
Biological Control and Habitat Management for the Control of Onion Thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae), in Onion Production in Quebec, Canada


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Sites
2.2. Pest Management Treatments
2.2.1. 2021 AAFC Site
2.2.2. 2022 AAFC Site
2.2.3. 2021 Sites 1 and 2
2.2.4. 2022 Site 1
2.3. Data Collection
2.4. Data Analysis
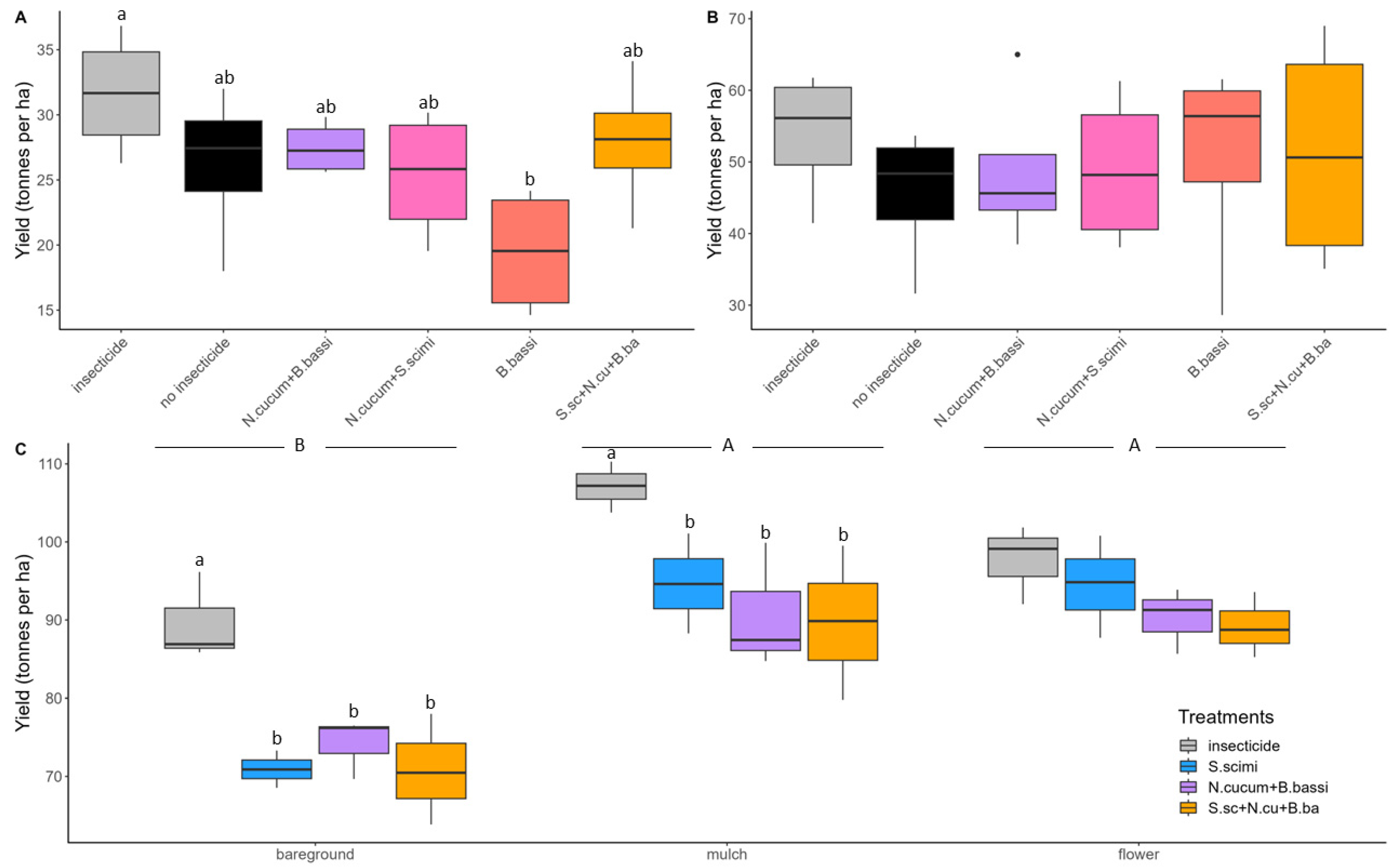
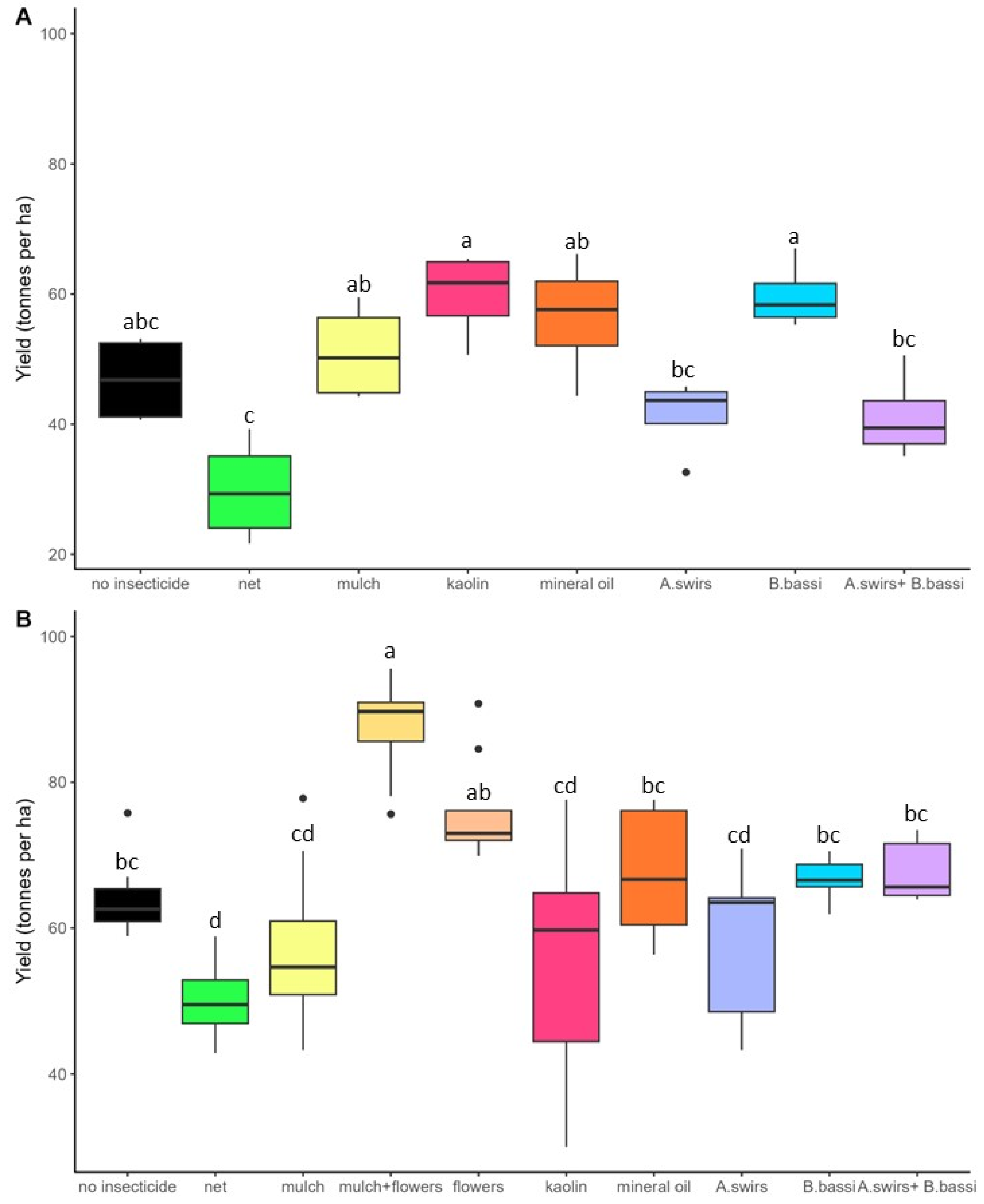
3. Results
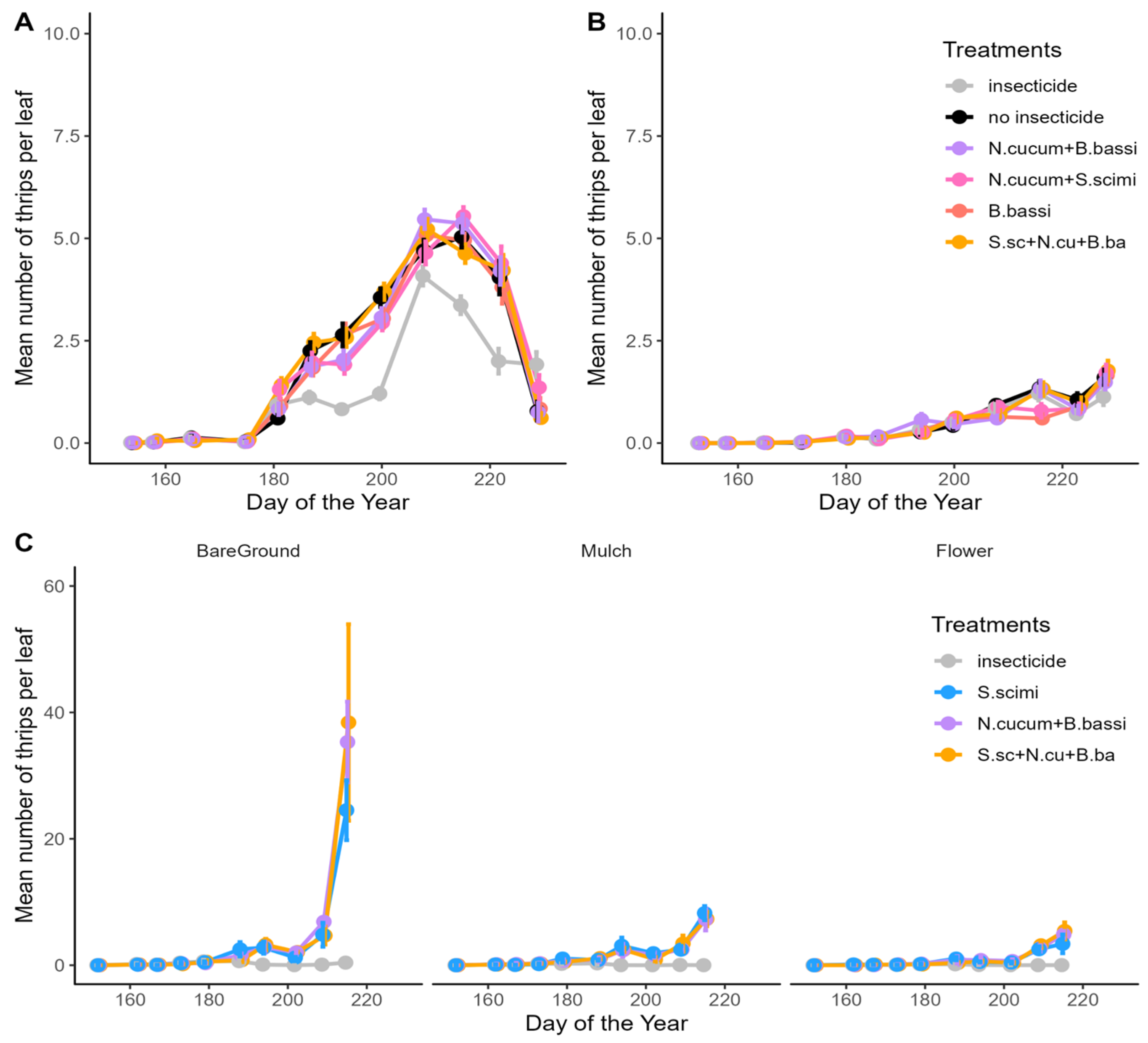
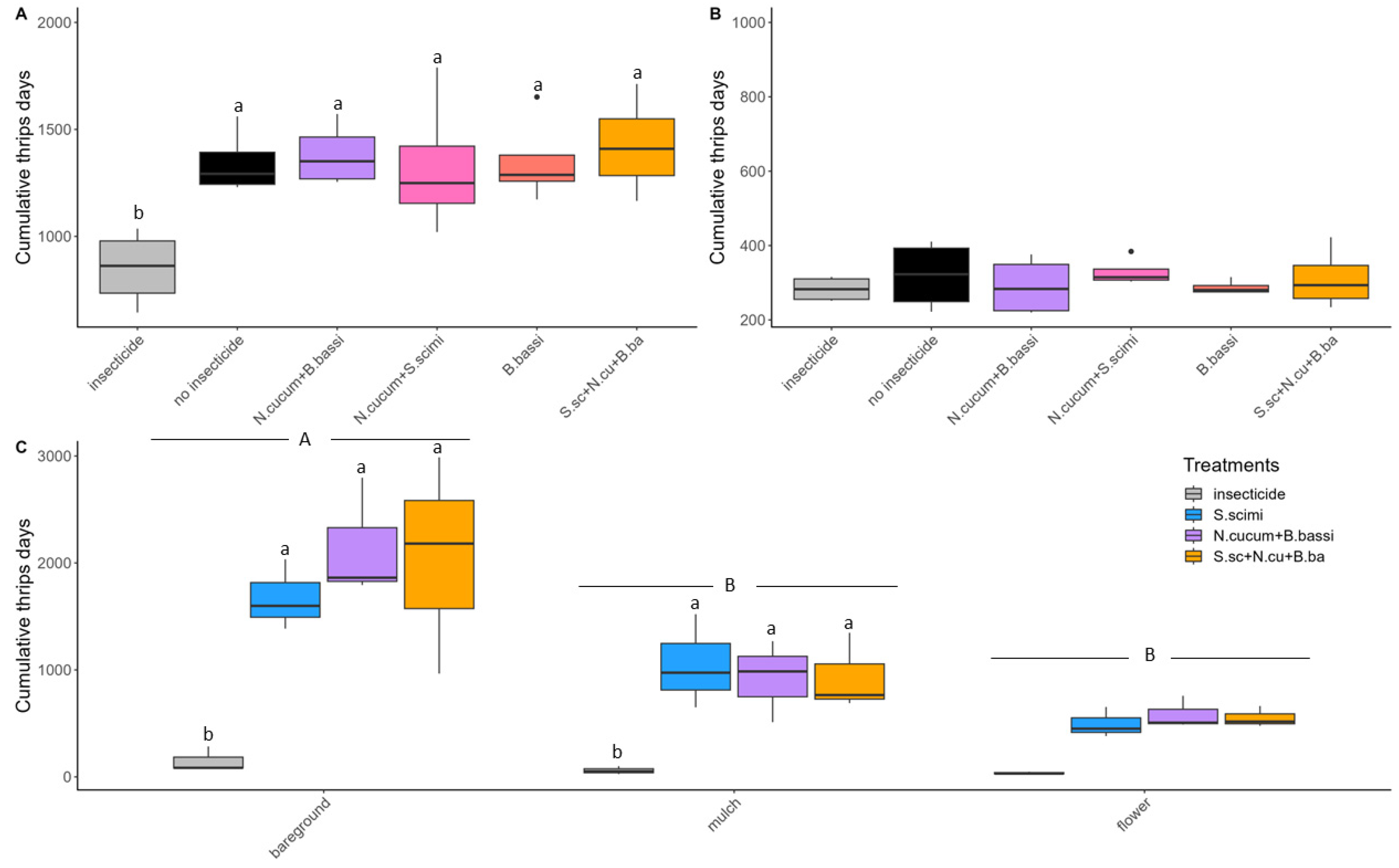
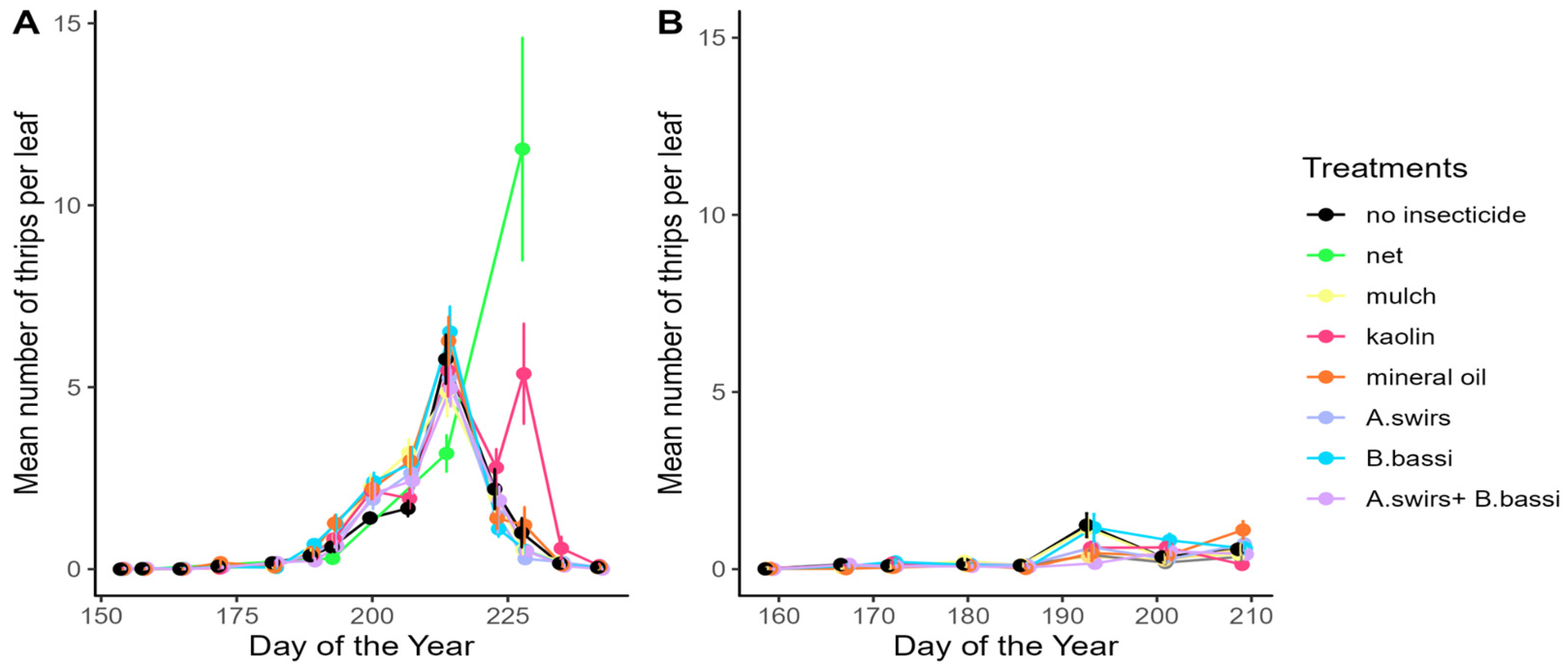
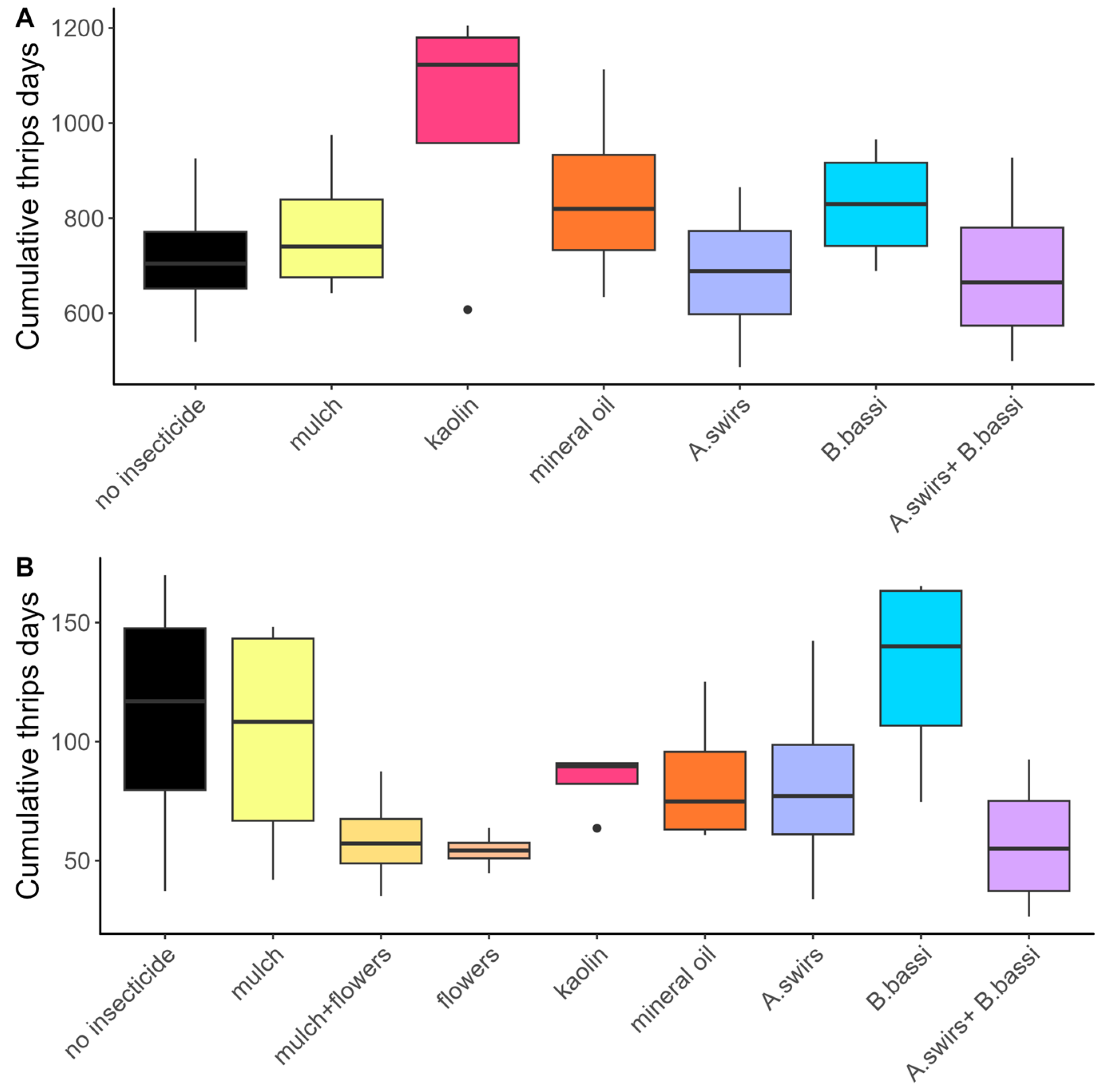
3.1. Abundance of Onion Thrips
3.2. Yield and Onion Categories
3.3. Correlation between Onion Yield and Insecticide Treatments
4. Discussion
4.1. Flower Strips and Straw Mulch: Promising Techniques for Onion Thrips Control
4.2. Biological Control Agents, Used Alone or in Combination
4.3. Products Aimed at Reducing Plant Colonization by Thrips
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- AAFC. Statistical Overview of the Canadian Field Vegetable Industry. Agriculture and Agri-Food Canada (AAFC), Government of Canada, Ottawa, Canada. 2022. Available online: https://agriculture.canada.ca/en/sector/horticulture/reports/statistical-overview-canadian-field-vegetable-industry-2022 (accessed on 22 March 2024).
- Statistics Canada. Area, Production and Farm Gate Value of Marketed Vegetables (Table 32-10-0365-01); Government of Canada: Ottawa, ON, Canada, 2022. [CrossRef]
- Diaz-Montano, J.; Fuchs, M.; Nault, B.A.; Fail, J.; Shelton, A.M. Onion thrips (Thysanoptera: Thripidae): A global pest of increasing concern in Onion. J. Econ. Entomol. 2011, 104, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gill, H.K.; Garg, H.; Gill, A.K.; Gillett-Kaufman, J.L.; Nault, B.A. Onion thrips (Thysanoptera: Thripidae) biology, ecology, and management in onion production systems. J. Integr. Pest Manag. 2015, 6, 6. [Google Scholar] [CrossRef]
- Fournier, F.; Boivin, G.; Stewart, R.K. Effect of Thrips tabaci (Thysanoptera: Thripidae) on yellow onion yields and economic thresholds for its management. J. Econ. Entomol. 1995, 88, 1401–1407. [Google Scholar] [CrossRef]
- Grode, A.S.; Brisco-McCann, E.; Wiriyajitsonboom, P.; Hausbeck, M.K.; Szendrei, Z. Managing onion thrips can limit bacterial stalk and leaf necrosis in Michigan onion fields. Plant Dis. 2019, 103, 938–943. [Google Scholar] [CrossRef] [PubMed]
- Leach, A.; Hay, F.; Harding, R.; Damann, K.C.; Nault, B. Relationship between onion thrips (Thrips tabaci) and Stemphylium vesicarium in the development of Stemphylium leaf blight in onion. Ann. Appl. Biol. 2020, 176, 55–64. [Google Scholar] [CrossRef]
- Kritzman, A.; Lampel, M.; Raccah, B.; Gera, A. Distribution and transmission of Iris yellow spot virus. Plant Dis. 2001, 85, 838–842. [Google Scholar] [CrossRef] [PubMed]
- Hoepting, C.A.; Allen, J.K.; Vanderkooi, K.D.; Hovius, M.Y.; Fuchs, M.F.; Pappu, H.R.; McDonald, M.R. First report of Iris yellow spot virus on onion in Canada. Plant Dis. 2008, 92, 318. [Google Scholar] [CrossRef]
- Larentzaki, E.; Shelton, A.M.; Musser, F.R.; Nault, B.A.; Plate, J. Overwintering locations and hosts for onion thrips (Thysanoptera: Thripidae) in the onion cropping ecosystem in New York. J. Econ. Entomol. 2007, 100, 1194–1200. [Google Scholar] [CrossRef]
- Smith, E.A.; Shields, E.J.; Nault, B.A. Impact of abiotic factors on onion thrips (Thysanoptera: Thripidae) aerial dispersal in an onion ecosystem. Environ. Entomol. 2016, 45, 1115–1122. [Google Scholar] [CrossRef]
- OMAFRA. Onion Thrips. Ontario Crop IPM. Ontario Ministry of Agriculture, Food and Rural Affairs. 2009. Available online: http://www.omafra.gov.on.ca/IPM/english/onions/insects/onion-thrips.html#advanced (accessed on 22 March 2024).
- Shelton, A.M.; Nault, B.A.; Plate, J.; Zhao, J.Z. Regional and temporal variation in susceptibility to lambda-cyhalothrin in onion thrips, Thrips tabaci (Thysanoptera: Thripidae), in onion fields in New York. J. Econ. Entomol. 2003, 96, 1843–1848. [Google Scholar] [CrossRef]
- Allen, J.K.M.; Scott-Dupree, C.D.; Tolman, J.H.; Harris, C.R. Resistance of Thrips tabaci to pyrethroid and organophosphorus insecticides in Ontario, Canada. Pest Manag. Sci. 2005, 61, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Adesanya, A.W.; Waters, T.D.; Lavine, M.D.; Walsh, D.B.; Lavine, L.C.; Zhu, F. Multiple insecticide resistance in onion thrips populations from Western USA. Pestic. Biochem. Physiol. 2020, 165, 104553. [Google Scholar] [CrossRef] [PubMed]
- Maniania, N.K.; Sithanantham, S.; Ekesi, S.; Ampong-Nyarko, K.; Baumgärtner, J.; Löhr, B.; Matoka, C.M. A field trial of the entomogenous fungus Metarhizium anisopliae for control of onion thrips, Thrips tabaci. Crop Prot. 2003, 22, 553–559. [Google Scholar] [CrossRef]
- Wu, S.; Gao, Y.; Xu, X.; Wang, E.; Wang, Y.; Lei, Z. Evaluation of Stratiolaelaps scimitus and Neoseiulus barkeri for biological control of thrips on greenhouse cucumbers. Biocontrol. Sci. Technol. 2014, 24, 1110–1121. [Google Scholar] [CrossRef]
- Bosco, L.; Tavella, L. Population dynamics and integrated pest management of Thrips tabaci on leek under field conditions in northwest Italy. Entomol. Exp. App. 2010, 135, 276–287. [Google Scholar] [CrossRef]
- Fok, E.J.; Petersen, J.D.; Nault, B.A. Relationships between insect predator populations and their prey, Thrips tabaci, in onion fields grown in large-scale and small-scale cropping systems. BioControl 2014, 59, 739–748. [Google Scholar] [CrossRef]
- Saito, T.; Brownbridge, M. Compatibility of soil-dwelling predators and microbial agents and their efficacy in controlling soil-dwelling stages of western flower thrips Frankliniella occidentalis. Biol. Control 2016, 92, 92–100. [Google Scholar] [CrossRef]
- Albrecht, M.; Kleijn, D.; Williams, N.M.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.J.; Dainese, M.; Drummond, F.A.; Entling, M.H.; et al. The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: A quantitative synthesis. Ecol. Lett. 2020, 23, 1488–1498. [Google Scholar] [CrossRef]
- Heimpel, G.E.; Jervis, M.A. Does floral nectar improve biological control by parasitoids? In Plant-Provided Food for Carnivorous Insects: A Protective Mutualism and Its Applications; Wäckers, F.L., van Rijn, P.C.J., Bruin, J., Eds.; Cambridge University Press: New York, NY, USA, 2005; pp. 267–304. [Google Scholar]
- Haenke, S.; Scheid, B.; Schaefer, M.; Tscharntke, T.; Thies, C. Increasing syrphid fly diversity and density in sown flower strips within simple vs. complex landscapes. J. Appl. Ecol. 2009, 46, 1106–1114. [Google Scholar] [CrossRef]
- Alcalá Herrera, R.; Cotes, B.; Agustí, N.; Tasin, M.; Porcel, M. Using flower strips to promote green lacewings to control cabbage insect pests. J. Pest Sci. 2022, 95, 669–683. [Google Scholar] [CrossRef]
- Masiunas, J.B. Production of vegetables using cover crop and living mulches—A review. J. Veg. Crop Prod. 1998, 4, 11–31. [Google Scholar] [CrossRef]
- Bryant, A.; Coudron, T.; Brainard, D.; Szendrei, Z. Cover crop mulches influence biological control of the imported cabbageworm (Pieris rapae L., Lepidoptera: Pieridae) in cabbage. Biol. Control 2014, 73, 75–83. [Google Scholar] [CrossRef]
- de Pedro, L.; Perera-Fernández, L.G.; López-Gallego, E.; Pérez-Marcos, M.; Sanchez, J.A. The effect of cover crops on the biodiversity and abundance of ground-dwelling arthropods in a Mediterranean pear orchard. Agronomy 2020, 10, 580. [Google Scholar] [CrossRef]
- Stavisky, J.; Funderburk, J.; Brodbeck, B.V.; Olson, S.M.; Andersen, P.C. Population dynamics of Frankliniella spp. and tomato spotted wilt incidence as influenced by cultural management tactics in tomato. J. Econ. Entomol. 2002, 95, 1216–1221. [Google Scholar] [CrossRef]
- Reitz, S.R.; Yearby, E.L.; Funderburk, J.E.; Stavisky, J.; Momol, M.T.; Olson, S.M. Integrated management tactics for Frankliniella thrips (Thysanoptera: Thripidae) in field-grown pepper. J. Econ. Entomol. 2003, 96, 1201–1214. [Google Scholar] [CrossRef]
- Larentzaki, E.; Plate, J.; Nault, B.A.; Shelton, A.M. Impact of straw mulch on populations of onion thrips (Thysanoptera: Thripidae) in onion. J. Econ. Entomol. 2008, 101, 1317–1324. [Google Scholar] [CrossRef]
- Iglesias, L.; Havey, M.J.; Nault, B.A. Management of onion thrips (Thrips tabaci) in organic onion production using multiple IPM tactics. Insects 2021, 12, 207. [Google Scholar] [CrossRef]
- Puterka, G.J.; Glenn, D.M.; Sekutowski, D.G.; Unruh, T.R.; Jones, S.K. Progress toward liquid formulations of particle films for insect and disease control in pear. Environ. Entomol. 2000, 29, 329–339. [Google Scholar] [CrossRef]
- Amalin, D.M.; Averion, L.; Bihis, D.; Legaspi, J.C.; David, E.F. Effectiveness of kaolin clay particle film in managing Helopeltis collaris (Hemiptera: Miridae), a major pest of cacao in the Philippines. Fla. Entomol. 2015, 98, 354–355. [Google Scholar] [CrossRef]
- Sharma, R.R.; Datta, S.C.; Varghese, E. Kaolin-based particle film sprays reduce the incidence of pests, diseases and storage disorders and improve postharvest quality of ‘Delicious’ apples. Crop Prot. 2020, 127, 104950. [Google Scholar] [CrossRef]
- Oliveira, D.F.; Benhadi-Marín, J.; Neto, J.; Sanz, L.; Garzo, E.; Aguiar, A.; Fereres, A.; Pereira, J.A. Kaolin particle films disrupt landing, settling behavior and feeding of Trioza erytrae on lemon plants. Pest Manag. Sci. 2022, 78, 4753–4763. [Google Scholar] [CrossRef] [PubMed]
- Larentzaki, E.; Shelton, A.M.; Plate, J. Effect of kaolin particle film on Thrips tabaci (Thysanoptera: Thripidae), oviposition, feeding and development on onions: A lab and field case study. Crop Prot. 2008, 27, 727–734. [Google Scholar] [CrossRef]
- Yang, Q.; Arthurs, S.; Lu, Z.; Liang, Z.; Mao, R. Use of horticultural mineral oils to control potato virus Y (PVY) and other non-persistent aphid-vectored viruses. Crop Prot. 2019, 118, 97–103. [Google Scholar] [CrossRef]
- Tansey, J.A.; Jones, M.M.; Vanaclocha, P.; Robertson, J.; Stansly, P.A. Costs and benefits of frequent low-volume applications of horticultural mineral oil for management of Asian citrus psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae). Crop Prot. 2015, 76, 59–67. [Google Scholar] [CrossRef]
- Buteler, M.; Stadler, T. A review on the mode of action and current use of petroleum distilled spray oils. In Pesticides in the Modern World—Pesticides Use and Management; Stoytcheva, M., Ed.; InTech: Rijeka, Croatia, 2011; pp. 119–136. [Google Scholar]
- Chouinard, G.; Pelletier, F.; Larose, M.; Knoch, S.; Pouchet, C.; Dumont, M.J.; Tavares, J.R. Insect netting: Effect of mesh size and shape on exclusion of some fruit pests and natural enemies under laboratory and orchard conditions. J. Pest Sci. 2023, 96, 857–869. [Google Scholar] [CrossRef]
- Böckmann, E. Effects of insect net coverage in field vegetables on pests, diseases, natural enemies, and yield. J. Plant Dis. Prot. 2022, 129, 1401–1415. [Google Scholar] [CrossRef]
- CRAAQ. Reference Guide for Crop Fertilization; Centre de Référence en Agriculture et Agroalimentaire du Québec: Quebec, QC, Canada, 2013; 473p. [Google Scholar]
- Park, J.; Mostafiz, M.M.; Hwang, H.S.; Jung, D.O.; Lee, K.Y. Comparison of the predation capacities of two soil-dwelling predatory mites, Gaeolaelaps aculeifer and Stratiolaelaps scimitus (Acari: Laelapidae), on three thrips species. J. Asia Pac. Entomol. 2021, 24, 397–401. [Google Scholar] [CrossRef]
- Bakker, F.M.; Sabelis, M.W. How larvae of Thrips tabaci reduce the attack success of phytoseiid predators. Entomol. Exp. App. 1989, 50, 47–51. [Google Scholar] [CrossRef]
- Lin, G.; Tanguay, A.; Guertin, C.; Todorova, S.; Brodeur, J. A new method for loading predatory mites with entomopathogenic fungi for biological control of their prey. Biol. Control 2017, 115, 105–111. [Google Scholar] [CrossRef]
- Lin, G.; Guertin, C.; Di Paolo, S.A.; Todorova, S.; Brodeur, J. Phytoseiid predatory mites can disperse entomopathogenic fungi to prey patches. Sci. Rep. 2019, 9, 19435. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Sun, L.; Lin, G.Y.; Lin, J.Z.; Chen, X.; Ji, J.; Zhi-Qiang, Z.; Saito, Y. A novel use of predatory mites for dissemination of fungal pathogen for insect biocontrol: The case of Amblyseius swirskii and Neoseiulus cucumeris (Phytoseiidae) as vectors of Beauveria bassiana against Diaphorina citri (Psyllidae). Syst. Appl. Acarol. 2015, 20, 177–187. [Google Scholar] [CrossRef]
- Fernandes, E.K.; Rangel, D.E.; Moraes, A.M.; Bittencourt, V.R.; Roberts, D.W. Variability in tolerance to UV-B radiation among Beauveria spp. isolates. J. Invertebr. Pathol. 2007, 96, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Surdek, N.; Lacoursière, E.; Gendron, M.; Lapointe, A. Recueil des Seuils D’intervention Contre les Insectes et Maladies en Cultures Maraîchères Édition 2021; Groupe Pleine Terre: Quebec, QC, Canada, 2021; 256p. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing (Version 4.2.2); R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: http://www.r-project.org/ (accessed on 22 March 2024).
- Gurr, G.M.; Wratten, S.D.; Landis, D.A.; You, M. Habitat management to suppress pest populations: Progress and prospects. Annu. Rev. Entomol. 2017, 62, 91–109. [Google Scholar] [CrossRef] [PubMed]
- Sekine, T.; Osaka, M.; Itabashi, T.; Chiba, N.; Yoshimura, H.; Uesugi, R.; Tabuchi, K.; Shimoda, T. Predation of syrphid larvae (Diptera: Syrphidae) on thrips in onion fields intercropped with barley. Appl. Entomol. Zool. 2022, 57, 305–311. [Google Scholar] [CrossRef]
- Yang, Q.; Men, X.; Zhao, W.; Li, C.; Zhang, Q.; Cai, Z.; Ge, F.; Ouyang, F. Flower strips as a bridge habitat facilitate the movement of predatory beetles from wheat to maize crops. Pest Manag. Sci. 2021, 77, 1839–1850. [Google Scholar] [CrossRef]
- Gowton, C.M.; Cabra-Arias, C.; Carrillo, J. Intercropping with peppermint increases ground dwelling insect and pollinator abundance and decreases Drosophila suzukii in fruit. Front. Sustain. Food Syst. 2021, 5, 700842. [Google Scholar] [CrossRef]
- Terry, L.I. Host selection, communication and reproductive behavior. In Thrips as Crop Pests; Lewis, T., Ed.; CABI International: London, UK, 1997; pp. 65–118. [Google Scholar]
- Teulon, D.A.; Penman, D.R.; Ramakers, P.M. Volatile chemicals for thrips (Thysanoptera: Thripidae) host finding and applications for thrips pest management. J. Econ. Entomol. 1993, 86, 1405–1415. [Google Scholar] [CrossRef]
- van Tol, R.W.; James, D.E.; De Kogel, W.J.; Teulon, D.A. Plant odours with potential for a push–pull strategy to control the onion thrips, Thrips tabaci. Entomol. Exp. Appl. 2007, 122, 69–76. [Google Scholar] [CrossRef]
- Windsor, F.M.; Tavella, J.; Rother, D.C.; Raimundo, R.L.; Devoto, M.; Guimarães, P.R., Jr.; Evans, D.M. Identifying plant mixes for multiple ecosystem service provision in agricultural systems using ecological networks. J. Appl. Ecol. 2021, 58, 2770–2782. [Google Scholar] [CrossRef]
- Gardarin, A.; Plantegenest, M.; Bischoff, A.; Valantin-Morison, M. Understanding plant–arthropod interactions in multitrophic communities to improve conservation biological control: Useful traits and metrics. J. Pest Sci. 2018, 91, 943–955. [Google Scholar] [CrossRef]
- Hatt, S.; Francis, F.; Xu, Q.; Wang, S.; Osawa, N. Perennial flowering strips for conservation biological control of insect pests: From picking and mixing flowers to tailored functional diversity. In Integrative Biological Control: Ecostacking for Enhanced Ecosystem Services; Gao, Y., Hokkanen, H.M.T., Menzler-Hokkanen, I., Eds.; Springer: Cham, Switzerland, 2020; pp. 57–71. [Google Scholar]
- Johnson, J.M.; Hough-Goldstein, J.A.; Vangessel, M.J. Effects of straw mulch on pest insects, predators, and weeds in watermelons and potatoes. Environ. Entomol. 2004, 33, 1632–1643. [Google Scholar] [CrossRef]
- Trdan, S.; Žnidarčič, D.; Kač, M.; Vidrih, M. Yield of early white cabbage grown under mulch and non-mulch conditions with low populations of onion thrips (Thrips tabaci Lindeman). Int. J. Pest Manag. 2008, 54, 309–318. [Google Scholar] [CrossRef]
- Iqbal, R.; Raza, M.A.S.; Valipour, M.; Saleem, M.F.; Zaheer, M.S.; Ahmad, S.; Toleikiene, M.; Haider, I.; Aslam, M.U.; Nazar, M.A. Potential agricultural and environmental benefits of mulches—A review. Bull. Natl. Res. Cent. 2020, 44, 1–16. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Basit, A.; Mohamed, H.I.; Ali, I.; Ullah, S.; Kamel, E.A.; Shalaby, T.A.; Ramadan, K.M.A.; Alkhateeb, A.A.; Ghazzawy, H.S. Mulching as a sustainable water and soil saving practice in agriculture: A review. Agronomy 2022, 12, 1881. [Google Scholar] [CrossRef]
- Moore, E.; Wiedenhoeft, M.; Kaspar, T.; Cambardella, C.A. Rye cover crop effects on soil quality in no-till corn silage–soybean cropping systems. Soil Sci. Soc. Am. J. 2014, 78, 968. [Google Scholar] [CrossRef]
- Barnes, J.P.; Putnam, A.R. Rye residues contribute weed suppression in no-tillage cropping systems. J. Chem. Ecol. 1983, 9, 1045–1057. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, H.F.; Gent, D.H.; Fichtner, S.M.; Hammon, R.; Cranshaw, W.S.; Mahaffey, L.; Camper, M.; Otto, K.; McMillan, M. Straw mulch and reduced-risk pesticide impacts on thrips and Iris yellow spot virus on western-grown onions. Southwest. Entomol. 2009, 34, 13–29. [Google Scholar] [CrossRef]
- Lu, F.M. Color preference and using silver mulches to control the onion thrips, Thrips tabaci Lindeman. Chin. J. Entomol. 1990, 10, 337–342. [Google Scholar]
- Till, C.; James, D.E.; Teulon, D.A.J. Evaluation of UV reflective mulches for protection against thrips (Thrips tabaci) in onion (Allium cepa) crops. N. Z. Plant Prot. 2004, 57, 209–213. [Google Scholar] [CrossRef]
- Perez-Alvarez, R.; Nault, B.A.; Poveda, K. Effectiveness of augmentative biological control depends on landscape context. Sci. Rep. 2019, 9, 8664. [Google Scholar] [CrossRef]
- van Lenteren, J.C.; Bueno, V.H.; Klapwijk, J.N. Augmentative biological control. In Biological Control: Global Impacts, Challenges and Future Directions of Pest Management; Mason, P.G., Ed.; CSIRO Publishing: Clayton South, Australia, 2021; pp. 90–109. [Google Scholar]
- Wu, S.; Gao, Y.; Xu, X.; Zhang, Y.; Wang, J.; Lei, Z.; Smagghe, G. Laboratory and greenhouse evaluation of a new entomopathogenic strain of Beauveria bassiana for control of the onion thrips Thrips tabaci. Biocontrol Sci. Technol. 2013, 23, 794–802. [Google Scholar] [CrossRef]
- Jung, D.O.; Hwang, H.S.; Kim, S.Y.; Lee, K.Y. Biological control of thrips using a self-produced predatory mite Stratiolaelaps scimitus (Acari: Laelapidae) in the greenhouse chrysanthemum. Korean J. Appl. Entomol. 2019, 58, 233–238. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, S.; Reitz, S.R.; Gao, Y. Simultaneous application of entomopathogenic Beauveria bassiana granules and predatory mites Stratiolaelaps scimitus for control of western flower thrips, Frankliniella occidentalis. J. Pest Sci. 2021, 94, 119–127. [Google Scholar] [CrossRef]
- Ebssa, L.; Borgemeister, C.; Poehling, H.M. Simultaneous application of entomopathogenic nematodes and predatory mites to control western flower thrips Frankliniella occidentalis. Biol. Control 2006, 39, 66–74. [Google Scholar] [CrossRef]
- Wimmer, D.; Hoffmann, D.; Schausberger, P. Prey suitability of western flower thrips, Frankliniella occidentalis, and onion thrips, Thrips tabaci, for the predatory mite Amblyseius swirskii. Biocontrol Sci. Technol. 2008, 18, 533–542. [Google Scholar] [CrossRef]
- Manners, A.G.; Dembowski, B.R.; Healey, M.A. Biological control of western flower thrips, Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae), in gerberas, chrysanthemums and roses. Aust. J. Entomol. 2013, 52, 246–258. [Google Scholar] [CrossRef]
- Shipp, J.L.; Van Houten, Y.M. Influence of temperature and vapor pressure deficit on survival of the predatory mite Amblyseius cucumeris (Acari: Phytoseiidae). Environ. Entomol. 1997, 26, 106–113. [Google Scholar] [CrossRef]
- Sabelis, M.W.; Van Rijn, P.C. Predation by insects and mites. In Thrips as Crop Pests; Lewis, T., Ed.; CABI International: London, UK, 1997; pp. 259–354. [Google Scholar]
- Summerfield, A. Integrated Pest Management of Onion Thrips (Thrips tabaci), an Emerging Pest of Greenhouse Floriculture Crops in Ontario. Master’s Thesis, University of Guelph, Guelph, ON, Canada, 2022; 178p. [Google Scholar]
- Ansari, M.A.; Brownbridge, M.; Shah, F.A.; Butt, T.M. Efficacy of entomopathogenic fungi against soil-dwelling life stages of western flower thrips, Frankliniella occidentalis, in plant-growing media. Entomol. Exp. App. 2008, 127, 80–87. [Google Scholar] [CrossRef]
- Jacobson, R.J.; Chandler, D.; Fenlon, J.; Russell, K.M. Compatibility of Beauveria bassiana (balsamo) Vuillemin with Amblyseius cucumeris Oudemans (Acarina: Phytoseiidae) to control Frankliniella occidentalis Pergande (Thysanoptera: Thripidae) on cucumber plants. Biocontrol Sci. Technol. 2001, 11, 391–400. [Google Scholar] [CrossRef]
- Rosenheim, J.A.; Harmon, J.P. The influence of intraguild predation on the suppression of a shared prey population: An empirical reassessment. In Trophic and Guild in Biological Interactions Control; Brodeur, J., Boivin, G., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 1–20. [Google Scholar]
- Denno, R.F.; Finke, D.L. Multiple predator interactions and food-web connectance: Implications for biological control. In Trophic and Guild in Biological Interactions Control; Brodeur, J., Boivin, G., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 45–70. [Google Scholar]
- Denoth, M.; Frid, L.; Myers, J.H. Multiple agents in biological control: Improving the odds? Biol. Control 2002, 24, 20–30. [Google Scholar] [CrossRef]
- Alhadidi, S.N.; Griffin, J.N.; Fowler, M.S. Natural enemy composition rather than richness determines pest suppression. BioControl 2018, 63, 575–584. [Google Scholar] [CrossRef]
- El-Aziz, A.S.E. Laboratory and field evaluation of kaolin and bentonite particle films against onion thrips, Thrips tabaci (Lind.) (Thysanoptera: Thripidae) on onion plants. J. Appl. Sci. Res. 2013, 9, 3141–3145. [Google Scholar]
- Singh, H.; Cheema, H.K.; Singh, R. Field evaluation of horticultural mineral oils and botanicals against bean thrips, Megalurothrips distalis (Karny) (Thysanoptera: Thripidae), in summer mung bean. Egypt. J. Biol. Pest Control 2020, 30, 1–8. [Google Scholar] [CrossRef]
- Moriyama, T.; Hayashi, M.; Ide, O. Study on choice and the index for insect barrier net superiority in air transmissivity. Bull. Fukuoka Agri. Res. Center 2008, 27, 99–103. [Google Scholar]
- Nagasaka, K.; Kumakura, H.; Tanaka, K.; Nakagawa, I.; Ojima, K. Effects of insect-proof screens in vegetable production. Plant Prot. 2003, 57, 169–173. [Google Scholar]
- Ohya, T.; Suzuki, M.; Mitsunaga, T.; Ohta, K.; Abe, H.; Nakamura, A.; Uekusa, H.; Itoyama, K. Effect of covering with red insect screen to control Thrips tabaci (Thysanoptera: Thripidae) on cabbage. Appl. Entomol. Zool. 2022, 57, 55–62. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gagnon, A.-È.; Fortier, A.-M.; Audette, C. Biological Control and Habitat Management for the Control of Onion Thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae), in Onion Production in Quebec, Canada. Insects 2024, 15, 232. https://doi.org/10.3390/insects15040232
Gagnon A-È, Fortier A-M, Audette C. Biological Control and Habitat Management for the Control of Onion Thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae), in Onion Production in Quebec, Canada. Insects. 2024; 15(4):232. https://doi.org/10.3390/insects15040232
Chicago/Turabian StyleGagnon, Annie-Ève, Anne-Marie Fortier, and Carolane Audette. 2024. "Biological Control and Habitat Management for the Control of Onion Thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae), in Onion Production in Quebec, Canada" Insects 15, no. 4: 232. https://doi.org/10.3390/insects15040232