
Antennal Sensitivity of Spotted Lanternflies, Lycorma delicatula: Differential Electrophysiological Responses of Males and Females to Compounds Derived from Host Plants and Conspecifics

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Chemicals
2.3. Electroantennogram (EAG) Preparation and Recording
2.4. Antennal Sensitivity Recording
2.4.1. Fixed-Dose Relative Response
2.4.2. Dose–Response
2.5. Statistical Analysis
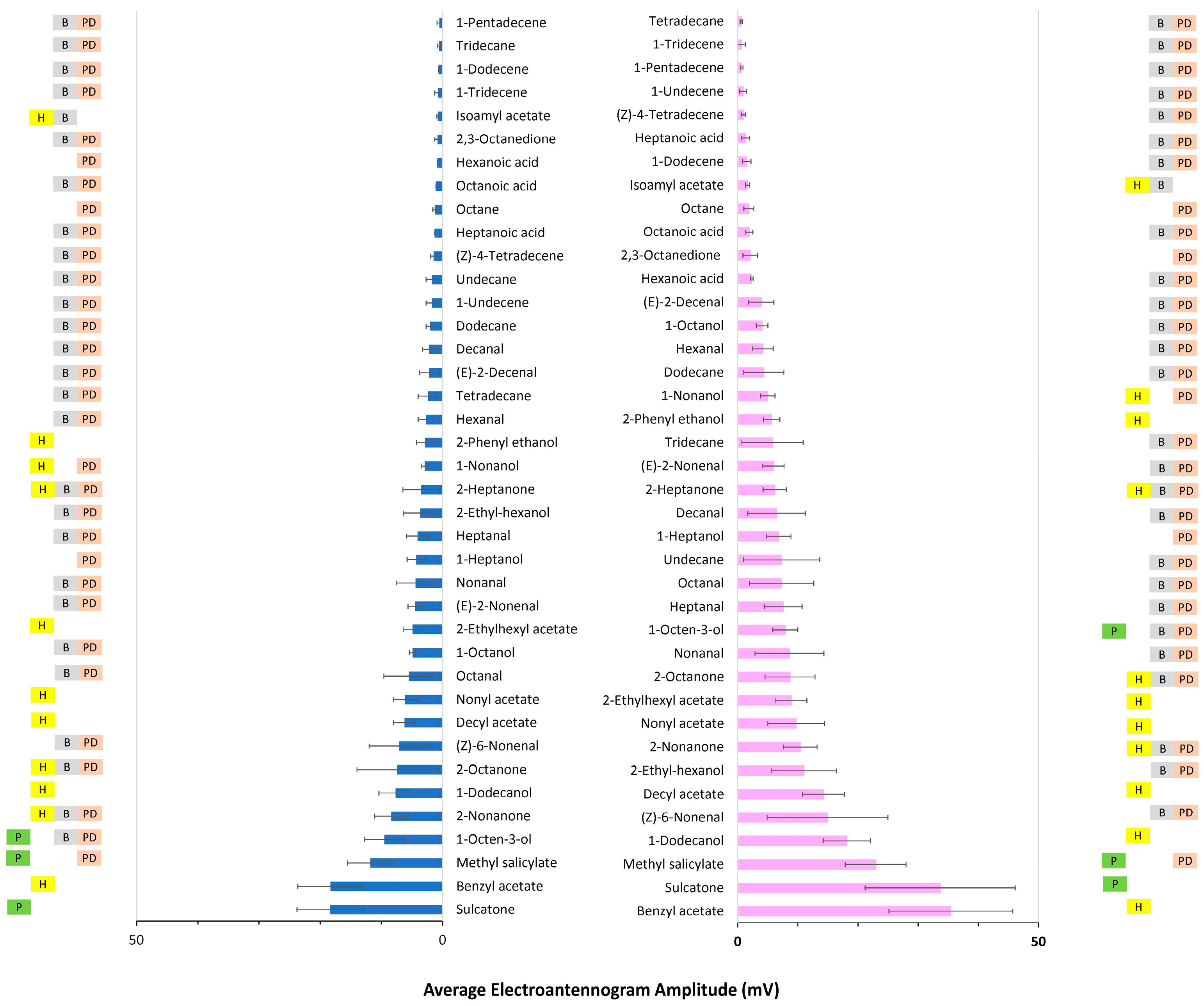
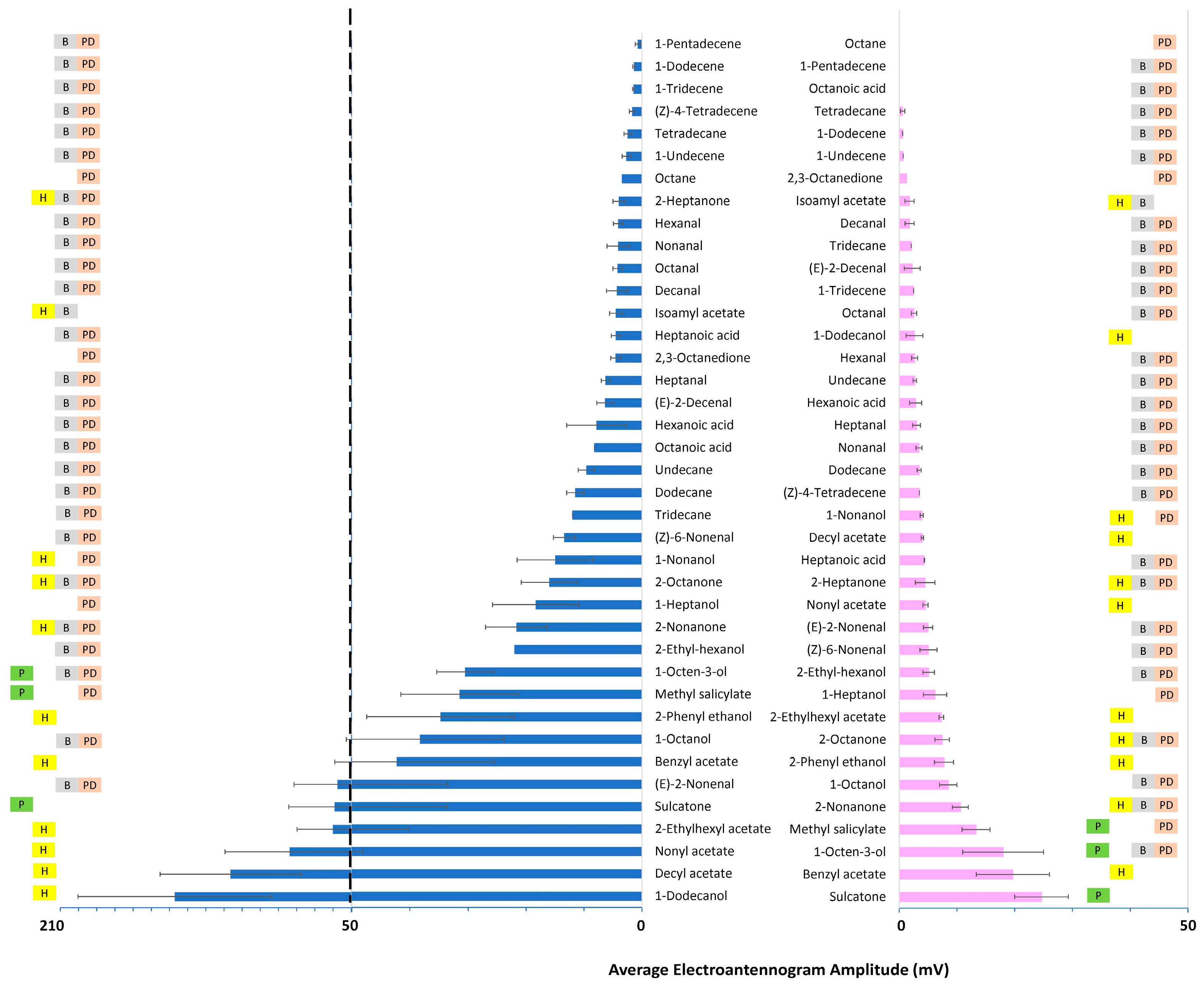
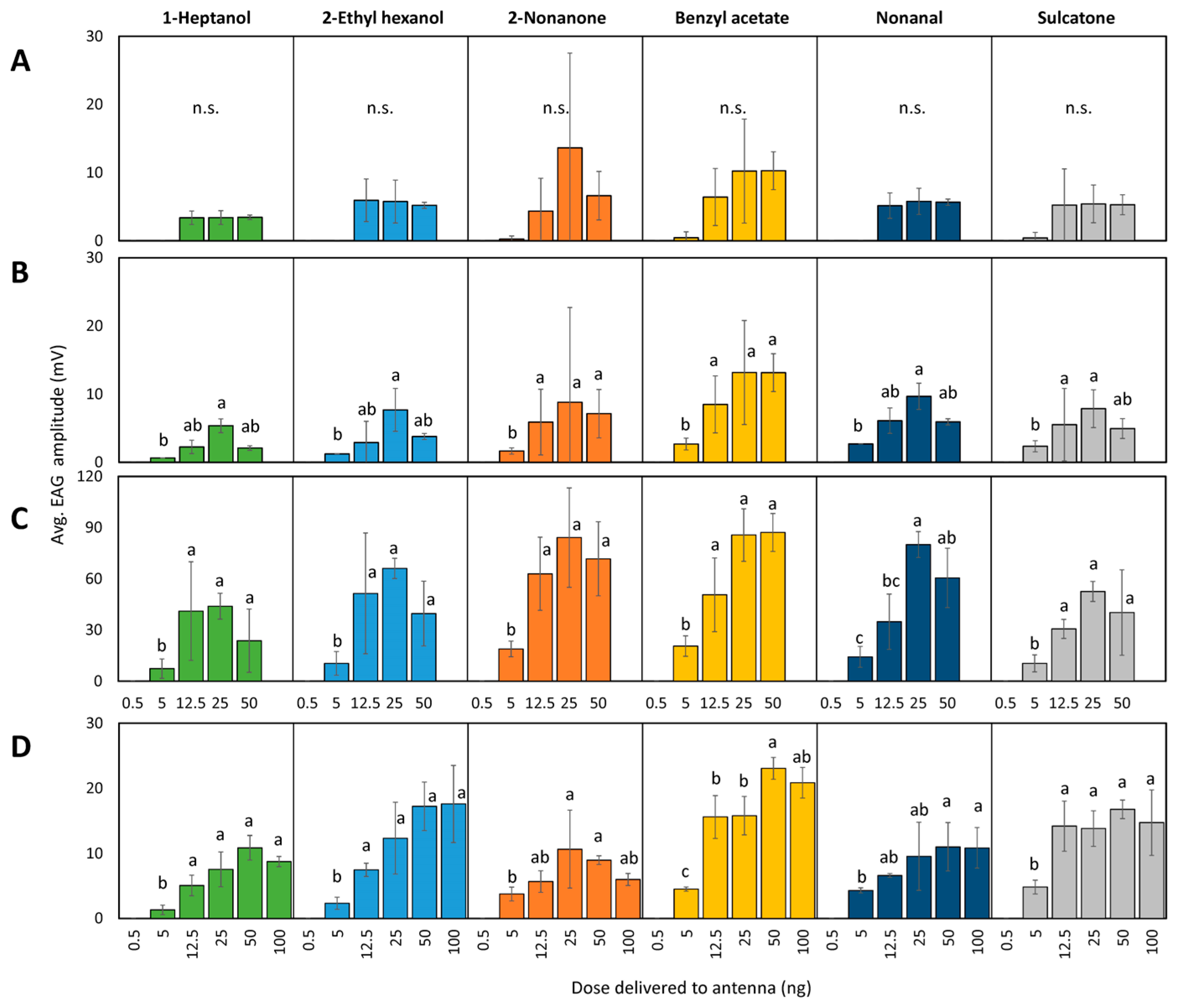
3. Results
3.1. Antennal Sensitivity
3.1.1. Fixed-Dose Relative Response
3.1.2. Dose–Response
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barringer, L.E.; Donovall, L.R.; Spichiger, S.-E.; Lynch, D.; Henry, D. The first new world record of Lycorma delicatula (Insecta: Hemiptera: Fulgoridae). Entomol. News 2015, 125, 20–23. [Google Scholar] [CrossRef]
- Barringer, L.; Ciafré, C.M. Worldwide feeding host plants of spotted lanternfly, with significant additions from North America. Environ. Entomol. 2020, 49, 999–1011. [Google Scholar] [CrossRef]
- Nixon, L.J.; Jones, S.K.; Tang, L.; Urban, J.; Felton, K.; Leskey, T.C. Survivorship and development of the invasive Lycorma delicatula (Hemiptera: Fulgoridae) on wild and cultivated temperate host plants. Environ. Entomol. 2022, 51, 222–228. [Google Scholar] [CrossRef]
- Urban, J.M. Perspective: Shedding light on spotted lanternfly impacts in the USA. Pest Manag. Sci. 2020, 76, 10–17. [Google Scholar] [CrossRef]
- Urban, J.M.; Leach, H. Biology and management of the spotted lanternfly, Lycorma delicatula (Hemiptera: Fulgoridae), in the united states. Annu. Rev. Entomol. 2023, 68, 151–167. [Google Scholar] [CrossRef]
- Nation Sr, J.L. Insect Physiology and Biochemistry, 4th ed.; CRC Press: Boca Raton, FL, USA, 2022; p. 555. [Google Scholar]
- Suh, E.; Bohbot, J.D.; Zwiebel, L.J. Peripheral olfactory signaling in insects. Curr. Opin. Insect Sci. 2014, 6, 86–92. [Google Scholar] [CrossRef]
- El-Shafie, H.A.F.; Faleiro, J.R. Semiochemicals and their potential use in pest management. In Biological Control of Pest and Vector Insects; Intech: London, UK, 2017; Volume 5772. [Google Scholar]
- Witzgall, P.; Kirsch, P.; Cork, A. Sex pheromones and their impact on pest management. J. Chem. Ecol. 2010, 36, 80–100. [Google Scholar] [CrossRef]
- Olsson, S.B.; Hansson, B.S. Electroantennogram and single sensillum recording in insect antennae. In Pheromone Signaling. Methods in Molecular Biology, vol 1068; Humana Press: Totowa, NJ, USA, 2013; pp. 157–177. [Google Scholar]
- Kaissling, K.-E. Pheromone reception in insects: The example of silk moths. In Neurobiology of Chemical Communication; Mucignat-Caretta, C., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 99–146. [Google Scholar]
- Yan, X.; Wang, Z.; Xie, J.; Deng, C.; Sun, X.; Hao, C. Glomerular organization of the antennal lobes of the diamondback moth, Plutella xylostella L. Front. Neuroanat. 2019, 13, 4. [Google Scholar] [CrossRef]
- Ngumbi, E.; Chen, L.; Fadamiro, H. Electroantennogram (EAG) responses of Microplitis croceipes and Cotesia marginiventris and their lepidopteran hosts to a wide array of odor stimuli: Correlation between EAG response and degree of host specificity? J. Insect Physiol. 2010, 56, 1260–1268. [Google Scholar] [CrossRef]
- Cooperband, M.F.; Wickham, J.D.; Warden, M.L. Factors guiding the orientation of nymphal spotted lanternfly, Lycorma delicatula. Insects 2023, 14, 279. [Google Scholar] [CrossRef]
- Faal, H.; Cooperband, M.F.; Canlas, I.; Carrillo, D. Evidence of pheromone use in a fulgorid, spotted lanternfly. Forests 2022, 13, 1639. [Google Scholar] [CrossRef]
- Faal, H.; Meier, L.R.; Canlas, I.J.; Murman, K.; Wallace, M.; Carrillo, D.; Cooperband, M.F. Volatiles from male honeydew excretions attract conspecific male spotted lanternflies, Lycorma delicatula (Hemiptera: Fulgoridae). Front. Insect Sci. 2022, 2, 982965. [Google Scholar] [CrossRef]
- Cooperband, M.F.; Murman, K. Responses of adult spotted lanternflies to artificial aggregations composed of all males or females. Front. Insect Sci. 2022, 2, 981832. [Google Scholar] [CrossRef]
- Derstine, N.T.; Meier, L.; Canlas, I.; Murman, K.; Cannon, S.; Carrillo, D.; Wallace, M.; Cooperband, M.F. Plant volatiles help mediate host plant selection and attraction of the spotted lanternfly (Hemiptera: Fulgoridae): A generalist with a preferred host. Environ. Entomol. 2020, 49, 1049–1062. [Google Scholar] [CrossRef]
- Cooperband, M.F.; Wickham, J.; Cleary, K.; Spichiger, S.-E.; Zhang, L.; Baker, J.; Canlas, I.; Derstine, N.; Carrillo, D. Discovery of three kairomones in relation to trap and lure development for spotted lanternfly (Hemiptera: Fulgoridae). J. Econ. Entomol. 2019, 112, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Faal, H.; Canlas, I.J.; Cossé, A.; Jones, T.H.; Carrillo, D.; Cooperband, M.F. Investigating photo-degradation as a potential pheromone production pathway in spotted lanternfly, Lycorma delicatula. Insects 2023, 14, 551. [Google Scholar] [CrossRef] [PubMed]
- Murman, K.; Setliff, G.P.; Pugh, C.V.; Toolan, M.J.; Canlas, I.; Cannon, S.; Abreu, L.; Fetchen, M.; Zhang, L.; Warden, M.L. Distribution, survival, and development of spotted lanternfly on host plants found in North America. Environ. Entomol. 2020, 49, 1270–1281. [Google Scholar] [CrossRef]
- Sant’ana, J.; Dickens, J.C. Comparative electrophysiological studies of olfaction in predaceous bugs, Podisus maculiventris and P. nigrispinus. J. Chem. Ecol. 1998, 24, 965–984. [Google Scholar] [CrossRef]
- Keena, M.A.; Hamilton, G.; Kreitman, D. The potential climatic range of spotted lanternfly may be broader than previously predicted. Front. Insect Sci. 2023, 3, 1092189. [Google Scholar] [CrossRef]
- Wang, R.-R.; Liu, J.-J.; Li, X.-Y.; Liang, A.-P.; Bourgoin, T. Relating antennal sensilla diversity and possible species behaviour in the planthopper pest Lycorma delicatula (Hemiptera: Fulgoromorpha: Fulgoridae). PLoS ONE 2018, 13, e0194995. [Google Scholar] [CrossRef]
- Beck, J.J.; Light, D.M.; Gee, W.S. Electrophysiological responses of male and female Amyelois transitella antennae to pistachio and almond host plant volatiles. Entomol. Exp. Appl. 2014, 153, 217–230. [Google Scholar] [CrossRef]
- Fraser, A.M.; Mechaber, W.L.; Hildebrand, J.G. Electroantennographic and behavioral responses of the sphinx moth Manduca sexta to host plant headspace volatiles. J. Chem. Ecol. 2003, 29, 1813–1833. [Google Scholar] [CrossRef]
- Pérez-Aparicio, A.; Torres-Vila, L.M.; Gemeno, C. EAG responses of adult Lobesia botrana males and females collected from Vitis vinifera and Daphne gnidium to larval host-plant volatiles and sex pheromone. Insects 2019, 10, 281. [Google Scholar] [CrossRef]
- Stelinski, L.L.; Il’Ichev, A.L.; Gut, L.J. Antennal and behavioral responses of virgin and mated oriental fruit moth (Lepidoptera: Tortricidae) females to their sex pheromone. Ann. Entomol. Soc. Am. 2006, 99, 898–904. [Google Scholar] [CrossRef]
- Raguso, R.A.; Light, D.M.; Pickersky, E. Electroantennogram responses of Hyles lineata (Sphingidae: Lepidoptera) to volatile compounds from Clarkia breweri (Onagraceae) and other moth-pollinated flowers. J. Chem. Ecol. 1996, 22, 1735–1766. [Google Scholar] [CrossRef]
- Spaethe, J.; Brockmann, A.; Halbig, C.; Tautz, J. Size determines antennal sensitivity and behavioral threshold to odors in bumblebee workers. Naturwissenschaften 2007, 94, 733–739. [Google Scholar] [CrossRef]
- Selzer, R. On the specificities of antennal olfactory receptor cells of Periplaneta americana. Chem. Senses 1984, 8, 375–395. [Google Scholar] [CrossRef]
- Vermeulen, A.; Rospars, J.-P. Why are insect olfactory receptor neurons grouped into sensilla? The teachings of a model investigating the effects of the electrical interaction between neurons on the transepithelial potential and the neuronal transmembrane potential. Eur. Biophys. J. 2004, 33, 633–643. [Google Scholar] [CrossRef]
- Pellegrino, M.; Nakagawa, T.; Vosshall, L.B. Single sensillum recordings in the insects Drosophila melanogaster and Anopheles gambiae. JoVE (J. Vis. Exp.) 2010, 36, e1725. [Google Scholar]
- Hao, Y.; Dietrich, C.H.; Dai, W. Structure and sensilla of the mouthparts of the spotted lanternfly Lycorma delicatula (Hemiptera: Fulgoromorpha: Fulgoridae), a polyphagous invasive planthopper. PLoS ONE 2016, 11, e0156640. [Google Scholar] [CrossRef]
- Syed, Z. Chemical ecology and olfaction in arthropod vectors of diseases. Curr. Opin. Insect Sci. 2015, 10, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Van Giessen, W.A.; Fescemyer, H.W.; Burrows, P.M.; Peterson, J.K.; Barnett, O.W. Quantification of electroantennogram responses of the primary rhinaria of Acyrthosiphon pisum (Harris) to C4-C8 primary alcohols and aldehydes. J. Chem. Ecol. 1994, 20, 909–927. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.S.; Mankin, R.W.; Lemire, G.F. Quantitation of the insect electroantennogram: Measurement of sensillar contributions, elimination of background potentials, and relationship to olfactory sensation. J. Insect Physiol. 1984, 30, 757–763. [Google Scholar] [CrossRef]
- Jacob, V.E.J.M. Current source density analysis of electroantennogram recordings: A tool for mapping the olfactory response in an insect antenna. Front. Cell. Neurosci. 2018, 12, 287. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Sex | Compound | p-Value | F-Ratio | d.f. | N |
|---|---|---|---|---|---|---|
| Fourth instar | Male | 1-Heptanol | 0.956 | 0.05 | 2, 9 | 12 |
| 2-Ethyl-hexanol | 0.980 | 0.02 | 2, 9 | 12 | ||
| 2-Nonanone | 0.255 | 1.61 | 3, 9 | 13 | ||
| Benzyl acetate | 0.081 | 3.11 | 3, 9 | 13 | ||
| Nonanal | 0.786 | 0.25 | 2, 9 | 12 | ||
| Sulcatone | 0.403 | 1.09 | 3, 9 | 13 | ||
| Female | 1-Heptanol | 0.013 | 5.80 | 3, 11 | 15 | |
| 2-Ethyl-hexanol | 0.012 | 5.93 | 3, 11 | 15 | ||
| 2-Nonanone | <0.001 | 14.72 | 3, 12 | 16 | ||
| Benzyl acetate | <0.001 | 28.23 | 3, 12 | 16 | ||
| Nonanal | 0.039 | 3.85 | 3, 12 | 16 | ||
| Sulcatone | 0.002 | 8.96 | 3, 12 | 16 | ||
| Adult | Male | 1-Heptanol | 0.002 | 8.31 | 3, 13 | 17 |
| 2-Ethyl-hexanol | 0.001 | 12.34 | 3, 12 | 16 | ||
| 2-Nonanone | <0.001 | 21.75 | 3, 13 | 17 | ||
| Benzyl acetate | <0.001 | 19.19 | 3, 13 | 17 | ||
| Nonanal | <0.001 | 15.04 | 3, 13 | 17 | ||
| Sulcatone | <0.001 | 12.90 | 3, 13 | 17 | ||
| Female | 1-Heptanol | <0.001 | 20.42 | 4, 10 | 15 | |
| 2-Ethyl-hexanol | <0.001 | 17.87 | 4, 10 | 15 | ||
| 2-Nonanone | 0.026 | 4.43 | 4, 10 | 15 | ||
| Benzyl acetate | <0.001 | 67.05 | 4, 10 | 15 | ||
| Nonanal | 0.034 | 4.03 | 4, 10 | 15 | ||
| Sulcatone | 0.001 | 13.43 | 4, 10 | 15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faal, H.; Cooperband, M.F. Antennal Sensitivity of Spotted Lanternflies, Lycorma delicatula: Differential Electrophysiological Responses of Males and Females to Compounds Derived from Host Plants and Conspecifics. Insects 2024, 15, 162. https://doi.org/10.3390/insects15030162
Faal H, Cooperband MF. Antennal Sensitivity of Spotted Lanternflies, Lycorma delicatula: Differential Electrophysiological Responses of Males and Females to Compounds Derived from Host Plants and Conspecifics. Insects. 2024; 15(3):162. https://doi.org/10.3390/insects15030162
Chicago/Turabian StyleFaal, Hajar, and Miriam F. Cooperband. 2024. "Antennal Sensitivity of Spotted Lanternflies, Lycorma delicatula: Differential Electrophysiological Responses of Males and Females to Compounds Derived from Host Plants and Conspecifics" Insects 15, no. 3: 162. https://doi.org/10.3390/insects15030162