
Diversity of Parasitoid Wasps and Comparison of Sampling Strategies in Rice Fields Using Metabarcoding

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Storage
2.2. DNA Extraction and Amplification
2.3. High Throughput Sequencing and Data Analyses
3. Results
3.1. OTU Taxonomic Assignment
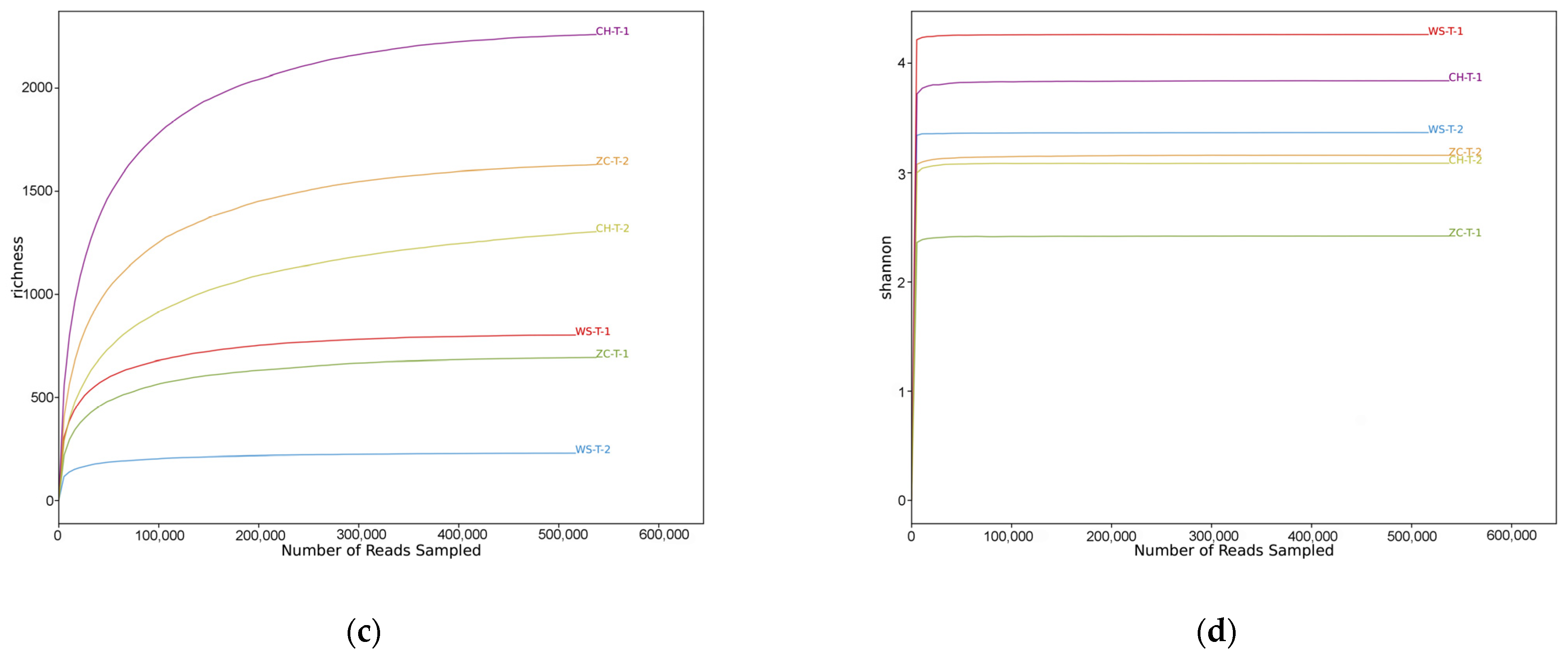
- Ethanol samples: After splicing and removing impurities, 7,391,894 sequences were obtained by high-throughput sequencing, with a total of 2,311,835,324 bp and an average fragment size of 312.753 bp. A total of 243,783 OTUs were obtained by removing chimera and repeat sequences. The OTU abundance in the 12 samples was flattened out for subsequent analysis. The coverage of 12 samples was between 0.93–0.99 (Table S1), indicating that the depth of sequencing was reasonable and fully reflected the richness of samples. Both the rarefaction curve (Figure 1a) and Shannon–Wiener curve (Figure 1b) tended to be flat, indicating that the sequencing data were large enough to reflect most of the biological information in the sample.
- Tissue samples: A total of 2,444,490 sequences were obtained by high-throughput sequencing with a total of 745,281,495 bp and an average fragment size of 304.88 bp. A total of 4446 OTUs were obtained. The OTU abundance of four samples was flattened out for subsequent analysis. The coverage of four samples was greater than 0.999 (Table S1), indicating that the depth of sequencing was reasonable and fully reflected the richness of samples. Both the rarefaction curve (Figure 1c) and Shannon–Wiener curve (Figure 1d) tended to be flat.
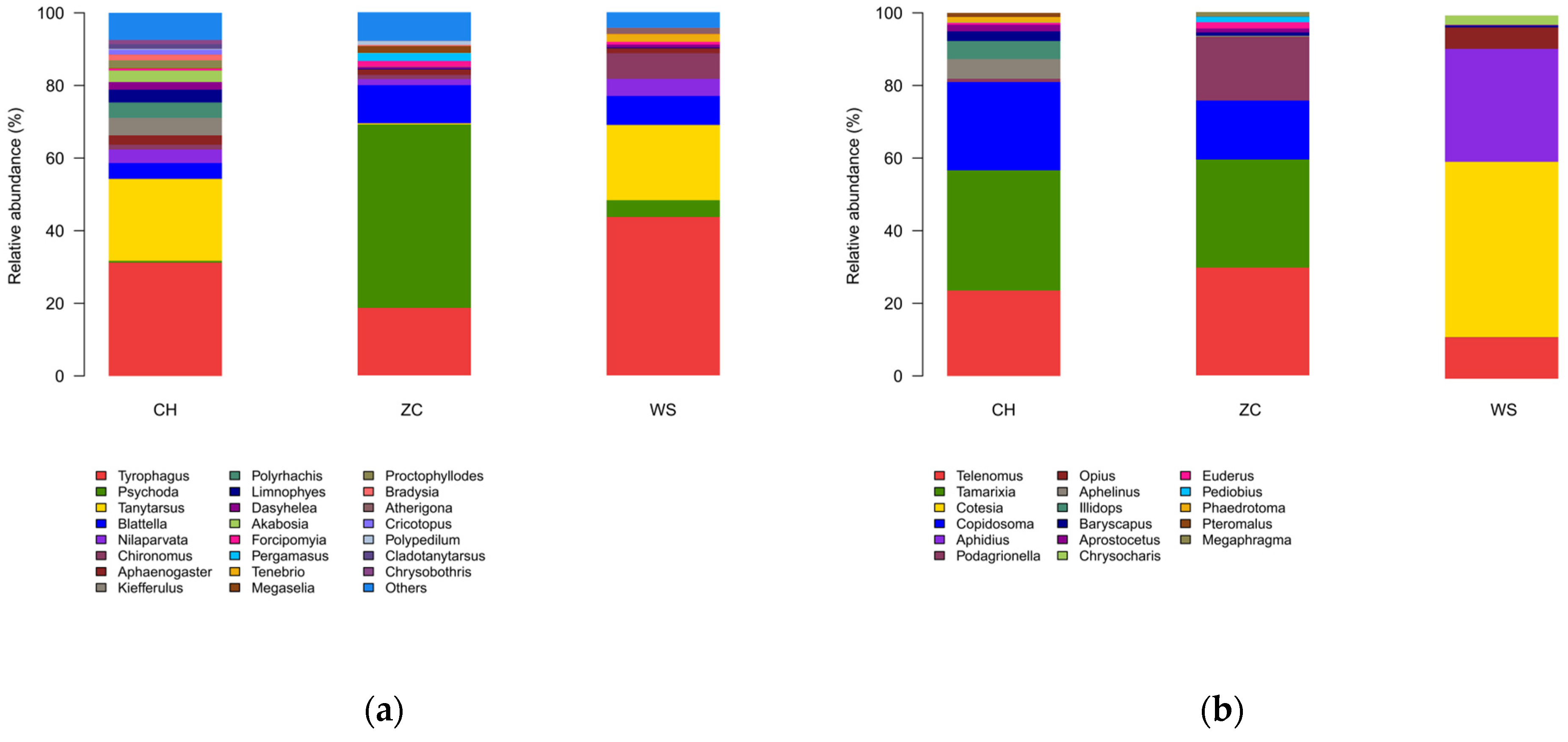
3.2. Species Composition Analysis
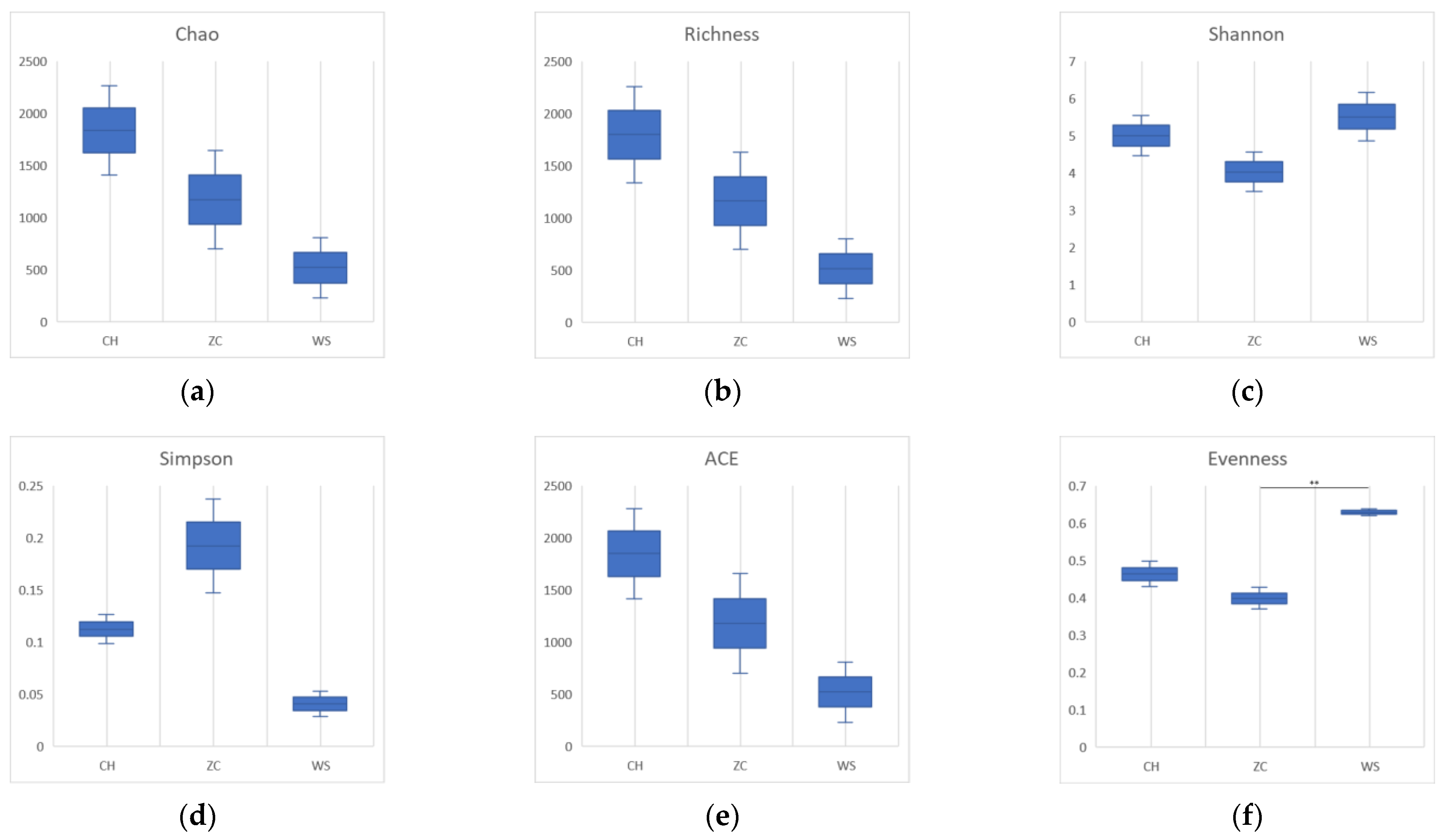
3.3. Group Comparison Analyses
3.4. Differences in Species Diversity of Parasitoid Wasps Noted in the Two Treatments
4. Discussion
4.1. Diversity of Parasitoid Wasp Communities
4.2. Limitations of Ethanol DNA Extraction Methods
4.3. Species Annotation and Abundance
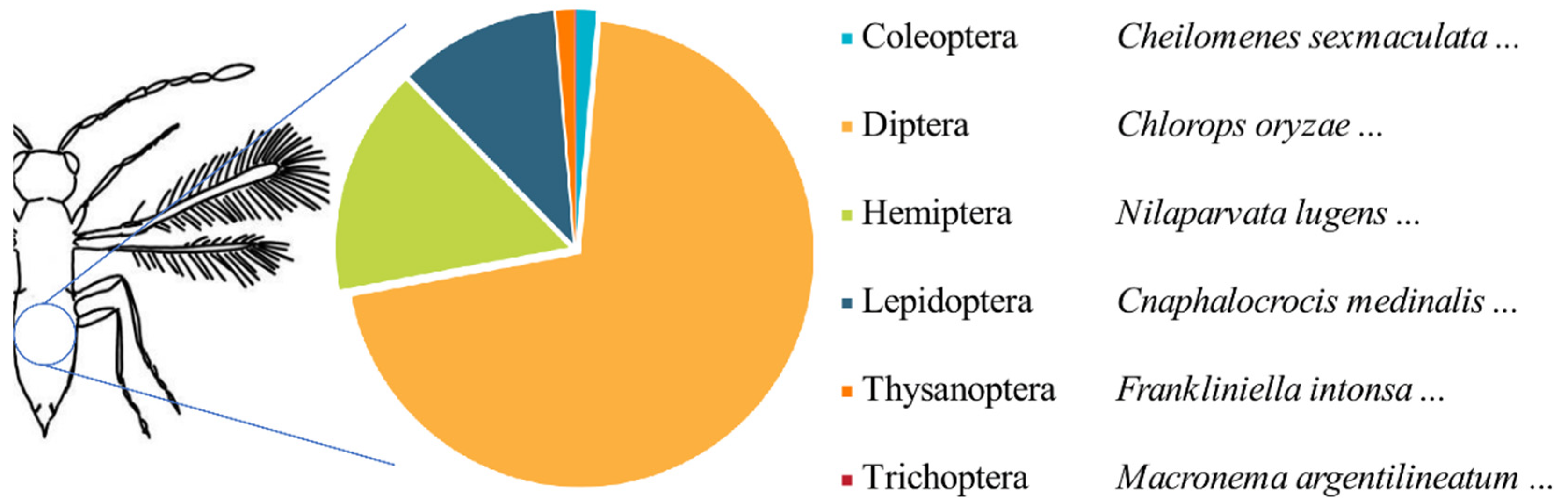
4.4. Potential Trophic Network of Parasitoid Wasps
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shields, M.W.; Johnson, A.C.; Pandey, S.; Cullen, R.; González-Chang, M.; Wratten, S.D.; Gurr, G.M. History, current situation and challenges for conservation biological control. Biol. Control 2019, 131, 25–35. [Google Scholar] [CrossRef]
- Holland, J.M.; Bianchi, F.J.J.A.; Entling, M.H.; Moonen, A.C.; Smith, B.M.; Jeanneret, P. Structure, function and management of semi-natural habitats for conservation biological control: A review of European studies. Pest Manag. Sci. 2016, 72, 1638–1651. [Google Scholar] [CrossRef] [PubMed]
- Gurr, G.M.; Liu, J.; Read, D.M.Y.; Catindig, J.L.A.; Cheng, J.A.; Lan, L.P.; Heong, K.L. Parasitoids of Asian rice planthopper (Hemiptera: Delphacidae) pests and prospects for enhancing biological control by ecological engineering. Ann. Appl. Biol. 2011, 158, 149–176. [Google Scholar] [CrossRef]
- Zang, L.S.; Wang, S.; Zhang, F.; Desneux, N. Biological control with Trichogramma in China: History, present status, and perspectives. Annu. Rev. Entomol. 2021, 66, 463–484. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Huang, T.F.; Tang, B.J.; Wang, B.Y.; Wang, L.Y.; Liu, J.B.; Zhou, Q. Transcriptome analysis and molecular characterization of soluble chemical communication proteins in the parasitoid wasp Anagrus nilaparvatae (Hymenoptera: Mymaridae). Ecol. Evol. 2022, 12, e8661. [Google Scholar] [CrossRef] [PubMed]
- Gurr, G.M.; Lu, Z.; Zheng, X.; Xu, H.; Zhu, P.; Chen, G.; Yao, X.; Cheng, J.; Zhu, Z.; Catindig, J.L.; et al. Multi-country evidence that crop diversification promotes ecological intensification of agriculture. Nat. Plants 2016, 2, 16014. [Google Scholar] [CrossRef]
- Drechsler, M.; Settele, J. Predator-prey interactions in rice ecosystems: Effects of guild composition, trophic relationships, and land use changes—A model study exemplified for Philippine rice terraces. Ecol. Model. 2001, 137, 135–159. [Google Scholar] [CrossRef]
- Dominik, C.; Seppelt, R.; Horgan, F.G.; Settele, J.; Vaclavik, T. Landscape heterogeneity filters functional traits of rice arthropods in tropical agroecosystems. Ecol. Appl. 2022, 32, e2560. [Google Scholar] [CrossRef] [PubMed]
- Horgan, F.G.; Crisol Martinez, E.; Stuart, A.M.; Bernal, C.C.; de Cima Martin, E.; Almazan, M.L.P.; Ramal, A.F. Effects of vegetation strips, fertilizer levels and varietal resistance on the integrated management of Arthropod biodiversity in a tropical rice ecosystem. Insects 2019, 10, 328. [Google Scholar] [CrossRef]
- Sattler, C.; Gianuca, A.T.; Schweiger, O.; Franzén, M.; Settele, J. Pesticides and land cover heterogeneity affect functional group and taxonomic diversity of arthropods in rice agroecosystems. Agric. Ecosyst. Environ. 2020, 297, 106927. [Google Scholar] [CrossRef]
- Priyadarshana, T.S.; Lee, M.B.; Ascher, J.S.; Qiu, L.; Goodale, E. Crop heterogeneity is positively associated with beneficial insect diversity in subtropical farmlands. J. Appl. Ecol. 2021, 58, 2747–2759. [Google Scholar] [CrossRef]
- Schoenly, K.G.; Barrion, A.T. Designing standardized and optimized surveys to assess invertebrate biodiversity in tropical irrigated rice using structured inventory and species richness models. Environ. Entomol. 2016, 45, 446–464. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Zhang, Q.L.; Liu, Q.S.; Meissle, M.; Yang, Y.; Wang, Y.N.; Hua, H.X.; Chen, X.P.; Peng, Y.F.; Romeis, J. Rice in China—Focusing the nontarget risk assessment. Plant Biotechnol. J. 2017, 15, 1340–1345. [Google Scholar] [CrossRef] [PubMed]
- He, J.C.; Hu, Y.; Zhang, M.; We, Q.; Li, B.; He, Y.T.; Wan, P.J.; Lai, F.X.; Wang, W.X.; Yu, W.J.; et al. Community structure and diversity of wasps which parasitize hemipteran pests in the rice-growing region of southern China. Chin. J. Appl. Entomol. 2022, 59, 1096–1108. [Google Scholar]
- Triapitsyn, S.V.; Rugman-Jones, P.F.; Tretiakov, P.S.; Shih, H.T.; Huang, S.H. New synonymies in the Anagrus incarnatus Haliday ‘species complex’ (Hymenoptera: Mymaridae) including a common parasitoid of economically important planthopper (Hemiptera: Delphacidae) pests of rice in Asia. J. Nat. Hist. 2018, 52, 2795–2822. [Google Scholar] [CrossRef]
- Wijerathna, T.; Gunathilaka, N. Morphological identification keys for adults of sand flies (Diptera: Psychodidae) in Sri Lanka. Parasites Vectors 2020, 13, 450. [Google Scholar] [CrossRef] [PubMed]
- Habel, J.C.; Ulrich, W.; Segerer, A.H.; Greifenstein, T.; Knubben, J.; Morinière, J.; Bozicevic, V.; Günter, A.; Hausmann, A. Insect diversity in heterogeneous agro-environments of Central Europe. Biodivers. Conserv. 2023, 32, 4665–4678. [Google Scholar] [CrossRef]
- Holmquist, A.J.; Adams, S.A.; Gillespie, R.G. Invasion by an ecosystem engineer changes biotic interactions between native and non-native taxa. Ecol. Evol. 2023, 13, e9820. [Google Scholar] [CrossRef]
- Simon, C.; Frati, F.; Andrew, B.; Bernie, C.; Hong, L. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 652–701. [Google Scholar] [CrossRef]
- Sann, C.; Wemheuer, F.; Beaurepaire, A.; Daniel, R.; Erler, S.; Vidal, S. Preliminary investigation of species diversity of rice hopper parasitoids in southeast Asia. Insects 2018, 9, 19. [Google Scholar] [CrossRef]
- Piper, A.M.; Batovska, J.; Cogan, N.O.I.; Weiss, J.; Cunningham, J.P.; Rodoni, B.C.; Blacket, M.J. Prospects and challenges of implementing DNA metabarcoding for high-throughput insect surveillance. Gigascience 2019, 8, giz092. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.Q.; Ashton, L.; Pedley, S.M.; Edwards, D.P.; Tang, Y.; Nakamura, A.; Kitching, R.; Dolman, P.M.; Woodcock, P.; Edwards, F.A.; et al. Reliable, verifiable and efficient monitoring of biodiversity via metabarcoding. Ecol. Lett. 2013, 16, 1245–1257. [Google Scholar] [CrossRef] [PubMed]
- Gibson, J.; Shokralla, S.; Porter, T.M.; King, I.; van Konynenburg, S.; Janzen, D.H.; Hallwachs, W.; Hajibabaei, M. Simultaneous assessment of the macrobiome and microbiome in a bulk sample of tropical arthropods through DNA metasystematics. Proc. Natl. Acad. Sci. USA 2014, 111, 8007–8012. [Google Scholar] [CrossRef] [PubMed]
- Hajibabaei, M.; Spall, J.L.; Shokralla, S.; van Konynenburg, S. Assessing biodiversity of a freshwater benthic macroinvertebrate community through non-destructive environmental barcoding of DNA from preservative ethanol. BMC Ecol. 2012, 12, 28. [Google Scholar] [CrossRef] [PubMed]
- Erdozain, M.; Thompson, D.G.; Porter, T.M.; Kidd, K.A.; Kreutzweiser, D.P.; Sibley, P.K.; Swystun, T.; Chartrand, D.; Hajibabaei, M. Metabarcoding of storage ethanol vs. conventional morphometric identification in relation to the use of stream macroinvertebrates as ecological indicators in forest management. Ecol. Indic. 2019, 101, 173–184. [Google Scholar] [CrossRef]
- Ali, M.P.; Kabir, M.M.M.; Haque, S.S.; Afrin, S.; Ahmed, N.; Pittendrigh, B.; Qin, X.H. Surrounding landscape influences the abundance of insect predators in rice field. BMC Zool. 2020, 5, 8. [Google Scholar] [CrossRef]
- Koh, C.N.; Chiu, M.C.; Jaung, L.M.; Lu, Y.J.; Lin, H.J. Effects of farming systems on insect communities in the paddy fields of a simplified landscape during a pest-control intervention. Zool. Stud. 2021, 60, e56. [Google Scholar] [CrossRef]
- Kirse, A.; Bourlat, S.J.; Langen, K.; Zapke, B.; Zizka, V.M.A. Comparison of destructive and nondestructive DNA extraction methods for the metabarcoding of arthropod bulk samples. Mol. Ecol. Resour. 2023, 23, 92–105. [Google Scholar] [CrossRef]
- Leray, M.; Yang, J.Y.; Meyer, C.P.; Mills, S.C.; Agudelo, N.; Ranwez, V.; Boehm, J.T.; Machida, R.J. A new versatile primer set targeting a short fragment of the mitochondrial COI region for metabarcoding metazoan diversity: Application for characterizing coral reef fish gut contents. Front. Zool. 2013, 10, 34. [Google Scholar] [CrossRef]
- Schloss, P.D.; Gevers, D.; Westcott, S.L. Reducing the effects of pcr amplification and sequencing artifacts on 16s rrna-based studies. PLoS ONE 2011, 6, e27310. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Zhu, P.; Zheng, X.; Zhang, F.; Xu, H.; Yang, Y.; Chen, G.; Lu, Z.; Johnson, A.C.; Gurr, G.M. Quantifying the respective and additive effects of nectar plant crop borders and withholding insecticides on biological control of pests in subtropical rice. J. Pest Sci. 2018, 91, 575–584. [Google Scholar] [CrossRef]
- Linard, B.; Arribas, P.; Andújar, C.; Crampton-Platt, A.; Vogler, A.P. Lessons from genome skimming of arthropod-preserving ethanol. Mol. Ecol. Resour. 2016, 16, 1365–1377. [Google Scholar] [CrossRef]
- Marquina, D.; Esparza-Salas, R.; Roslin, T.; Ronquist, F. Establishing arthropod community composition using metabarcoding: Surprising inconsistencies between soil samples and preservative ethanol and homogenate from Malaise trap catches. Mol. Ecol. Resour. 2019, 19, 1516–1530. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.M.; Shi, Z.Y.; Zhang, S.Q.; Zhang, P.; Wilson, J.J.; Shih, C.; Li, J.; Li, X.D.; Yu, G.Y.; Zhang, A.B. Plant–herbivorous insect networks: Who is eating what revealed by long barcodes using high-throughput sequencing and Trinity assembly. Insect Sci. 2020, 28, 127–143. [Google Scholar] [CrossRef] [PubMed]
- Berthier, K.; Chapuis, M.P.; Moosavi, S.M.; Tohidi-Esfahani, D.; Sword, G.A. Nuclear insertions and heteroplasmy of mitochondrial DNA as two sources of intra-individual genomic variation in grasshoppers. Syst. Entomol. 2011, 36, 285–299. [Google Scholar] [CrossRef]
- Magnacca, K.N.; Brown, M.J.F. Tissue segregation of mitochondrial haplotypes in heteroplasmic Hawaiian bees: Implications for DNA barcoding. Mol. Ecol. Resour. 2010, 10, 60–68. [Google Scholar] [CrossRef]
- Boykin, L.M.; Armstrong, K.; Kubatko, L.; De Barro, P. DNA barcoding invasive insects: Database roadblocks. Invertebr. Syst. 2012, 26, 506–514. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The barcode of life data system (www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Landaverde-González, P.; Moo-Valle, H.; Murray, T.E.; Paxton, R.J.; Quezada-Euán, J.J.G.; Husemann, M. Sympatric lineage divergence in cryptic Neotropical sweat bees (Hymenoptera: Halictidae). Org. Divers. Evol. 2017, 17, 251–265. [Google Scholar] [CrossRef]
- Parslow, B.A.; Schwarz, M.P.; Stevens, M.I. Molecular diversity and species delimitation in the family Gasteruptiidae (Hymenoptera: Evanioidea). Genome 2021, 64, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Mioduchowska, M.; Czyz, M.J.; Goldyn, B.; Kur, J.; Sell, J. Instances of erroneous DNA barcoding of metazoan invertebrates: Are universal cox1 gene primers too “universal”? PLoS ONE 2018, 13, e0199609. [Google Scholar] [CrossRef] [PubMed]
- Porter, T.M.; Hajibabaei, M. Scaling up: A guide to high-throughput genomic approaches for biodiversity analysis. Mol. Ecol. 2018, 27, 313–338. [Google Scholar] [CrossRef] [PubMed]
- Zenker, M.M.; Specht, A.; Fonseca, V.G. Assessing insect biodiversity with automatic light traps in Brazil: Pearls and pitfalls of metabarcoding samples in preservative ethanol. Ecol. Evol. 2020, 10, 2352–2366. [Google Scholar] [CrossRef] [PubMed]
- Pawluczyk, M.; Weiss, J.; Links, M.G.; Aranguren, M.E.; Wilkinson, M.D.; Egea-Cortines, M. Quantitative evaluation of bias in PCR amplification and next-generation sequencing derived from metabarcoding samples. Anal. Bioanal. Chem. 2015, 407, 1841–1848. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.J.; Soubrier, J.; Weyrich, L.S.; Cooper, A. Environmental metabarcodes for insects: PCR reveals potential for taxonomic bias. Mol. Ecol. Resour. 2014, 14, 1160–1170. [Google Scholar] [CrossRef] [PubMed]
- Lowe, A.; Jones, L.; Brennan, G.; Creer, S.; de Vere, N. Seasonal progression and differences in major floral resource use by bees and hoverflies in a diverse horticultural and agricultural landscape revealed by DNA metabarcoding. J. Appl. Ecol. 2022, 59, 1484–1495. [Google Scholar] [CrossRef]
- Zhou, X.; Li, Y.Y.; Liu, S.L.; Yang, Q.; Su, X.; Zhou, L.L.; Tang, M.; Fu, R.B.; Li, J.G.; Huang, Q.F. Ultra-deep sequencing enables high-fidelity recovery of biodiversity for bulk arthropod samples without PCR amplification. Gigascience 2013, 2, 4. [Google Scholar] [CrossRef]
- Toju, H.; Baba, Y.G. DNA metabarcoding of spiders, insects, and springtails for exploring potential linkage between above- and below-ground food webs. Zool. Lett. 2018, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.N.; Bukhman, Y.V.; Jusino, M.A.; Scully, E.D.; Spiesman, B.J.; Gratton, C. Using high-throughput amplicon sequencing to determine diet of generalist lady beetles in agricultural landscapes. Biol. Control 2022, 170, 104920. [Google Scholar] [CrossRef]
- Wilson, T.; Looney, C.; Tembrock, L.R.; Dickerson, S.; Orr, J.; Gilligan, T.M.; Wildung, M. Insights into the prey of Vespa mandarinia (Hymenoptera: Vespidae) in Washington state, obtained from metabarcoding of larval feces. Front. Insect Sci. 2023, 3, 1134781. [Google Scholar] [CrossRef] [PubMed]
- Berman, T.S.; Inbar, M. Revealing cryptic interactions between large mammalian herbivores and plant-dwelling arthropods via DNA metabarcoding. Ecology 2022, 103, e03548. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Marco, F.; Urbaneja, A.; Jaques, J.A.; Rugman-Jones, P.F.; Stouthamer, R.; Tena, A. Untangling the aphid-parasitoid food web in citrus: Can hyperparasitoids disrupt biological control? Biol. Control 2015, 81, 111–121. [Google Scholar] [CrossRef]
- Zhu, Y.L.; Yang, F.; Yao, Z.W.; Wu, Y.K.; Liu, B.; Yuan, H.B.; Lu, Y.H. A molecular detection approach for a cotton aphid-parasitoid complex in northern China. Sci. Rep. 2019, 9, 15836. [Google Scholar] [CrossRef]
- Athey, K.J.; Chapman, E.G.; Harwood, J.D. A tale of two fluids: Does storing specimens together in liquid preservative cause DNA cross-contamination in molecular gut-content studies? Entomol. Exp. Appl. 2017, 163, 338–343. [Google Scholar] [CrossRef]
- Varennes, Y.D.; Boyer, S.; Wratten, S.D. Un-nesting DNA Russian dolls—The potential for constructing food webs using residual DNA in empty aphid mummies. Mol. Ecol. 2014, 23, 3925–3933. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Wu, H.; He, W.; Lai, G.; Li, J.; Liu, S.; Zhou, Q. Diversity of Parasitoid Wasps and Comparison of Sampling Strategies in Rice Fields Using Metabarcoding. Insects 2024, 15, 228. https://doi.org/10.3390/insects15040228
Wang L, Wu H, He W, Lai G, Li J, Liu S, Zhou Q. Diversity of Parasitoid Wasps and Comparison of Sampling Strategies in Rice Fields Using Metabarcoding. Insects. 2024; 15(4):228. https://doi.org/10.3390/insects15040228
Chicago/Turabian StyleWang, Liyang, Hongxuan Wu, Wei He, Guihong Lai, Junxi Li, Siling Liu, and Qiang Zhou. 2024. "Diversity of Parasitoid Wasps and Comparison of Sampling Strategies in Rice Fields Using Metabarcoding" Insects 15, no. 4: 228. https://doi.org/10.3390/insects15040228