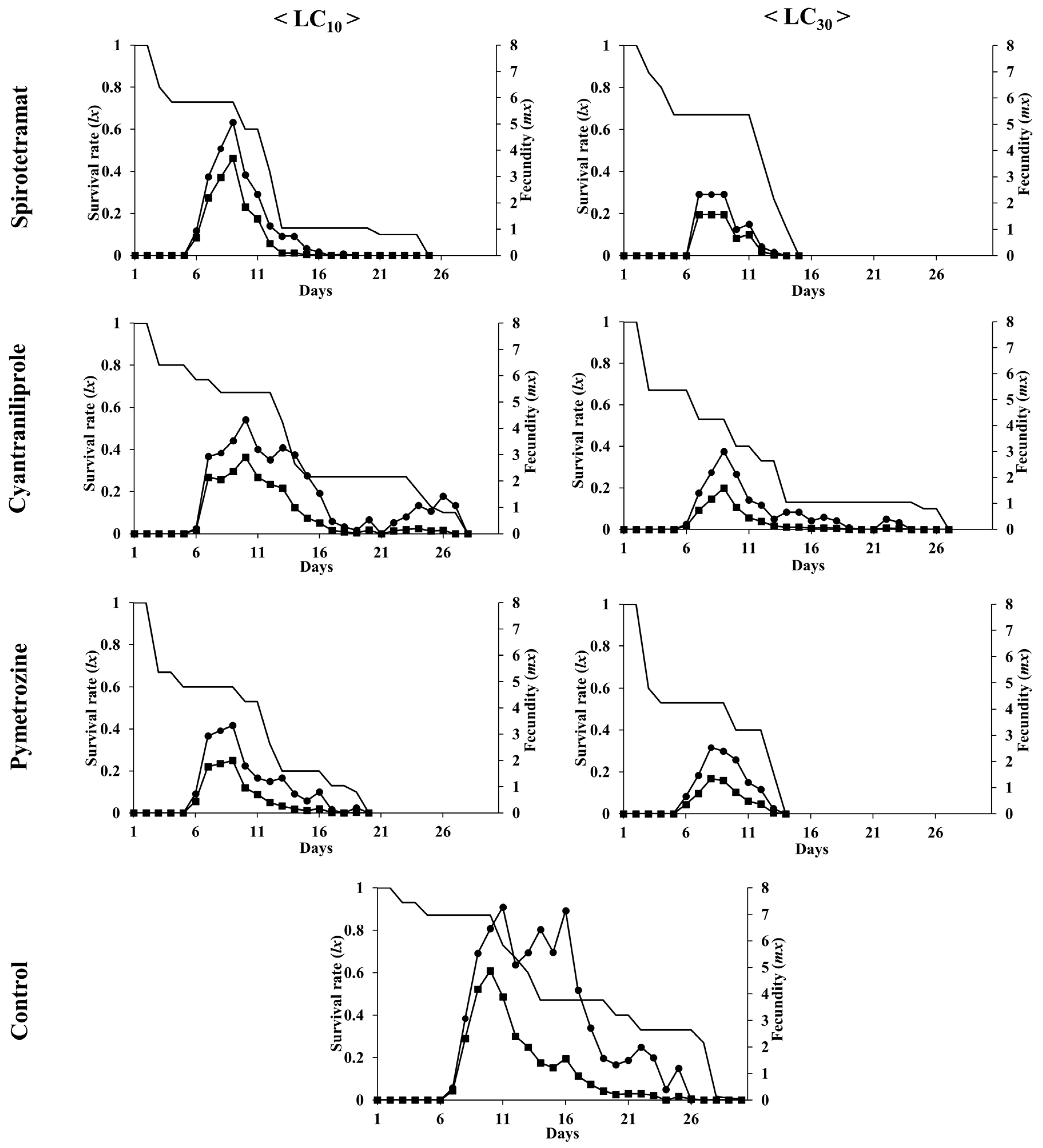
3.3. Effects of Insecticides on the Population Growth of A. gossypii
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ebert, T.; Cartwright, B. Biology and ecology of Aphis gossypii Glover (Homoptera: Aphididae). Southwest. Entomol. 1997, 22, 116–153. [Google Scholar]
- Saito, T.; Hama, H.; Suzuki, K. Insecticide resistance in clones of the cotton aphid, Aphis gossypii Glover (Homoptera: Aphididae), and synergistic effect of esterase and mixed-function oxidase inhibitors. Jpn. J. Appl. Entomol. Zool. 1995, 39, 151–158. [Google Scholar] [CrossRef]
- Herron, G.A.; Wilson, L.J. Can resistance management strategies recover insecticide susceptibility in pests?: A case study with cotton aphid Aphis gossypii (Aphididae: Hemiptera) in Australian cotton. Austral Entomol. 2017, 56, 1–13. [Google Scholar] [CrossRef]
- Koo, H.N.; An, J.J.; Park, S.E.; Kim, J.I.; Kim, G.H. Regional susceptibilities to 12 insecticides of melon and cotton aphid, Aphis gossypii (Hemiptera: Aphididae) and a point mutation associated with imidacloprid resistance. Crop Prot. 2014, 55, 91–97. [Google Scholar] [CrossRef]
- Carletto, J.; Martin, T.; Vanlerberghe-Masutti, F.; Brévault, T. Insecticide resistance traits differ among and within host races in Aphis gossypii. Pest Manag. Sci. 2010, 66, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tie, M.; Chen, A.; Ma, K.; Li, F.; Liang, P.; Liu, Y.; Song, D.; Gao, X. Pyrethroid resistance associated with M918 L mutation and detoxifying metabolism in Aphis gossypii from Bt cotton growing regions of China. Pest Manag. Sci. 2017, 73, 2353–2359. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Biondi, A.; Desneux, N.; Gao, X.W. Assessment of physiological sublethal effects of imidacloprid on the mirid bug Apolygus lucorum (Meyer-Dür). Ecotoxicology 2012, 21, 1989–1997. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhao, J.; Zheng, Y.; Weng, Q.; Biondi, A.; Desneux, N.; Wu, K. Assessment of potential sublethal effects of various insecticides on key biological traits of the tobacco whitefly, Bemisia tabaci. Int. J. Biol. Sci. 2013, 9, 246. [Google Scholar] [CrossRef]
- Kerns, D.; Stewart, S. Sublethal effects of insecticides on the intrinsic rate of increase of cotton aphid. Entomol. Exp. Appl. 2000, 94, 41–49. [Google Scholar] [CrossRef]
- Stark, J.D.; Banks, J.E. Population-level effects of pesticides and other toxicants on arthropods. Annu. Rev. Entomol. 2003, 48, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.-Y.; Yang, X.-M.; Sun, L.-J.; Zhao, C.-D.; Chi, H.; Zheng, C.-Y. Sublethal effect of spirotetramat on the life table and population growth of Frankliniella occidentalis (Thysanoptera: Thripidae). Entomol. Gen. 2021, 41, 219–231. [Google Scholar] [CrossRef]
- Iftikhar, A.; Hafeez, F.; Hashim, M.; Rehman, M. Assessment of sublethal and transgenerational effects of spirotetramat, on population growth of cabbage aphid, Brevicoryne brassicae L. (Hemiptera: Aphididae). Front. Physiol. 2022, 13, 1014190. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.M.; Feng, H.L.; Wang, G.D.; Zhang, L.L.; Zulu, L.; Liu, Y.H.; Zheng, Y.L.; Rao, Q. Sublethal effects of three insecticides on development and reproduction of Spodoptera frugiperda (Lepidoptera: Noctuidae). Agronomy 2022, 12, 1334. [Google Scholar] [CrossRef]
- Lutz, A.L.; Bertolaccini, I.; Scotta, R.R.; Curis, M.C.; Favaro, M.A.; Fernandez, L.N.; Sánchez, D.E. Lethal and sublethal effects of chlorantraniliprole on Spodoptera cosmioides (Lepidoptera: Noctuidae). Pest Manag. Sci. 2018, 74, 2817–2821. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.M.; Dong, J.F.; Chen, J.H.; Ji, Q.E.; Cui, J.J. The sublethal effects of chlorantraniliprole on Helicoverpa armigera (Lepidoptera: Noctuidae). J. Integr. Agric. 2013, 12, 457–466. [Google Scholar] [CrossRef]
- Liang, P.Z.; Ma, K.S.; Chen, X.W.; Tang, C.Y.; Xia, J.; Chi, H.; Gao, X.W. Toxicity and sublethal effects of flupyradifurone, a novel butenolide insecticide, on the development and fecundity of Aphis gossypii (Hemiptera: Aphididae). J. Econ. Entomol. 2019, 112, 852–858. [Google Scholar] [CrossRef] [PubMed]
- Xin, J.J.; Yu, W.X.; Yi, X.Q.; Gao, J.P.; Gao, X.W.; Zeng, X.P. Sublethal effects of sulfoxaflor on the fitness of two species of wheat aphids, Sitobion avenae (F.) and Rhopalosiphum padi (L.). J. Integr. Agric. 2019, 18, 1613–1623. [Google Scholar] [CrossRef]
- Lümmen, P.; Khajehali, J.; Luther, K.; Van Leeuwen, T. The cyclic keto-enol insecticide spirotetramat inhibits insect and spider mite acetyl-CoA carboxylases by interfering with the carboxyltransferase partial reaction. Insect Biochem. Mol. Biol. 2014, 55, 1–8. [Google Scholar] [CrossRef]
- Brück, E.; Elbert, A.; Fischer, R.; Krueger, S.; Kühnhold, J.; Klueken, A.M.; Nauen, R.; Niebes, J.F.; Reckmann, U.; Schnorbach, H.J. Movento®, an innovative ambimobile insecticide for sucking insect pest control in agriculture: Biological profile and field performance. Crop Prot. 2009, 28, 838–844. [Google Scholar] [CrossRef]
- Nauen, R.; Reckmann, U.; Thomzik, J.; Thielert, W. Biological profile of spirotetramat (Movento®)—A new two-way systemic (ambimobile) insecticide against sucking pest species. Bayer CropSci. J. 2008, 61, 245–278. [Google Scholar]
- Salazar-López, N.J.; Aldana-Madrid, M.L.; Silveira-Gramont, M.I.; Aguiar, J.L. Spirotetramat—An Alternative for the Control of Parasitic Sucking Insects and Its Fate in the Environment; ImntechOpen: London, UK, 2016; ISBN 978-953-51-2258-6. [Google Scholar]
- Kühnhold, J.; Klueken, A.; De Maeyer, L.; Van Waetermeulen, X.; Brück, E.; Elbert, A. Movento®, an innovative solution for sucking insect pest control in agriculture: Field performance in fruits and vegetables. Bayer CropSci. J. 2008, 61, 279–306. [Google Scholar]
- Vermeer, R.; Baur, P. Movento® product development: Custom-made formulations for an exceptional active ingredient. Bayer CropSci. J. 2008, 61, 141–157. [Google Scholar]
- Teixeira, L.A.; Andaloro, J.T. Diamide insecticides: Global efforts to address insect resistance stewardship challenges. Pestic. Biochem. Physiol. 2013, 106, 76–78. [Google Scholar] [CrossRef]
- Foster, S.P.; Denholm, I.; Rison, J.L.; Portillo, H.E.; Margaritopoulis, J.; Slater, R. Susceptibility of standard clones and European field populations of the green peach aphid, Myzus persicae, and the cotton aphid, Aphis gossypii (Hemiptera: Aphididae), to the novel anthranilic diamide insecticide cyantraniliprole. Pest Manag. Sci. 2012, 68, 629–633. [Google Scholar] [CrossRef]
- Cordova, D.; Benner, E.; Sacher, M.; Rauh, J.; Sopa, J.; Lahm, G.; Selby, T.; Stevenson, T.; Flexner, L.; Gutteridge, S. Anthranilic diamides: A new class of insecticides with a novel mode of action, ryanodine receptor activation. Pestic. Biochem. Physiol. 2006, 84, 196–214. [Google Scholar] [CrossRef]
- Sattelle, D.B.; Cordova, D.; Cheek, T.R. Insect ryanodine receptors: Molecular targets for novel pest control chemicals. Invertebr. Neurosci. 2008, 8, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Thrash, B.; Adamczyk, J.; Lorenz, G.; Scott, A.; Armstrong, J.; Pfannenstiel, R.; Taillon, N. Laboratory evaluations of lepidopteran-active soybean seed treatments on survivorship of fall armyworm (Lepidoptera: Noctuidae) larvae. Fla. Entomol. 2013, 96, 724–728. [Google Scholar] [CrossRef]
- Bielza, P.; Guillén, J. Cyantraniliprole: A valuable tool for Frankliniella occidentalis (Pergande) management. Pest Manag. Sci. 2015, 71, 1068–1074. [Google Scholar] [CrossRef]
- Zhang, R.; He, S.; Chen, J. Monitoring of Bactrocera dorsalis (Diptera: Tephritidae) resistance to cyantraniliprole in the south of China. J. Econ. Entomol. 2014, 107, 1233–1238. [Google Scholar] [CrossRef]
- Barry, J.D.; Portillo, H.E.; Annan, I.B.; Cameron, R.A.; Clagg, D.G.; Dietrich, R.F.; Watson, L.J.; Leighty, R.M.; Ryan, D.L.; McMillan, J.A. Movement of cyantraniliprole in plants after foliar applications and its impact on the control of sucking and chewing insects. Pest Manag. Sci. 2015, 71, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Fuog, D.; Fergusson, S.J.; Flückiger, C. Pymetrozine: A novel insecticide affecting aphids and whiteflies. In Insecticides with Novel Modes of Action: Mechanisms and Application, 2nd ed.; Ishaaya, I., Degheele, D., Eds.; Springer: Berlin/Heidelberg, Germany, 1998; pp. 40–49. [Google Scholar]
- Stark, J.D.; Wennergren, U. Can population effects of pesticides be predicted from demographic toxicological studies? J. Econ. Entomol. 1995, 88, 1089–1096. [Google Scholar] [CrossRef]
- Harrewijn, P.; Kayser, H. Pymetrozine, a fast-acting and selective inhibitor of aphid feeding. In-situ studies with electronic monitoring of feeding behaviour. Pestic. Sci. 1997, 49, 130–140. [Google Scholar] [CrossRef]
- SAS Institute. SAS User’s Guide, Statistics Version 9, 1st ed.; SAS Institute: Cary, NC, USA, 2009. [Google Scholar]
- Dunnam, E.; Clark, J. Cotton aphid multiplication following treatment with calcium arsenate. J. Econ. Entomol. 1941, 34, 587–588. [Google Scholar] [CrossRef]
- Kidd, P.; Rummel, D. Effect of insect predators and a pyrethroid insecticide on cotton aphid, Aphis gossypii Glover, population density. Southwest. Entomol. 1997, 22, 381–393. [Google Scholar]
- Slosser, J.; Pinchak, W.; Rummel, D. A review of known and potential factors affecting the population dynamics of the cotton aphid. Southwest. Entomol. 1989, 14, 302–313. [Google Scholar]
- Kerns, D.; Gaylor, M. Induction of cotton aphid outbreaks by insecticides in cotton. Crop Prot. 1993, 12, 387–393. [Google Scholar] [CrossRef]
- Robertson, J.L.; Jones, M.M.; Olguin, E.; Alberts, B. Bioassays with Arthropods, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Wang, Z.H.; Gong, Y.J.; Jin, G.H.; Zhu, L.; Wei, S.J. Effects of spirotetramat on development and reproduction of (Hemiptera: Aphididae). Austral Entomol. 2016, 55, 235–241. [Google Scholar] [CrossRef]
- Abdel-Fatah, R.M.; Mohamed, S.M.; Aly, A.A.; Sabry, A.-K.H. Biochemical characterization of spiromesifen and spirotetramat as lipid synthesis inhibitors on cotton leaf worm, Spodoptera littoralis. Bull. Natl. Res. Cent. 2019, 43, 65. [Google Scholar] [CrossRef]
- Yang, X.; Zhou, G.; Sun, L.; Zheng, C. Ovicidal activity of spirotetramat and its effect on hatching, development and formation of Frankliniella occidentalis egg. Sci. Rep. 2021, 11, 20751. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Marshall, K.E. The many roles of fats in overwintering insects. J. Exp. Biol. 2018, 7, 221. [Google Scholar] [CrossRef] [PubMed]
- Emile, V. Fuel metabolism of the mosquito (Culex quinquefasciatus) embryo. J. Insect Physiol. 1993, 39, 831–833. [Google Scholar]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef] [PubMed]
- Beenakkers, A.T.; Bloemen, R.; De Vlieger, T.; Van der Horst, D.; Van Marrewijk, W. Insect adipokinetic hormones. Peptides 1985, 6, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aal, A.E. Effect of chlorfluazuron, nuclear polyhydrosis virus (SLNPV) and Bacillus thuringiensis on some biological and enzymes activity of cotton leafworm, Spodoptera littoralis (Boisd). Bull. Entomol. Soc. Egypt/Econ. Ser. 2006, 32, 171–185. [Google Scholar]
- Sak, O.; Uçkan, F.; Ergin, E. Effects of cypermethrin on total body weight, glycogen, protein, and lipid contents of Pimpla turionellae (L.) (Hymenoptera: Ichneumonidae). Belg. J. Zool. 2006, 136, 53–58. [Google Scholar]
- Alimohammadi, N.; Samih, M.; Izadi, H.; Shahidi Noghabi, S. Developmental and biochemical effects of hexaflumuron and spirodiclofen on the ladybird beetle, Hippodamia variegata (Goeze) (Coleoptera: Coccinellidae). J. Crop Prot. 2014, 3, 335–344. [Google Scholar]
- Ali, N.S.; Ali, S.S.; Shakoori, A.R. Biochemical response of malathion-resistant and-susceptible adults of Rhyzopertha dominica to the sublethal doses of deltamethrin. Pak. J. Zool. 2014, 46, 853–861. [Google Scholar]
- Amrutsagar, M.; Joshi, S. Insect repellent induced lipid diversions of Periplaneta americana. Int. J. Recent Sci. Res. 2017, 8, 18370–18372. [Google Scholar]
- Lahm, G.P.; Cordova, D.; Barry, J.D. New and selective ryanodine receptor activators for insect control. Bioorgan. Med. Chem. 2009, 17, 4127–4133. [Google Scholar] [CrossRef]
- Hummel, N.; Mészáros, A.; Ring, D.; Beuzelin, J.; Stout, M. Evaluation of seed treatment insecticides for management of the rice water weevil, Lissorhoptrus oryzophilus Kuschel (Coleoptera: Curculionidae), in commercial rice fields in Louisiana. Crop Prot. 2014, 65, 37–42. [Google Scholar] [CrossRef]
- Lanka, S.K.; Blouin, D.C.; Stout, M.J. Integrating flood depth and plant resistance with chlorantraniliprole seed treatments for management of rice water weevil, Lissorhoptrus oryzophilus (Coleoptera: Curculionidae). Insect Sci. 2015, 22, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xu, C.; Ding, J.; Zhao, Y.; Lin, J.; Liu, F.; Mu, W. Cyantraniliprole seed treatment efficiency against Agrotis ipsilon (Lepidoptera: Noctuidae) and residue concentrations in corn plants and soil. Pest Manag. Sci. 2019, 75, 1464–1472. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Wang, K.; Li, Y.; Wang, S. Lethal and sublethal effects of cyantraniliprole on Helicoverpa assulta (Lepidoptera: Noctuidae). Pestic. Biochem. Physiol. 2017, 136, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Zhang, S.; Shen, F.; Liu, M.; Ren, C.; Gao, X. Residual toxicity and sublethal effects of chlorantraniliprole on Plutella xylostella (Lepidoptera: Plutellidae). Pest Manag. Sci. 2012, 68, 1184–1190. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Yao, Y.S.; Zhu, X.Z.; Wang, L.; Li, D.Y.; Zhang, K.X.; Gao, X.K.; Wu, C.C.; Niu, L.; Ji, J.C. Evaluation of sublethal and transgenerational effects of sulfoxaflor on Aphis gossypii via life table parameters and 16S rRNA sequencing. Pest Manag. Sci. 2021, 77, 3406–3418. [Google Scholar] [CrossRef] [PubMed]
- Lashkari, M.R.; Sahragard, A.; Ghadamyari, M. Sublethal effects of imidacloprid and pymetrozine on population growth parameters of cabbage aphid, Brevicoryne brassicae on rapeseed, Brassica napus L. Insect Sci. 2007, 14, 207–212. [Google Scholar] [CrossRef]
- Amini Jam, N.; Kocheili, F.; Mossadegh, M.S.; Rasekh, A.; Saber, M. Lethal and sublethal effects of imidacloprid and pirimicarb on the melon aphid, Aphis gossypii Glover (Hemiptera: Aphididae) under laboratory conditions. J. Crop Prot. 2014, 3, 89–98. [Google Scholar]
- Rasheed, M.A.; Khan, M.M.; Hafeez, M.; Zhao, J.; Islam, Y.; Ali, S.; Ur-Rehman, S.; e-Hani, U.; Zhou, X. Lethal and sublethal effects of chlorpyrifos on biological traits and feeding of the aphidophagous predator Harmonia axyridis. Insects 2020, 11, 491. [Google Scholar] [CrossRef]


{kind=link}
| Insecticides | Stage | n a | DAT b | LC10 | RT d | LC30 | RT | LC50 | RT | LC70 | RT | LC90 | RT | Slope ± SE | df | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (95% CL c) | (95% CL) | (95% CL) | (95% CL) | (95% CL) | ||||||||||||
| Spirotetramat | Nymph | 677 | 4 | 0.07 (0.05–0.10) | 8.0 | 0.23 (0.18–0.29) | 8.2 | 0.52 (0.43–0.61) | 8.9 | 1.14 (0.96–1.37) | 8.9 | 3.58 (2.80–4.84) | 9.5 | 1.53 ± 0.10 | 6 | <0.0001 |
| Adult | 500 | 4 | 0.56 (0.38–0.77) | 1.89 (1.47–2.33) | 4.36 (3.62–5.21) | 10.09 (8.37–12.45) | 33.89 (25.76–47.68) | 1.44 ± 0.10 | 5 | <0.0001 | ||||||
| Cyantraniliprole | Nymph | 599 | 3 | 0.04 (0.03–0.05) | 2.5 | 0.15 (0.12–0.18) | 3.3 | 0.38 (0.32–0.46) | 3.9 | 1.00 (0.81–1.25) | 4.5 | 3.99 (2.95–5.75) | 5.4 | 1.26 ± 0.07 | 7 | <0.0001 |
| Adult | 478 | 3 | 0.10 (0.06–0.15) | 0.50 (0.37–0.65) | 1.50 (1.20–1.88) | 4.47 (3.50–5.92) | 21.61 (1.94–34.38) | 1.11 ± 0.08 | 5 | <0.0001 | ||||||
| Pymetrozine | Nymph | 571 | 3 | 0.34 (0.21–0.51) | 1.2 | 1.76 (1.30–2.26) | 1.4 | 5.46 (4.42–6.68) | 1.7 | 16.95 (13.68–21.56) | 1.9 | 86.95 (62.22–131.65) | 2.3 | 1.07 ± 0.07 | 7 | <0.0001 |
| Adult | 501 | 3 | 0.41 (0.22–0.66) | 2.55 (1.79–3.48) | 9.08 (6.93–11.78) | 32.34 (24.48–44.45) | 202.27 (132.31–345.99) | 0.95 ± 0.07 | 5 | <0.0001 |
| Insecticides | Treatment | P | F1 | F2 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Developmental Period (Day) | Adult Survival Rate (%) | Adult Longevity (Day) | Fecundity (%) | Deformity Rate (%) | Developmental Period (Day) | Adult Survival Rate (%) | Fecundity (%) | Deformity Rate (%) | ||
| Spirotetramat | Control | 5.35 ± 0.16 b | 96.30 ± 0.55 a | 16.01 ± 0.91 a | 100.0 ± 0.00 a | 0.0 ± 0.00 c | 5.06 ± 0.17 c | 83.04 ± 2.51 a | 100.0 ± 0.00 a | 0.0 ± 0.00 |
| LC10 | 5.52 ± 0.07 b | 91.60 ± 0.84 a | 10.53 ± 0.59 b | 43.24 ± 4.43 b | 0.0 ± 0.00 c | 5.40 ± 0.17 c | 82.56 ± 2.40 a | 63.29 ± 10.95 b | ||
| LC30 | 5.40 ± 0.03 b | 78.07 ± 1.39 b | 9.25 ± 1.05 b | 24.72 ± 1.86 c | 19.33 ± 1.93 b | 6.22 ± 0.12 b | 56.57 ± 3.64 b | 42.40 ± 5.58 c | ||
| LC50 | 6.03 ± 0.09 a | 54.93 ± 2.78 c | 10.17 ± 0.88 b | 9.72 ± 1.32 d | 24.13 ± 1.70 b | 6.99 ± 0.14 ab | 24.24 ± 4.10 c | 18.77 ± 4.12 d | ||
| LC70 | 6.10 ± 0.08 a | 22.73 ± 2.94 d | 9.49 ± 0.45 b | 9.32 ± 0.64 d | 59.55 ± 7.23 a | 6.65 ± 0.23a | 23.47 ± 2.44 c | 5.63 ± 2.82 d | ||
| Cyantraniliprole | Control | 5.17 ± 0.12 a | 93.20 ± 2.11 a | 15.26 ± 0.57 a | 100.0 ± 0.00 a | 0.0 ± 0.00 b | 5.09 ± 0.49 a | 85.79 ± 3.35 a | 100.0 ± 0.00 a | |
| LC10 | 5.72 ± 0.07 a | 87.00 ± 3.06 a | 12.93 ± 0.93 b | 88.19 ± 3.53 b | 1.07 ± 0.27 b | 5.15 ± 0.08 a | 73.15 ± 3.20 a | 91.67 ± 8.80 a | ||
| LC30 | 5.65 ± 0.26 a | 69.40 ± 3.66 b | 9.56 ± 0.13 c | 41.64 ± 0.63 c | 4.54 ± 0.20 b | 5.14 ± 0.14 a | 51.01 ± 1.24 b | 51.83 ± 5.83 a | ||
| LC50 | 5.48 ± 0.02 a | 67.77 ± 3.39 b | 9.45 ± 0.49 c | 18.45 ± 0.75 d | 15.73 ± 2.17 a | 5.84 ± 0.20 a | 42.48 ± 7.59 b | 36.65 ± 2.96 b | ||
| LC70 | 5.60 ± 0.25 a | 38.87 ± 1.30 c | 8.41 ± 0.11 c | 15.58 ± 1.00 d | 18.15 ± 4.40 a | 5.91 ± 0.09 a | 21.51 ± 2.43 c | 21.48 ± 5.10 b | ||
| Pymetrozine | Control | 5.38 ± 0.26 ab | 92.33 ± 2.10 a | 16.41 ± 1.62 a | 100.0 ± 0.00 a | 0.0 ± 0.00 a | 5.09 ± 0.29 a | 84.63 ± 3.00 a | 100.0 ± 0.00 a | |
| LC10 | 5.23 ± 0.03 b | 84.93 ± 3.87 a | 9.19 ± 1.15 b | 45.63 ± 4.55 b | 0.0 ± 0.00 a | 5.30 ± 0.07 a | 71.99 ± 2.00 b | 77.55 ± 3.54 b | ||
| LC30 | 5.43 ± 0.09 ab | 70.33 ± 3.25 b | 7.21 ± 0.61 b | 42.19 ± 1.63 b | 0.0 ± 0.00 a | 5.46 ± 0.15 a | 62.23 ± 1.81 b | 51.90 ± 4.85 c | ||
| LC50 | 5.80 ± 0.18 ab | 56.40 ± 3.16 c | 6.44 ± 0.16 b | 18.17 ± 0.78 c | 0.0 ± 0.00 a | 5.65 ± 0.23 a | 34.78 ± 4.82 c | 37.25 ± 8.90 c | ||
| LC70 | 5.83 ± 0.19 a | 28.80 ± 4.77 d | 6.16 ± 0.02 b | 14.43 ± 1.36 c | 0.0 ± 0.00 a | 5.73 ± 0.29 a | 35.12 ± 4.35 c | 17.30 ± 2.10 d | ||
| Insecticides | Treatment | P | F1 | F2 | |||||
|---|---|---|---|---|---|---|---|---|---|
| Adult Survival Rate (%) | Adult Longevity (Day) | Fecundity (%) | Deformity Rate (%) | Developmental Period (Day) | Adult Survival Rate (%) | Fecundity (%) | Deformity Rate (%) | ||
| Spirotetramat | Control | 92.77 ± 1.63 a | 13.85 ± 0.88 a | 100.0 ± 0.0 a | 0.0 ± 0.00 d | 4.73 ± 0.29 d | 95.97 ± 1.16 a | 100.0 ± 0.0 a | 0.0 ± 0.00 |
| LC10 | 70.30 ± 2.46 b | 8.03 ± 0.27 b | 68.88 ± 6.33 b | 0.0 ± 0.00 d | 5.43 ± 0.15 c | 92.03 ± 0.52 b | 47.91 ± 3.14 b | ||
| LC30 | 40.67 ± 2.85 c | 6.64 ± 0.39 bc | 38.28 ± 2.30 c | 25.46 ± 5.22 c | 6.25 ± 0.10 b | 66.27 ± 0.90 c | 30.34 ± 2.58 c | ||
| LC50 | 23.40 ± 0.40 d | 6.36 ± 0.87 bc | 31.77 ± 7.09 cd | 51.38 ± 5.63 b | 6.85 ± 0.23 ab | 41.43 ± 1.45 d | 22.05 ± 2.07 d | ||
| LC70 | 18.07 ± 3.67 d | 5.13 ± 0.33 c | 21.69 ± 0.35 d | 63.79 ± 2.63 a | 6.65 ± 0.23 a | 13.53 ± 1.12 e | 20.47 ± 2.68 d | ||
| Cyantraniliprole | Control | 90.17 ± 0.17 a | 12.48 ± 0.75 a | 100.0 ± 0.0 a | 0.0 ± 0.00 c | 4.62 ± 0.26 b | 92.37 ± 0.78 a | 100.0 ± 0.0 a | |
| LC10 | 78.97 ± 1.71 b | 11.75 ± 0.41 b | 63.32 ± 8.17 b | 0.0 ± 0.00 c | 5.29 ± 0.07 a | 90.90 ± 0.95 a | 59.80 ± 7.75 b | ||
| LC30 | 52.73 ± 1.87 b | 9.49 ± 0.84 bc | 40.54 ± 3.40 c | 19.42 ± 2.15 b | 5.46 ± 0.15 a | 73.07 ± 1.97 b | 33.20 ± 5.41 c | ||
| LC50 | 52.17 ± 3.00 c | 7.50 ± 0.61 c | 34.17 ± 0.89 c | 14.83 ± 1.14 b | 5.65 ± 0.23 a | 61.07 ± 1.94 c | 31.50 ± 3.71 c | ||
| LC70 | 37.07 ± 4.30 c | 7.38 ± 0.36 c | 38.95 ± 7.14 c | 34.85 ± 3.24 a | 5.73 ± 0.29 a | 18.27 ± 1.75 d | 30.01 ± 2.12 c | ||
| Pymetrozine | Control | 90.20 ± 2.52 a | 13.08 ± 2.20 a | 100.0 ± 0.0 a | 0.0 ± 0.00 a | 4.68 ± 0.32 b | 92.57 ± 2.09 a | 100.0 ± 0.0 a | |
| LC10 | 67.40 ± 3.85 b | 8.45 ± 1.06 a | 57.35 ± 6.46 b | 0.0 ± 0.00 a | 5.15 ± 0.08 b | 87.83 ± 1.16 a | 40.23 ± 3.99 b | ||
| LC30 | 66.07 ± 2.72 c | 6.59 ± 0.34 ab | 40.08 ± 11.66 bc | 0.0 ± 0.00 a | 5.14 ± 0.14 b | 78.53 ± 1.79 b | 21.14 ± 1.75 c | ||
| LC50 | 37.17 ± 3.42 c | 6.16 ± 0.50 b | 30.19 ± 0.55 c | 0.0 ± 0.00 a | 5.84 ± 0.20 a | 59.80 ± 1.80 c | 27.03 ± 3.22 c | ||
| LC70 | 36.23 ± 5.06 d | 5.77 ± 0.25 b | 36.56 ± 4.02 c | 0.0 ± 0.00 a | 5.91 ± 0.09 a | 22.40 ± 1.47 d | 23.35 ± 2.83 c | ||
| Insecticides | Treatment | R0 | T | rm | λ | DT |
|---|---|---|---|---|---|---|
| Spirotetramat | LC10 | 18.01 ± 2.53 bc | 8.75 ± 0.56 b | 0.32 ± 0.02 ab | 1.37 ± 0.01 ab | 2.32 ± 0.21 a |
| LC30 | 10.15 ± 1.37 d | 8.62 ± 0.17 b | 0.25 ± 0.02 c | 1.29 ± 0.03 c | 2.84 ± 0.24 a | |
| p-value | 0.0073 | 0.6109 | 0.0171 | 0.0145 | 0.0366 | |
| Cyantraniliprole | LC10 | 24.20 ± 4.44 b | 10.41 ± 0.32 a | 0.26 ± 0.04 bc | 1.30 ± 0.01 c | 3.26 ± 1.06 a |
| LC30 | 11.12 ± 1.50 cd | 9.14 ± 0.24 b | 0.25 ± 0.02 c | 1.29 ± 0.02 c | 2.90 ± 0.11 a | |
| p-value | 0.0405 | 0.0818 | 0.9947 | 0.709 | 0.4551 | |
| Pymetrozine | LC10 | 14.32 ± 1.85 cd | 8.74 ± 0.34 b | 0.30 ± 0.01 abc | 1.35 ± 0.01 abc | 2.33 ± 0.05 a |
| LC30 | 11.82 ± 0.64 cd | 8.64 ± 0.20 b | 0.29 ± 0.04 bc | 1.33 ± 0.04 bc | 2.56 ± 0.12 a | |
| p-value | 0.2192 | 0.9601 | 0.1729 | 0.2055 | 0.1669 | |
| Control | - | 38.58 ± 3.72 a | 10.66 ± 0.58 a | 0.35 ± 0.01 a | 1.42 ± 0.02 a | 2.01 ± 0.07 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.E.; Kim, H.K.; Kim, G.H. Sublethal Effects of Spirotetramat, Cyantraniliprole, and Pymetrozine on Aphis gossypii (Hemiptera: Aphididae). Insects 2024, 15, 247. https://doi.org/10.3390/insects15040247
Kim SE, Kim HK, Kim GH. Sublethal Effects of Spirotetramat, Cyantraniliprole, and Pymetrozine on Aphis gossypii (Hemiptera: Aphididae). Insects. 2024; 15(4):247. https://doi.org/10.3390/insects15040247
Chicago/Turabian StyleKim, Se Eun, Hyun Kyung Kim, and Gil Hah Kim. 2024. "Sublethal Effects of Spirotetramat, Cyantraniliprole, and Pymetrozine on Aphis gossypii (Hemiptera: Aphididae)" Insects 15, no. 4: 247. https://doi.org/10.3390/insects15040247