
Optimal dsRNA Concentration for RNA Interference in Asian Citrus Psyllid
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Asian Citrus Psyllid Colony Establishment and Maintenance
2.2. Design and Cloning of Double-Stranded RNA Targets and RNA Synthesis
2.2.1. Selection of dsRNA Target Region and Off-Target Analysis
2.2.2. Molecular Cloning of Double-Stranded RNA Targets in Psyllid and dsRNA Synthesis
2.2.3. Ingestion-Access Period on Double-Stranded RNA
2.2.4. RNA Isolation, cDNA Synthesis, and Real-Time, Quantitative Reverse Transcriptase PCR Amplification
3. Results
3.1. Knockdown of Gene Expression for ACP Adults and 3rd Instar Nymphs, Post-dsRNA Ingestion-Access Period
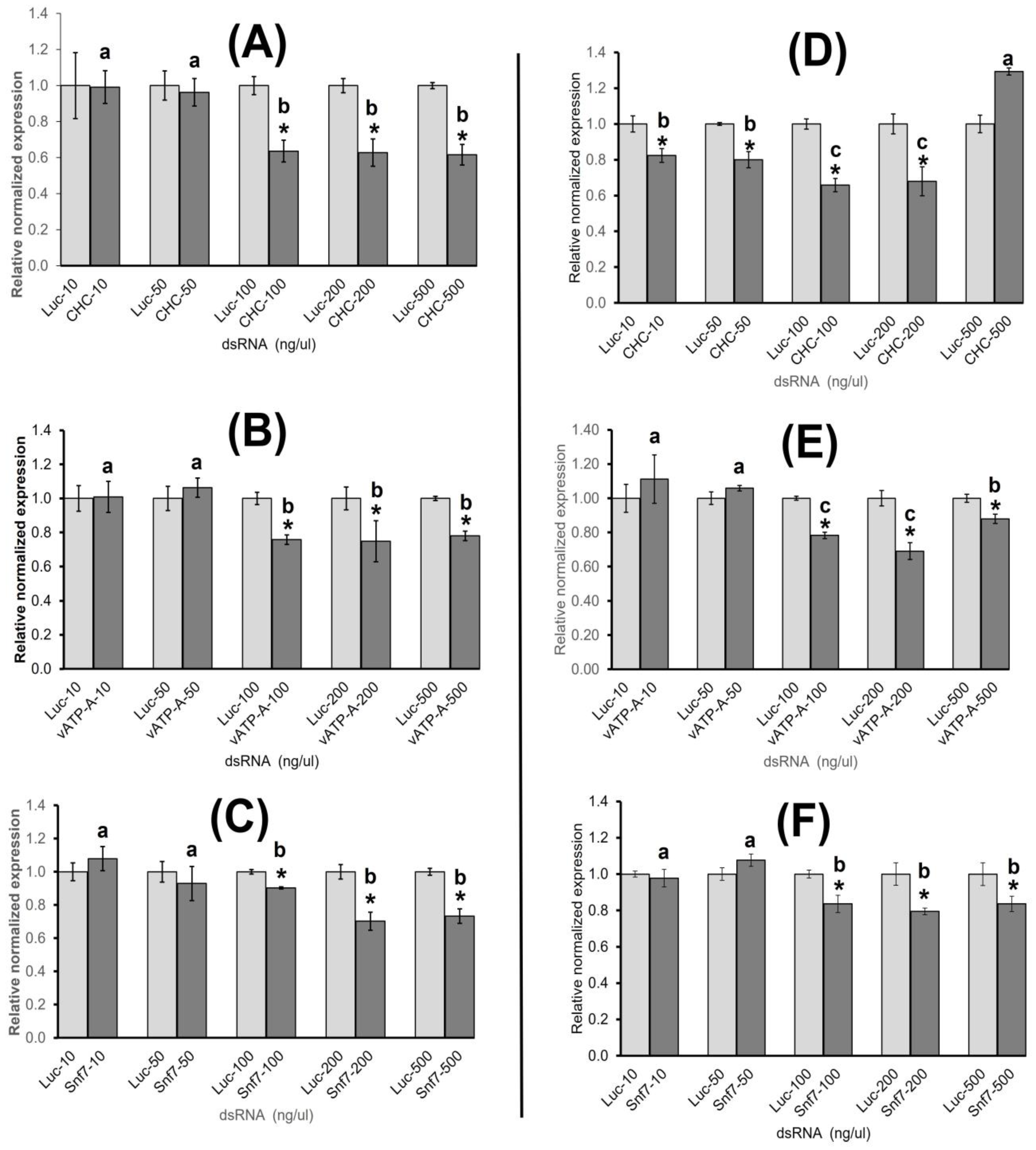
3.1.1. Relative Normalized Gene Expression of Target Genes in ACP Adults
3.1.2. Relative Normalized Gene Expression of Target Genes in ACP Nymphs
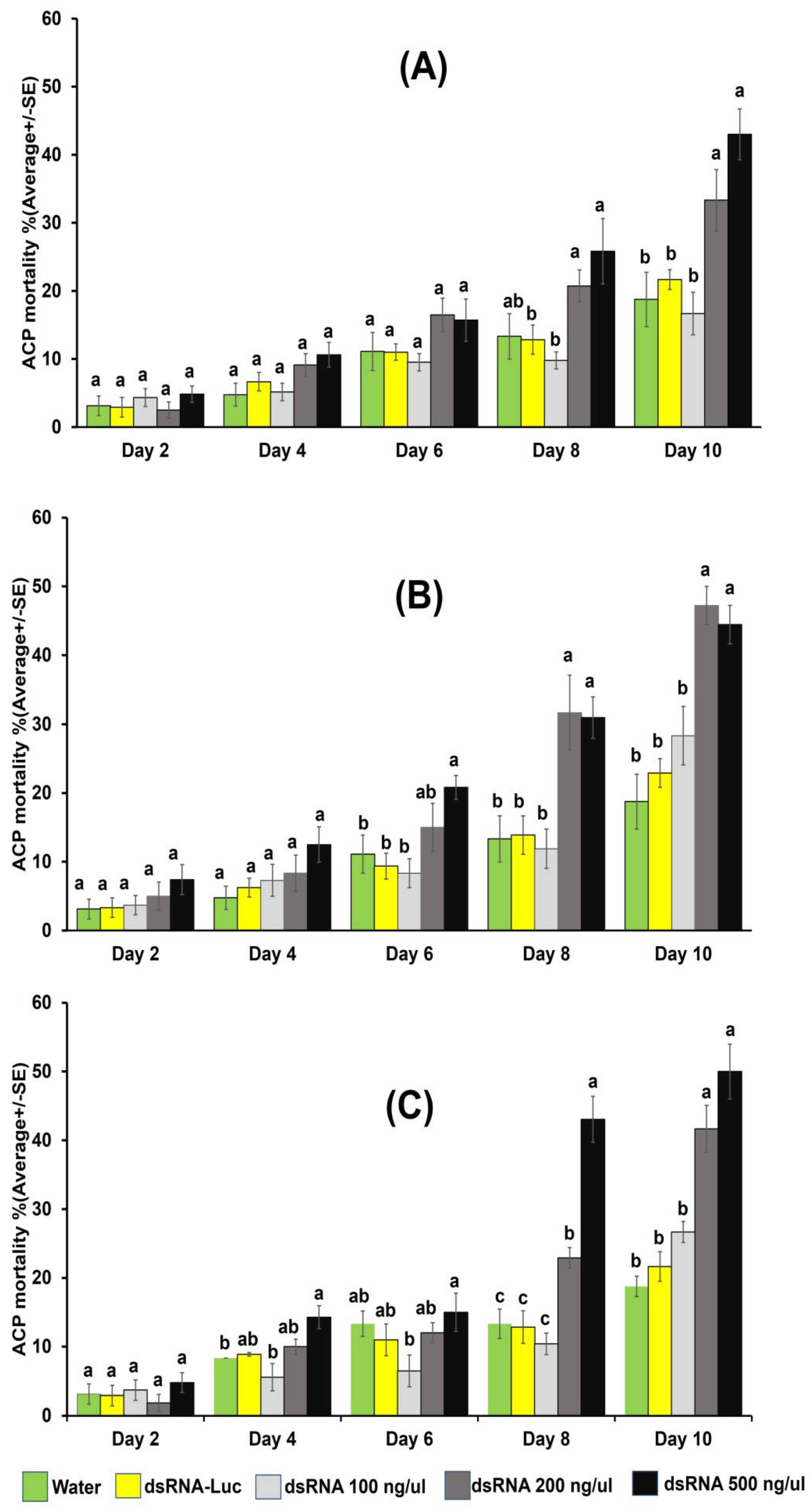
3.2. Mortality of Psyllid Adults through 10-Day Continuous Ingestion-Access Period on dsRNA Concentrations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hall, D.G.; Richardson, M.L.; Ammar, E.D.; Halbert, S.E. Asian citrus psyllid, Diaphorina citri, vector of citrus Huanglongbing disease. Entomol. Exp. Appl. 2013, 146, 207–223. [Google Scholar] [CrossRef]
- Wang, N.; Pierson, E.A.; Setubal, J.C.; Xu, J.; Levy, J.G.; Zhang, Y.; Li, J.; Rangel, L.T.; Martins, J. The Candidatus Liberibacter–host interface: Insights into pathogenesis mechanisms and disease control. Annu. Rev. Phytopathol. 2017, 55, 451–482. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, J.A.; Kostyk, B.C.; Stansly, P.A. Insecticidal suppression of Asian citrus psyllid Diaphorina citri (Hemiptera: Liviidae) Vector of Huanglongbing Pathogens. PLoS ONE 2014, 9, e112331. [Google Scholar] [CrossRef] [PubMed]
- Pardo, S.; Martínez, A.M.; Figueroa, J.I.; Chavarrieta, J.M.; Viñuela, E.; Rebollar-Alviter, Á.; Miranda, M.A.; Valle, J.; Pineda, S. Insecticide resistance of adults and nymphs of Asian citrus psyllid populations from Apatzingán valley, Mexico. Pest Manag. Sci. 2018, 74, 135–140. [Google Scholar] [CrossRef]
- Tiwari, S.; Mann, R.S.; Rogers, M.E.; Stelinski, L.L. Insecticide resistance in field populations of Asian citrus psyllid in Florida. Pest Manag. Sci. 2011, 67, 1258–1268. [Google Scholar] [CrossRef] [PubMed]
- San Miguel, K.; Scott, J.G. The next generation of insecticides: DsRNA is stable as a foliar-applied insecticide. Pest Manag. Sci. 2016, 72, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Taning, C.N.T.; Andrade, E.C.; Hunter, W.B.; Christiaens, O.; Smagghe, G. Asian citrus psyllid RNAi pathway-RNAi evidence. Sci. Rep. 2016, 6, 38082. [Google Scholar] [CrossRef] [PubMed]
- Christiaens, O.; Niu, J.; Taning, C.N.T. RNAi in Insects: A revolution in fundamental research and pest control Applications. Insects 2020, 11, 415. [Google Scholar] [CrossRef]
- Ruby, J.G.; Jan, C.H.; Bartel, D.P. Intronic microRNA precursors that bypass drosha processing. Nature 2007, 448, 83–86. [Google Scholar] [CrossRef]
- Tuschl, T.; Zamore, P.D.; Lehmann, R.; Bartel, D.P.; Sharp, P.A. Targeted mRNA degradation by double-stranded RNA in vitro. Genes Dev. 1999, 13, 3191–3197. [Google Scholar] [CrossRef]
- Andrade, E.C.; Hunter, W.B. RNAi Feeding Bioassay: Development of a non-transgenic approach to control Asian citrus psyllid and other hemipterans. Entomol. Exp. Appl. 2017, 162, 389–396. [Google Scholar] [CrossRef]
- Yu, X.; Killiny, N. RNA Interference-Mediated Control of Asian citrus psyllid, the vector of the huanglongbing bacterial pathogen. Trop. Plant Pathol. 2020, 45, 298–305. [Google Scholar] [CrossRef]
- Hajeri, S.; Killiny, N.; El-Mohtar, C.; Dawson, W.O.; Gowda, S. Citrus tristeza virus-based RNAi in citrus plants induces gene silencing in Diaphorina citri, a Phloem-Sap Sucking Insect Vector of Citrus Greening Disease (Huanglongbing). J. Biotechnol. 2014, 176, 42–49. [Google Scholar] [CrossRef]
- El-Shesheny, I.; Hajeri, S.; El-Hawary, I.; Gowda, S.; Killiny, N. Silencing abnormal wing disc gene of the Asian citrus psyllid, Diaphorina Citri disrupts adult wing development and increases nymph mortality. PLoS ONE 2013, 8, e65392. [Google Scholar] [CrossRef]
- Galdeano, D.M.; Breton, M.C.; Lopes, J.R.S.; Falk, B.W.; Machado, M.A. Oral delivery of double-stranded RNAs induces mortality in nymphs and adults of the Asian citrus psyllid, Diaphorina citri. PLoS ONE 2017, 12, e0171847. [Google Scholar] [CrossRef] [PubMed]
- Kishk, A.; Anber, H.A.I.; AbdEl-Raof, T.K.; El-Sherbeni, A.E.H.D.; Hamed, S.; Gowda, S.; Killiny, N. RNA interference of carboxyesterases causes nymph mortality in the Asian citrus psyllid, Diaphorina citri. Arch. Insect Biochem. Physiol. 2017, 94, e21377. [Google Scholar] [CrossRef] [PubMed]
- Santos-Ortega, Y.; Killiny, N. Silencing of sucrose hydrolase causes nymph mortality and disturbs adult osmotic homeostasis in Diaphorina citri (Hemiptera: Liviidae). Insect Biochem. Mol. Biol. 2018, 101, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Montero, J.R.; Arif, U.; Brown, J.K. Knockdown of ecdysteroid synthesis genes results in impaired molting and high mortality in Bactericera Cockerelli (Hemiptera: Triozidae). Pest Manag. Sci. 2022, 78, 2204–2214. [Google Scholar] [CrossRef]
- Wuriyanghan, H.; Rosa, C.; Falk, B.W. Oral Delivery of double-stranded RNAs and SiRNAs induces RNAi effects in the potato/tomato psyllid, Bactericerca cockerelli. PLoS ONE 2011, 6, e27736. [Google Scholar] [CrossRef]
- Hunter, W.B.; Wintermantel, W.M. Optimizing efficient rnai-mediated control of Hemipteran pests (Psyllids, Leafhoppers, Whitefly): Modified pyrimidines in dsRNA triggers. Plants 2021, 10, 1782. [Google Scholar] [CrossRef]
- Mamta, B.; Rajam, M.V. RNAi technology: A new platform for crop pest control. Physiol. Mol. Biol. Plants 2017, 23, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Joga, M.R.; Zotti, M.J.; Smagghe, G.; Christiaens, O. RNAi efficiency, systemic properties, and novel delivery methods for pest insect control: What we know so far. Front. Physiol. 2016, 7, 553. [Google Scholar] [CrossRef]
- Mondal, M.; Carver, M.; Brown, J.K. Characteristics of environmental RNAi in potato psyllid, Bactericera cockerelli (Sulc) (Hemiptera: Psylloidea: Triozidae). Front. Physiol. 2022, 13, 931951. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Killiny, N. Effect of silencing a boule homologue on the survival and reproduction of Asian citrus psyllid Diaphorina citri. Physiol. Entomol. 2018, 43, 268–275. [Google Scholar] [CrossRef]
- Killiny, N.; Kishk, A. Delivery of dsRNA through topical feeding for RNA interference in the citrus sap piercing-sucking Hemipteran, Diaphorina citri. Arch. Insect Biochem. Physiol. 2017, 95, e21394. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.J.; Huang, Y.L.; Yu, H.Z.; Li, N.Y.; Xie, Y.X.; Zhang, Q.; Zeng, X.D.; Hu, H.; Huang, A.J.; Yi, L.; et al. Silencing of the chitin synthase gene is lethal to the Asian citrus psyllid, Diaphorina citri. Int. J. Mol. Sci. 2019, 20, 3734. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.-j.; Zhou, C.-h.; Yu, H.-z.; Huang, Y.-l.; Liu, Y.-x.; Xie, Y.-x.; Wang, J.; Hu, W.; Huang, A.-j.; Su, H.-n.; et al. Potential roles of insect tropomyosin1-x1 isoform in the process of Candidatus Liberibacter asiaticus infection of Diaphorina citri. J. Insect Physiol. 2019, 114, 125–135. [Google Scholar] [CrossRef]
- Yu, H.Z.; Li, N.Y.; Li, B.; Toufeeq, S.; Xie, Y.X.; Huang, Y.L.; Du, Y.M.; Zeng, X.D.; Zhu, B.; Lu, Z.J. Immune functional analysis of chitin deacetylase 3 from the Asian citrus psyllid Diaphorina citri. Int. J. Mol. Sci. 2020, 21, 64. [Google Scholar] [CrossRef]
- Killiny, N.; Hajeri, S.; Tiwari, S.; Gowda, S.; Stelinski, L.L. Double-stranded RNA uptake through topical application, mediates silencing of five CYP4 genes and suppresses insecticide resistance in Diaphorina citri. PLoS ONE 2014, 9, e110536. [Google Scholar] [CrossRef]
- Tian, F.; Wang, Z.; Li, C.; Liu, J.; Zeng, X. UDP-glycosyltransferases are involved in imidacloprid resistance in the Asian citrus psyllid, Diaphorina citri (Hemiptera: Lividae). Pestic. Biochem. Physiol. 2019, 154, 23–31. [Google Scholar] [CrossRef]
- Kunte, N.; McGraw, E.; Bell, S.; Held, D.; Avila, L.A. Prospects, challenges and current status of RNAi through insect feeding. Pest Manag. Sci. 2020, 76, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Vijaya Sudhakara Rao, K.; Renuka, P.; Sheshu Madhav, M.; Mangrauthia, S.K. Key enzymes and proteins of crop insects as candidate for RNAi based gene silencing. Front. Physiol. 2015, 6, 119. [Google Scholar] [CrossRef]
- Guo, C.F.; Qiu, J.H.; Hu, Y.W.; Xu, P.P.; Deng, Y.Q.; Tian, L.; Wei, Y.Y.; Sang, W.; Liu, Y.T.; Qiu, B.L. Silencing of V-ATPase-E gene causes midgut apoptosis of Diaphorina citri and affects its acquisition of huanglongbing pathogen. Insect Sci. 2023, 30, 1022–1034. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Gowda, S.; Killiny, N. Double-stranded rna delivery through soaking mediates silencing of the muscle protein 20 and increases mortality to the Asian citrus psyllid, Diaphorina citri. Pest Manag. Sci. 2017, 73, 1846–1853. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Hoy, M.A. Clathrin heavy chain is important for viability, oviposition, embryogenesis and, possibly, systemic rnai response in the predatory mite Metaseiulus occidentalis. PLoS ONE 2014, 9, e110874. [Google Scholar] [CrossRef] [PubMed]
- Dow, J.A.T.; Davies, S.A.; Guo, Y.; Graham, S.; Finbow, M.E.; Kaiser, K. Molecular Genetic analysis of V-ATPase function in Drosophila melanogaster. J. Exp. Biol. 1997, 200, 237–245. [Google Scholar] [CrossRef]
- Grace, R.; Massimino, C.; Shippy, T.; Tank, W.; Hosmani, P.S.; Flores-Gonzalez, M.; Mueller, L.A.; Hunter, W.B.; Benoit, J.B.; Brown, S.J.; et al. Genomic identification, annotation, and comparative analysis of Vacuolar-type ATP synthase subunits in Diaphorina citri. GigaByte 2022, 2022, gigabyte39. [Google Scholar] [CrossRef]
- Wollert, T.; Wunder, C.; Lippincott-Schwartz, J.; Hurley, J.H. Membrane scission by the ESCRT-III complex. Nature 2009, 458, 172–177. [Google Scholar] [CrossRef]
- Thakre, N.; Carver, M.; Paredes-Montero, J.R.; Mondal, M.; Saberi, E.; Ponvert, N.; Qureshi, J.A.; Brown, J.K. UV-LASER adjuvant facilitated delivery of 100–600 base pair dsRNAs to the tomato plant vasculature and evidence of biological activity by gene knockdown in the potato psyllid. Pest Manag. Sci. 2023. online ahead of print. [Google Scholar] [CrossRef]
- Fisher, T.; Vyas, M.; He, R.; Nelson, W.; Cicero, J.; Willer, M.; Kim, R.; Kramer, R.; May, G.; Crow, J.; et al. Comparison of potato and Asian citrus psyllid adult and nymph transcriptomes identified vector transcripts with potential involvement in circulative, propagative Liberibacter transmission. Pathogens 2014, 3, 875–907. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef]
- Güssow, D.; Clackson, T. Direct clone characterization from plaques and colonies by the polymerase chain reaction. Nucleic Acids Res. 1989, 17, 4000. [Google Scholar] [CrossRef] [PubMed]
- Russell, C.W.; Pelz-Stelinski, K.S. Development of an artificial diet and feeding system for juvenile stages of the Asian citrus psyllid, Diaphorina citri. Entomol. Exp. Appl. 2015, 154, 171–176. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ibanez, F.; Tamborindeguy, C. Selection of reference genes for expression analysis in the potato psyllid, Bactericera cockerelli. Insect Mol. Biol. 2016, 25, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Marques, V.V.; Angelotti-Mendonça, J.; Roberto, S.R. Advances and Challenges in RNA Interference Technology for Citrus Huanglongbing Vector Control. Horticulturae 2021, 7, 277. [Google Scholar] [CrossRef]
- Jain, R.G.; Robinson, K.E.; Asgari, S.; Mitter, N. Current scenario of RNAi-based hemipteran control. Pest Manag. Sci. 2021, 77, 2188–2196. [Google Scholar] [CrossRef]
- Hunter, W.B.; Glick, E.; Paldi, N.; Bextine, B.R. Advances in RNA interference: DsRNA treatment in trees and grapevines for insect pest suppression. Southwest. Entomol. 2012, 37, 85–87. [Google Scholar] [CrossRef]
- Pacheco, I.d.S.; Galdeano, D.M.; Maluta, N.K.P.; Lopes, J.R.S.; Machado, M.A. Gene silencing of Diaphorina citri candidate effectors promotes changes in feeding behaviors. Sci. Rep. 2020, 10, 5992. [Google Scholar] [CrossRef]
- Terenius, O.; Papanicolaou, A.; Garbutt, J.S.; Eleftherianos, I.; Huvenne, H.; Kanginakudru, S.; Albrechtsen, M.; An, C.; Aymeric, J.L.; Barthel, A.; et al. RNA interference in lepidoptera: An overview of successful and unsuccessful studies and implications for experimental design. J. Insect Physiol. 2011, 57, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Walker, W.B.; Allen, M.L. RNA Interference-Mediated knockdown of IAP in Lygus lineolaris induces mortality in adult and pre-adult life stages. Entomol. Exp. Appl. 2011, 138, 83–92. [Google Scholar] [CrossRef]
- Araujo, R.N.; Santos, A.; Pinto, F.S.; Gontijo, N.F.; Lehane, M.J.; Pereira, M.H. RNA interference of the salivary gland nitrophorin 2 in the triatomine bug Rhodnius prolixus (Hemiptera: Reduviidae) by dsRNA ingestion or injection. Insect Biochem. Mol. Biol. 2006, 36, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Peng, H.; Yao, Q.; Chen, H.; Xie, Q.; Tang, B.; Zhang, W. Developmental control of a lepidopteran pest Spodoptera exigua by ingestion of bacteria expressing dsRNA of a non-midgut gene. PLoS ONE 2009, 4, e6225. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.G.; Michel, K.; Bartholomay, L.C.; Siegfried, B.D.; Hunter, W.B.; Smagghe, G.; Zhu, K.Y.; Douglas, A.E. Towards the elements of successful insect RNAi. J. Insect Physiol. 2013, 59, 1212–1221. [Google Scholar] [CrossRef] [PubMed]
- Angelotti-Mendonça, J.; Bassan, M.M.; Marques, J.P.R.; Yamamoto, P.T.; Figueira, A.; Piedade, S.M.D.S.; Mourão Filho, F.A.A. Knockdown of calreticulin, laccase, and Snf7 genes through RNAi Is not effective to control the Asian citrus psyllid (Hemiptera: Livideae). J. Econ. Entomol. 2020, 113, 2931–2940. [Google Scholar] [CrossRef]
- Mao, J.; Zeng, F. Plant-mediated RNAi of a gap gene-enhanced tobacco tolerance against the Myzus persicae. Transgenic Res. 2014, 23, 145–152. [Google Scholar] [CrossRef]
- Vaschetto, L. RNAi strategies for pest management: Methods and protocols. In Methods in Molecular Biology; Springer: New York, NY, USA, 2022. [Google Scholar] [CrossRef]
- Pitino, M.; Coleman, A.D.; Maffei, M.E.; Ridout, C.J.; Hogenhout, S.A. Silencing of aphid genes by dsRNA feeding from plants. PLoS ONE 2011, 6, e25709. [Google Scholar] [CrossRef]
- Christiaens, O.; Swevers, L.; Smagghe, G. dsRNA degradation in the pea aphid (Acyrthosiphon pisum) associated with lack of response in RNAi feeding and injection assay. Peptides 2014, 53, 307–314. [Google Scholar] [CrossRef]
- Winston, W.M.; Molodowitch, C.; Hunter, C.P. Systemic RNAi in C. elegans requires the putative transmembrane protein SID-1. Science 2002, 295, 2456–2459. [Google Scholar] [CrossRef]
- Huvenne, H.; Smagghe, G. Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: A Review. J. Insect Physiol. 2010, 56, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Bolognesi, R.; Ramaseshadri, P.; Anderson, J.; Bachman, P.; Clinton, W.; Flannagan, R.; Ilagan, O.; Lawrence, C.; Levine, S.; Moar, W. Characterizing the mechanism of action of double-stranded RNA activity against western corn rootworm (Diabrotica virgifera virgifera LeConte). PLoS ONE 2012, 7, e47534. [Google Scholar] [CrossRef]
- Ye, C.; Hu, X.S.; Wang, Z.W.; Wei, D.; Smagghe, G.; Christiaens, O.; Niu, J.; Wang, J.J. Involvement of clathrin-dependent endocytosis in cellular dsRNA uptake in aphids. Insect Biochem. Mol. Biol. 2021, 132, 103557. [Google Scholar] [CrossRef] [PubMed]
- Maxson, M.E.; Grinstein, S. The vacuolar-type H+-ATPase at a glance—More than a proton pump. J. Cell Sci. 2014, 127, 4987–4993. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.L.; Goulding, D.; Field, M.C. Clathrin-mediated endocytosis is essential in Trypanosoma brucei. EMBO J. 2003, 22, 4991–5002. [Google Scholar] [CrossRef]
- Grant, B.; Hirsh, D. Receptor-mediated endocytosis in the Caenorhabditis elegans oocyte. Mol. Biol. Cell 1999, 10, 4311–4326. [Google Scholar] [CrossRef]
- Li, C.; Xia, Y. Vacuolar ATPase subunit H is essential for the survival and moulting of Locusta migratoria manilensis. Insect Mol. Biol. 2012, 21, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Miyata, K.; Ozawa, S.; Hasegawa, K. Systemic RNAi of V-ATPase subunit B causes molting defect and developmental abnormalities in Periplaneta fuliginosa. Insect Sci. 2019, 26, 721–731. [Google Scholar] [CrossRef]
- Yao, J.; Rotenberg, D.; Afsharifar, A.; Barandoc-Alviar, K.; Whitfield, A.E. Development of RNAi methods for peregrinus maidis, the corn planthopper. PLoS ONE 2013, 8, e70243. [Google Scholar] [CrossRef]
- Zeng, J.; Mu, L.L.; Jin, L.; Ali Anjum, A.; Li, G.Q. Evaluation of three vacuolar ATPase genes as potential RNAi target in Henosepilachna vigintioctopunctata. J. Asia. Pac. Entomol. 2021, 24, 55–63. [Google Scholar] [CrossRef]
- Cuccato, G.; Polynikis, A.; Siciliano, V.; Graziano, M.; di Bernardo, M.; di Bernardo, D. Modeling RNA interference in mammalian cells. BMC Syst. Biol. 2011, 5, 19. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| dsRNA Primers | Target Gene | Sequence (5′ to 3′) | Tm | |
|---|---|---|---|---|
| vATPase-A F | vATPase-A | TTATCTCAGAGCATTTACATCCCCAAGGGTG | 61.4 | |
| vATPase-A R | CTTGACAAGTGTATTCTCATGAACAAGACC | 58.1 | ||
| Snf7 F | Snf7 | GCTCACAAGCACATGGACGTGAACC | 62.5 | |
| Snf7 R | AACTTCAGTAGATCCTTGTCCAGTTCTTCC | 59.6 | ||
| CHC F | CHC | CAGTACGCGGACGTGGAAGGAGG | 63.0 | |
| CHC R | TCGAAATAGCCGCGGTCCTGG | 61.1 | ||
| qRT-PCR Primers and Probes | Target Gene | Sequence (5′ to 3′) | Tm | Efficiency |
| vATPase-A qF | vATPase-A | AGTGGTTATCCTGCCTACCT | 53.7 | 95% |
| vATPase-A qR | CGTGCTGGCAGAGTCAAATGCTTG | 61.1 | ||
| Probe | GGAGATACAGCACCCACAATAC | 54.7 | ||
| Snf7 qF | Snf7 | CAGCAGATTGATGGCACATTG | 54.5 | 92% |
| Snf7 qR | GGCATTCTTCATGGTGGTAAGA | 60.7 | ||
| Probe | AATTGAGATGCAGCGGGAAGCTCT | 54.5 | ||
| CHC qF | CHC | AGCGAGGAGTTCCGTTTG | 54.1 | 90% |
| CHC qR | GGTCCTGGTAGTAGTTGATGAG | 59.6 | ||
| Probe | CGTGGTGCACGCAGATGAACTA | 53.5 | ||
| RPL5 qF | RPL5 | TCCAAAGGCAAGATCCAGAAA | 53.7 | 93.5% |
| RPL5 qR | AGAAGCTCACTTTGGCTCAACGGA | 60.5 | ||
| Probe | GGAAGTTAGCTTTGGCAGTAGA | 54.2 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saberi, E.; Mondal, M.; Paredes-Montero, J.R.; Nawaz, K.; Brown, J.K.; Qureshi, J.A. Optimal dsRNA Concentration for RNA Interference in Asian Citrus Psyllid. Insects 2024, 15, 58. https://doi.org/10.3390/insects15010058
Saberi E, Mondal M, Paredes-Montero JR, Nawaz K, Brown JK, Qureshi JA. Optimal dsRNA Concentration for RNA Interference in Asian Citrus Psyllid. Insects. 2024; 15(1):58. https://doi.org/10.3390/insects15010058
Chicago/Turabian StyleSaberi, Esmaeil, Mosharrof Mondal, Jorge R. Paredes-Montero, Kiran Nawaz, Judith K. Brown, and Jawwad A. Qureshi. 2024. "Optimal dsRNA Concentration for RNA Interference in Asian Citrus Psyllid" Insects 15, no. 1: 58. https://doi.org/10.3390/insects15010058