

Novel Methodology for Localizing and Studying Insect Dorsal Rim Area Morphology in 2D and 3D



{kind=link}
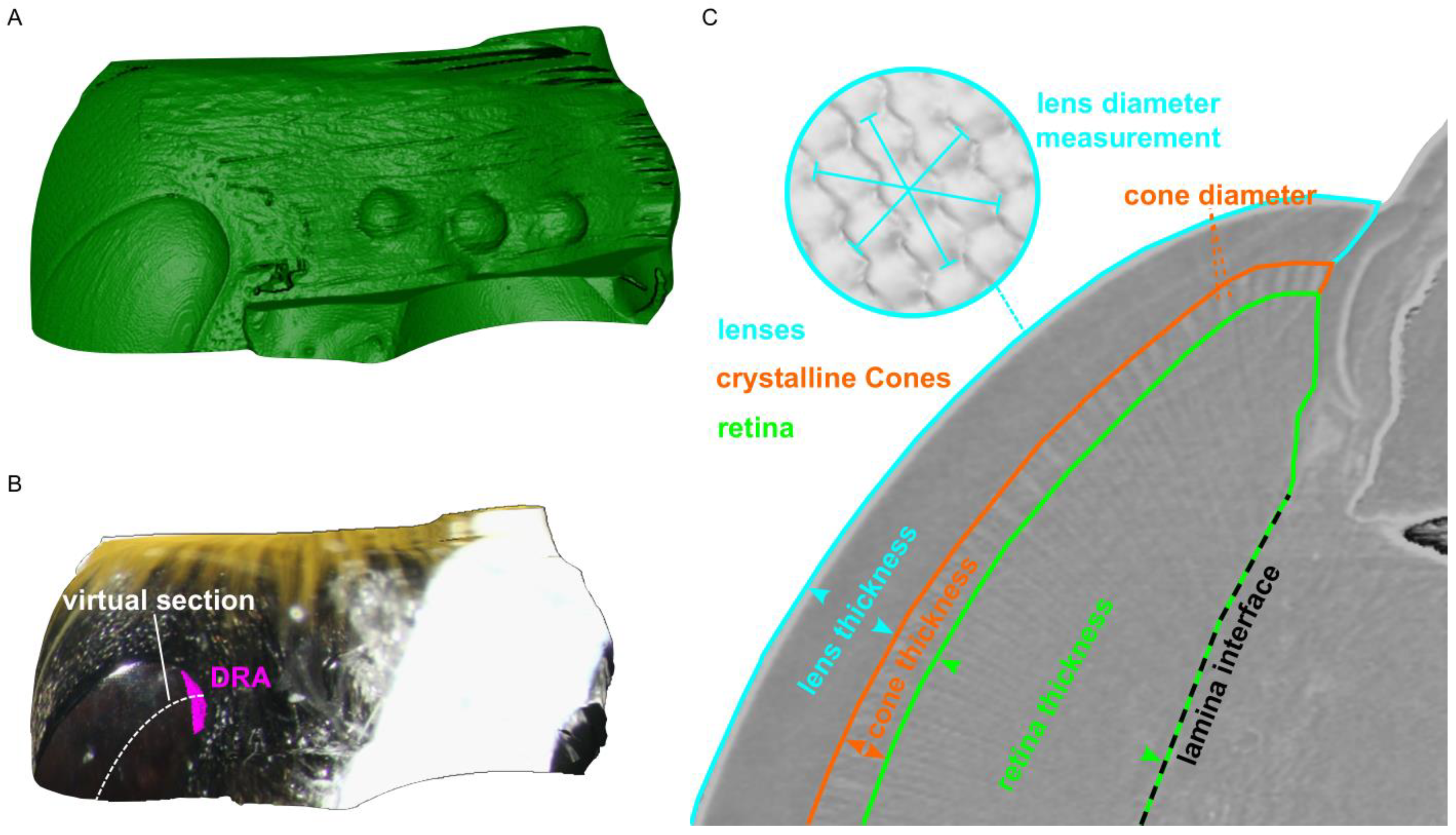{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Handling
2.2. Acquisition of 2D DRA Images and Surface Area Measurements
2.3. Staining and Embedding
2.4. Micro-CT
2.5. The 3D Compound Eye and Surface Area Measuremets
2.6. Localizing the DRA on a 3D Head Model
2.7. DRA Allometry and Overall Compound Eye Surface Area
2.8. DRA Ommatidial Structure Measurements
3. Results and Discussion
3.1. DRA Localization on a Homogeneous Eye Surface
3.2. A Comprehensive Study of the Internal and External Morphology of DRA Structures
3.3. Method Implementation on Bombus terrestris to Study How the DRA Varies with Body Size
3.4. Limitations of the Method
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wiltschko, R.; Wiltschko, W. The process of learning sun compass orientation in young homing pigeons. Naturwissenschaften 1980, 67, 512–514. [Google Scholar] [CrossRef]
- Wehner, R. The hymenopteran skylight compass: Matched filtering and parallel coding. J. Exp. Biol. 1989, 146, 63–85. [Google Scholar] [CrossRef]
- Dyer, F.C.; Dickinson, J.A. Development of sun compensation by honeybees: How partially experienced bees estimate the sun’s course. Proc. Natl. Acad. Sci. USA 1994, 91, 4471–4474. [Google Scholar] [CrossRef]
- Dacke, M.; Byrne, M.J.; Scholtz, C.H.; Warrant, E.J. Lunar orientation in a beetle. Proc. R. Soc. B Biol. Sci. 2004, 271, 361–365. [Google Scholar] [CrossRef]
- Dacke, M.; Nilsson, D.E.; Scholtz, C.H.; Byrne, M.; Warrant, E.J. Insect orientation to polarized moonlight. Nature 2003, 424, 33. [Google Scholar] [CrossRef]
- Nørgaard, T.; Nilsson, D.E.; Henschel, J.R.; Garm, A.; Wehner, R. Vision in the nocturnal wandering spider Leucorchestris arenicola (Araneae: Sparassidae). J. Exp. Biol. 2008, 211, 816–823. [Google Scholar] [CrossRef] [Green Version]
- Wehner, R. Arthropods. In Animal Homing; Papi, F., Ed.; Springer: Berlin/Heidelberg, Germany, 1992; pp. 45–144. [Google Scholar]
- Papi, F. Animal navigation at the end of the century: A retrospect and a look forward. Ital. J. Zool. 2001, 68, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Wehner, R. The ant’s celestial compass system: Spectral and polarization channelss. In Orientation and Communication in Arhtropods; Lehrer, M., Ed.; Springer: Berlin/Heidelberg, Germany, 1997; pp. 145–185. [Google Scholar]
- Cartwright, B.A.; Collett, T.S. Landmark learning in bees—Experiments and models. J. Comp. Physiol. A 1983, 151, 521–543. [Google Scholar] [CrossRef]
- Wolf, H.; Wehner, R. Pinpointing food sources: Olfactory and anemotactic orientation in desert ants, Cataglyphis fortis. J. Exp. Biol. 2000, 203, 857–868. [Google Scholar] [CrossRef]
- Wehner, R. Desert ant navigation: How miniature brains solve complex tasks. J. Comp. Physiol. A Neuroethol. Sensory, Neural, Behav. Physiol. 2003, 189, 579–588. [Google Scholar] [CrossRef] [Green Version]
- Wehner, R.; Michel, B.; Antonsen, P. Visual navigation in insects: Coupling of egocentric and geocentric information. J. Exp. Biol. 1996, 199, 129–140. [Google Scholar] [CrossRef]
- Wehner, R.; Boyer, M.; Loertscher, F.; Sommer, S.; Menzi, U. Ant navigation: One-way routes rather than maps. Curr. Biol. 2006, 16, 75–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knaden, M.; Wehner, R. Nest mark orientation in desert ants Cataglyphis: What does it do to the path integrator? Anim. Behav. 2005, 70, 1349–1354. [Google Scholar] [CrossRef]
- Cheng, K.; Narendra, A.; Sommer, S.; Wehner, R. Traveling in clutter: Navigation in the central Australian desert ant Melophorus bagoti. Behav. Process. 2009, 80, 261–268. [Google Scholar] [CrossRef]
- Bisch-Knaden, S.; Wehner, R. Local vectors in desert ants: Context-dependent landmark learning during outbound and homebound runs. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2003, 189, 181–187. [Google Scholar] [CrossRef]
- Bregy, P.; Sommer, S.; Wehner, R. Nest-mark orientation versus vector navigation in desert ants. J. Exp. Biol. 2008, 211, 1868–1873. [Google Scholar] [CrossRef] [Green Version]
- Seidl, T.; Wehner, R. Visual and tactile learning of ground structures in desert ants. J. Exp. Biol. 2006, 209, 3336–3344. [Google Scholar] [CrossRef] [Green Version]
- Steck, K.; Knaden, M.; Hansson, B.S. Do desert ants smell the scenery in stereo? Anim. Behav. 2010, 79, 939–945. [Google Scholar] [CrossRef]
- Wehner, R.; Labhart, T. Polarisation Vision. In Invertebrate Vision; Warrant, E., Nilsson, D.-E., Eds.; Cambridge University Press: Cambridge, UK, 2006; pp. 291–348. [Google Scholar]
- Frisch, K.V. The Dance Language and Orientation of Bees; Harvard University Press: Cambridge, MA, USA, 1993. [Google Scholar]
- Brines, M.L.; Gould, J.L. Skylight polarization patterns and animal orientation. J. Exp. Biol. 1982, 96, 69–91. [Google Scholar] [CrossRef]
- Stockhammer, K. Die orientierung nach der schwingungsrichtung linear polarisierten lichtes und ihre sinnesphysiologischen grundlagen. In Ergebnisse der Biologie; Springer: Berlin/Heidelberg, Germany, 1959; pp. 23–56. [Google Scholar]
- Israelachvili, J.; Wilson, M. Absorption characteristics of oriented photopigments in microvilli. Biol. Cybern. 1976, 21, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, T.H.; Wehner, R. Restrictions on rotational and translational diffusion of pigment in the membranes of a rhabdomeric photoreceptor. J. Gen. Physiol. 1977, 70, 453–490. [Google Scholar] [CrossRef]
- Hardie, R.C. Properties of photoreceptors R7 and R8 in dorsal marginal ommatidia in the compound eyes of Musca and Calliphora. J. Comp. Physiol. A 1984, 154, 157–165. [Google Scholar] [CrossRef]
- Labhart, T.; Meyer, E.P. Detectors for polarized skylight in insects: A survey of ommatidial specializations in the dorsal rim area of the compound eye. Microsc. Res. Tech. 1999, 47, 368–379. [Google Scholar] [CrossRef]
- Gokan, N. The compound eye of the dung beetle Geotrupes auratus (Coleoptera: Scarabaeidae). Appl. Entomol. Zool. 1989, 24, 133–146. [Google Scholar] [CrossRef] [Green Version]
- Gokan, N. The compound eye of the dung beetle, Onthophagus posticus (Coleoptera: Scarabaeidae). N. Zeal. Entomol. 1990, 13, 7–15. [Google Scholar] [CrossRef]
- Dacke, M.; Nordström, P.; Scholtz, C.H.; Warrant, E.J. A specialized dorsal rim area for polarized light detection in the compound eye of the Scarab Beetle Pachysoma striatum. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2002, 188, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Dacke, M.; Nordström, P.; Scholtz, C.H. Twilight orientation to polarised light in the crepuscular dung beetle Scarabaeus zambesianus. J. Exp. Biol. 2003, 206, 1535–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dacke, M.; Nordström, P.; Scholtz, C. Two sets of compound eyes in dungbeetles Pachysoma: A specialization for skylight navigation. In Proceedings of the 27th Göttingen Neurobiology Conference, Stuttgart, Germany, 1 June 1999. [Google Scholar]
- Labhart, T.; Meyer, E.P.; Schenker, L. Specialized ommatidia for polarization vision in the compound eye of cockchafers, Melolontha melolontha (Coleoptera, Scarabaeidae). Cell Tissue Res. 1992, 268, 419–429. [Google Scholar] [CrossRef]
- Hardie, R.; Vogt, K.; Rudolph, A. The compound eyes of the tsetse fly (Glossina morsitans morsitans and Glossina palpalis palpalis). J. Insect Physiol. 1989, 35, 423–431. [Google Scholar] [CrossRef]
- Strausfeld, N.J.; Wunderer, H. Optic lobe projections of marginal ommatidia in Calliphora erythrocephala specialized for detecting polarized light. Cell Tissue Res. 1985, 242, 163–178. [Google Scholar] [CrossRef]
- Wada, S. Spezielle randzonale ommatidien der fliegen (Diptera: Brachycera): Architektur und verteilung in den komplexauaen. Z. Für Morphol. Der Tiere 1974, 77, 87–125. [Google Scholar] [CrossRef]
- Wada, S. Spezielle randzonale ommatidien von Calliphora erythrocephala meig (Diptera: Calliphoridae): Architektur der zentralen rhabdomeren-kolumne und topographie im komplexauge. Int. J. Insect Morphol. Embryol. 1974, 3, 397–424. [Google Scholar] [CrossRef]
- Wunderer, H.; Smola, U. Fine structure of ommatidia at the dorsal eye margin of Calliphora erythrocephala meigen (Diptera: Calliphoridae): An eye region specialised for the detection of polarized light. Int. J. Insect Morphol. Embryol. 1982, 11, 25–38. [Google Scholar] [CrossRef]
- Aepli, F.; Labhart, T.; Meyer, E.P. Structural specializations of the cornea and retina at the dorsal rim of the compound eye in Hymenopteran insects. Cell Tissue Res. 1985, 239, 19–24. [Google Scholar] [CrossRef]
- Zeil, J.; Ribi, W.; Nerandra, A. Polarisation vision in ants, bees and wasps. In Polarized Light and Polarization Vision in Animal Sciences; Horvath, G., Marshall, J., Collin, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 41–60. [Google Scholar]
- Reid, S.F. Life in the Dark: Vision and Navigation in a Nocturnal Bull Ant. Ph.D. Thesis, The Australian National University, Canberra, Australia, January 2010. [Google Scholar]
- von Helversen, O.; Edrich, W. The spectral sensitivity of polarized light orientation in the honey bee. J. Comp. Physiol. 1974, 94, 33–47. [Google Scholar] [CrossRef]
- Räber, F.W. Retinatopographie und Sehfeldtopologie des Komplexauges von Cataglyphis bicolor (Formicidae, Hymenoptera) und Einiger Verwandter Formiciden-Arten. Ph.D. Thesis, University of Zürich, Zürich, Switzerland, 1979. [Google Scholar]
- Sommer, E.W. Untersuchungen zur Topographischen Anatomie der Retina und zur Sehfeldtopologie im Auge der Honigbiene, Apis mellifera (Hymenoptera). Ph.D. Thesis, University of Zürich, Zürich, Switzerland, 1979. [Google Scholar]
- Meyer, E.P.; Labhart, T. Pore canals in the cornea of a functionally specialized area of the honey bee’s compound eye. Cell Tissue Res. 1981, 216, 491–501. [Google Scholar] [CrossRef]
- Greiner, B.; Cronin, T.W.; Ribi, W.A.; Wcislo, W.T.; Warrant, E.J. Anatomical and physiological evidence for polarisation vision in the nocturnal bee Megalopta genalis. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2007, 193, 591–600. [Google Scholar] [CrossRef]
- Herrling, P.L. Regional distribution of three ultrastructural retinula types in the retina of Cataglyphis bicolor Fabr. (Formicidae, Hymenoptera). Cell Tissue Res. 1976, 169, 247–266. [Google Scholar] [CrossRef]
- Labhart, T. The Electrophysiology of photoreceptors in different eye regions of the desert ant, Cataglyphis bicolor. J. Comp. Physiol. A 1986, 158, 1–7. [Google Scholar] [CrossRef]
- Meyer, E.P.; Domanico, V. Microvillar orientation in the photoreceptors of the ant Cataglyphis bicolor. Cell Tissue Res. 1999, 295, 355–361. [Google Scholar] [CrossRef]
- Duelli, P.; Wehner, R. The spectral sensitivity of polarized light orientation in Cataglyphis bicolor (Formicidae, Hymenoptera). J. Comp. Physiol. 1973, 86, 37–53. [Google Scholar] [CrossRef]
- Narendra, A.; Alkaladi, A.; Raderschall, C.A.; Robson, S.K.A.; Ribi, W.A. Compound eye adaptations for diurnal and nocturnal lifestyle in the intertidal ant, Polyrhachis sokolova. PLoS ONE 2013, 8, e76015. [Google Scholar] [CrossRef]
- Narendra, A.; Ramirez-Esquivel, F.; Ribi, W.A. Compound eye and ocellar structure for walking and flying modes of locomotion in the Australian ant, Camponotus consobrinus. Sci. Rep. 2016, 6, 22331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labhart, T.; Baumann, F.; Bernard, G.D. Specialized ommatidia of the polarization-sensitive dorsal rim area in the eye of monarch butterflies have non-functional reflecting tapeta. Cell Tissue Res. 2009, 338, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Kolb, G. Ultrastructure and adaptation in the retina of Aglais urticae (Lepidoptera). Zoomorphology 1985, 105, 90–98. [Google Scholar] [CrossRef]
- Stalleicken, J.; Labhart, T.; Mouritsen, H. Physiological characterization of the compound eye in monarch butterflies with focus on the dorsal rim area. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2006, 192, 321–331. [Google Scholar] [CrossRef]
- Hämmerle, B.; Kolb, G. Retinal ultrastructure of the dorsal eye region of Pararge aegeria (Linne) (Lepidoptera: Satyridae). Int. J. Insect Morphol. Embryol. 1996, 25, 305–315. [Google Scholar] [CrossRef]
- Meinecke, C.C. The fine structure of the compound eye of the African armyworm moth, Spodoptera exempta Walk. (Lepidoptera, Noctuidae). Cell Tissue Res. 1981, 12, 333–347. [Google Scholar] [CrossRef]
- Meyer, E.P.; Labhart, T. Morphological specializations of dorsal rim ommatidia in the compound eye of dragonflies and damselfies (Odonata). Cell Tissue Res. 1993, 272, 17–22. [Google Scholar] [CrossRef]
- Egelhaaf, A.; Dambach, M. Giant rhabdomes in a specialized region of the compound eye of a cricket: Cycloptiloides canariensis (Insecta, Gryllidae). Zoomorphology 1983, 102, 65–77. [Google Scholar] [CrossRef]
- Labhart, T.; Keller, K. Fine structure and growth of the polarization- sensitive dorsal rim area in the compound eye of larval crickets. Naturwissenschaften 1992, 79, 527–529. [Google Scholar] [CrossRef]
- Homberg, U.; Paech, A. Ultrastructure and orientation of ommatidia in the dorsal rim area of the locust compound eye. Arthropod Struct. Dev. 2002, 30, 271–280. [Google Scholar] [CrossRef]
- Goulson, D. Social organization and conflict. In Bumblebees, Behaviour, Ecology, and Conservation; Oxford University Press: Oxford, UK, 2010; pp. 21–43. [Google Scholar]
- Cane, J.H. Estimation of bee size using intertegular span (Apoidea). J. Kansas Entomol. Soc. 1987, 60, 145–147. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.B.; Bernhardt, G.; Raine, N.E.; Abel, R.L.; Sykes, D.; Ahmed, F.; Pedroso, I.; Gill, R.J. Exploring miniature insect brains using micro-CT scanning techniques. Sci. Rep. 2016, 6, 21768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, G.J.; Ribi, W.; Bech, M.; Bodey, A.J.; Rau, C.; Steuwer, A.; Warrant, E.J.; Baird, E. The dual function of orchid bee ocelli as revealed by X-ray microtomography. Curr. Biol. 2016, 26, 1319–1324. [Google Scholar] [CrossRef]
- Romell, J.; Jie, V.W.; Miettinen, A.; Baird, E.; Hertz, H.M. Laboratory phase-contrast nanotomography of unstained Bombus terrestris compound eyes. J. Microsc. 2021, 283, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Paganin, D.; Mayo, S.C.; Gureyev, T.E.; Miller, P.R.; Wilkins, S.W. Simultaneous phase and amplitude extraction from a single defocused image of a homogeneous object. J. Microsc. 2002, 206, 33–40. [Google Scholar] [CrossRef]
- Marone, F.; Stampanoni, M. Regridding reconstruction algorithm for real-time tomographic imaging. J. Synchrotron Radiat. 2012, 19, 1029–1037. [Google Scholar] [CrossRef] [Green Version]
- Limaye, A. Drishti: A volume exploration and presentation tool. In Proceedings of the Developments in X-ray Tomography VIII; SPIE: Bellingham, WA, USA, 2012; Volume 8506, pp. 191–199. [Google Scholar] [CrossRef] [Green Version]
- Taylor, G.J.; Tichit, P.; Schmidt, M.D.; Bodey, A.J.; Rau, C.; Baird, E. Bumblebee visual allometry results in locally improved resolution and globally improved sensitivity. eLife 2019, 8, e40613. [Google Scholar] [CrossRef]
- Ijiri, T.; Todo, H.; Hirabayashi, A.; Kohiyama, K.; Dobashi, Y. Digitization of natural objects with micro CT and photographs. PLoS ONE 2018, 13, e0195852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaethe, J.; Chittka, L. Interindividual variation of eye optics and single object resolution in bumblebees. J. Exp. Biol. 2003, 206, 3447–3453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaethe, J.; Weidenmüller, A. Size variation and foraging rate in bumblebees (Bombus terrestris). Insectes Soc. 2002, 49, 142–146. [Google Scholar] [CrossRef] [Green Version]
- Handschuh, S.; Baeumler, N.; Schwaha, T.; Ruthensteiner, B. A correlative approach for combining microCT, light and transmission electron microscopy in a single 3D scenario. Front. Zool. 2013, 10, 44. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jie, V.W.; Miettinen, A.; Baird, E. Novel Methodology for Localizing and Studying Insect Dorsal Rim Area Morphology in 2D and 3D. Insects 2023, 14, 670. https://doi.org/10.3390/insects14080670
Jie VW, Miettinen A, Baird E. Novel Methodology for Localizing and Studying Insect Dorsal Rim Area Morphology in 2D and 3D. Insects. 2023; 14(8):670. https://doi.org/10.3390/insects14080670
Chicago/Turabian StyleJie, Vun Wen, Arttu Miettinen, and Emily Baird. 2023. "Novel Methodology for Localizing and Studying Insect Dorsal Rim Area Morphology in 2D and 3D" Insects 14, no. 8: 670. https://doi.org/10.3390/insects14080670