

Biochemical Defense Responses in Red Rice Genotypes Possessing Differential Resistance to Brown Planthopper, Nilaparvata lugens (Stål)
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Total Phenol
2.2. Assay of Defense Enzymes
2.2.1. Peroxidase (PO)
2.2.2. Polyphenol Oxidase (PPO)
2.2.3. Superoxide Dismutase (SOD)
2.2.4. Catalase (CAT)
2.3. Soluble Protein
2.4. Crude Silica
2.5. Statistical Analysis
3. Results
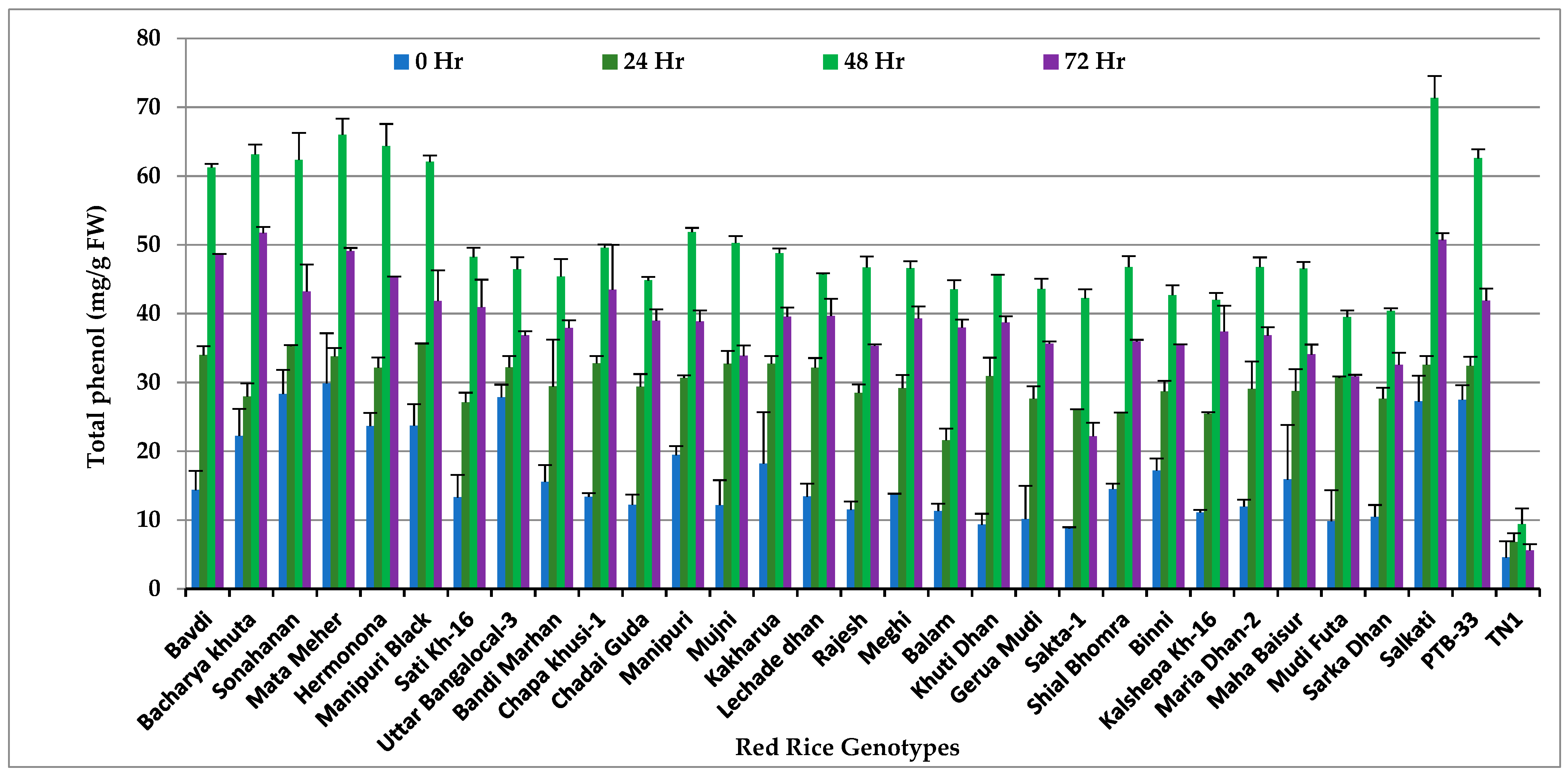
3.1. Total Phenols
3.2. Peroxidase (POD)
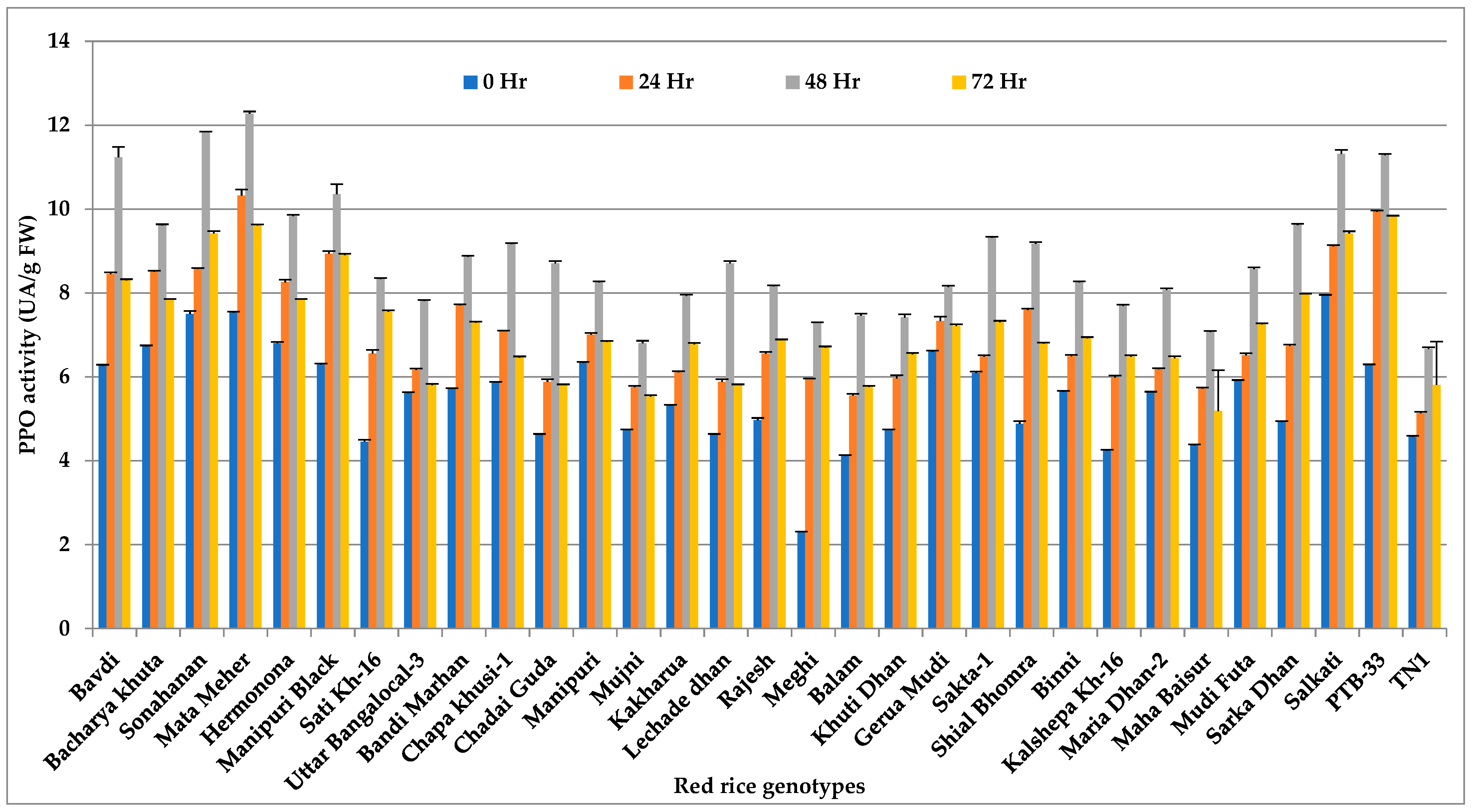
3.3. Polyphenol Oxidase (PPO)
3.4. Superoxide Dismutase
3.5. Catalase (CAT)
3.6. Soluble Proteins
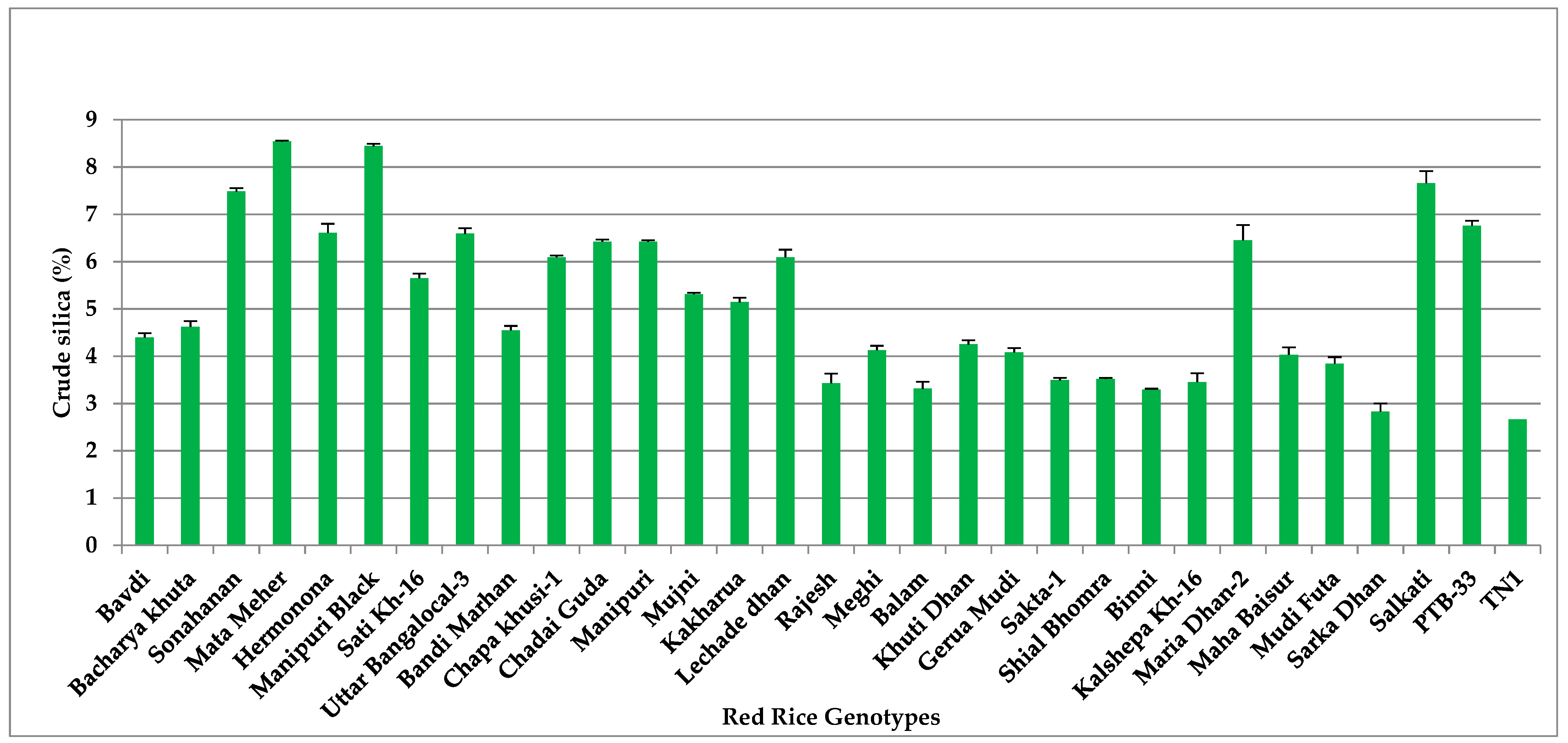
3.7. Crude Silica (Si)
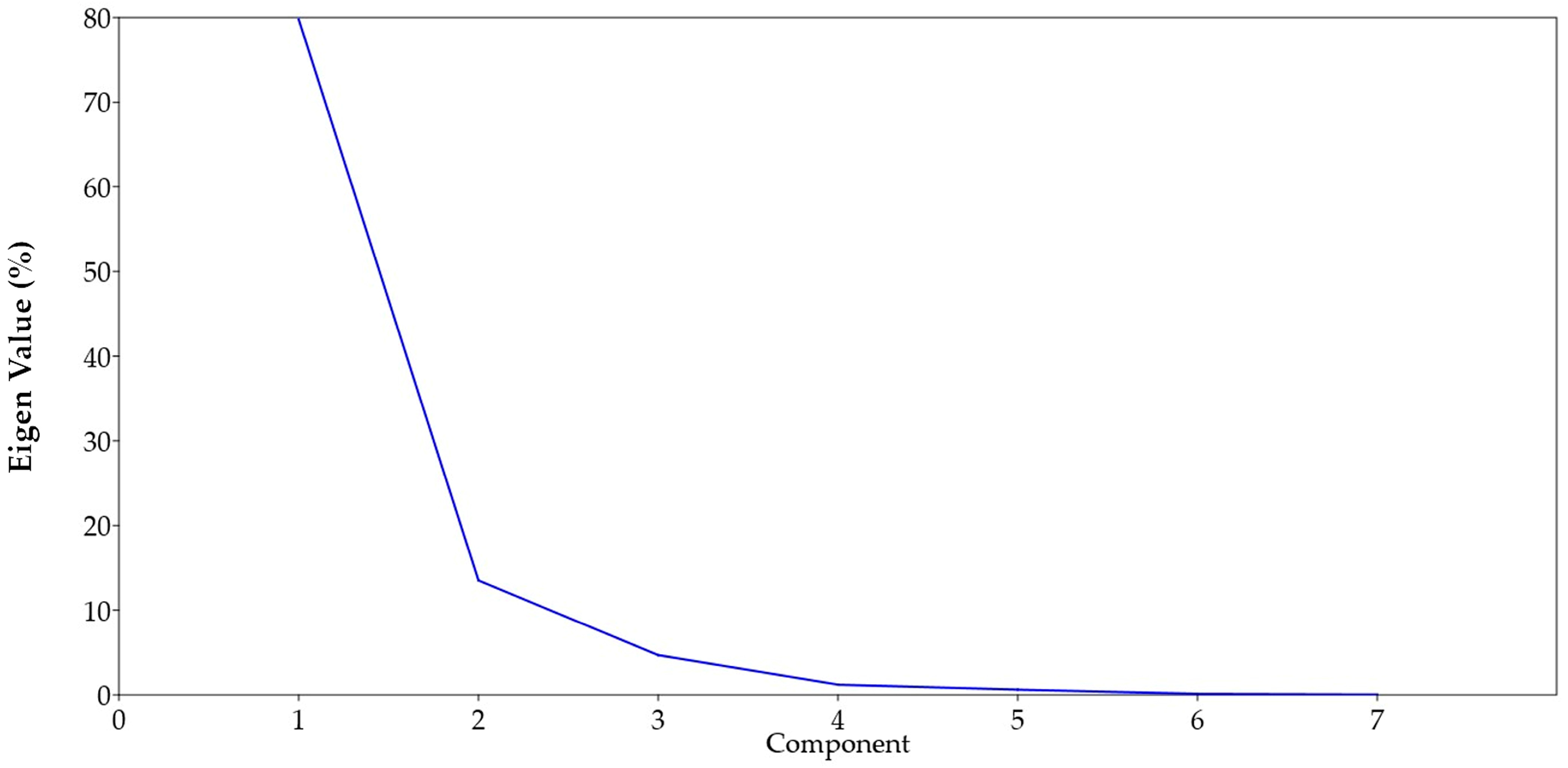
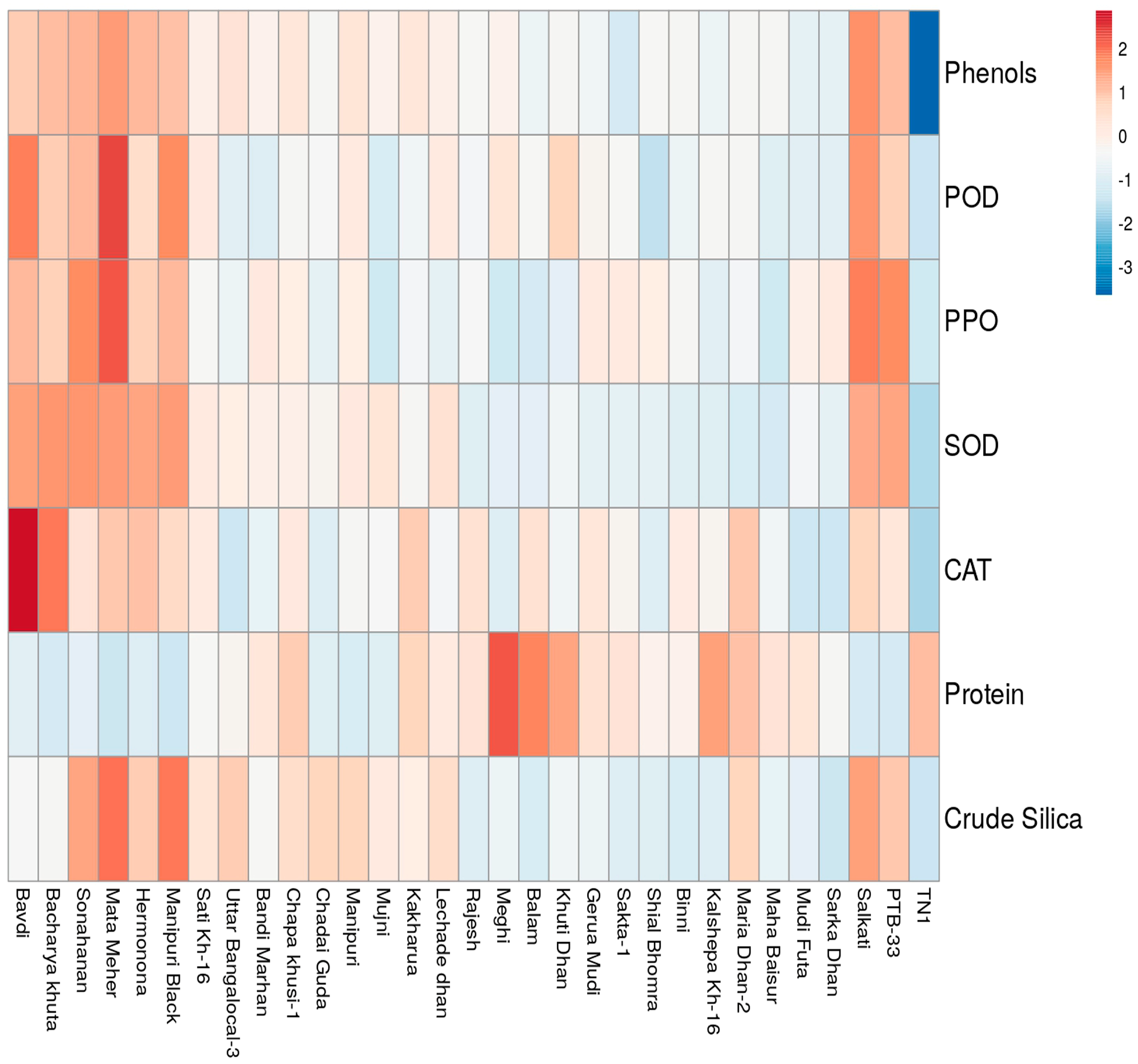
3.8. Principal Component (PCA) and Cluster Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khush, G.S. What it will take to feed 5.0 billion rice consumers in 2030. Plant Mol. Biol. 2005, 59, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Pathak, H.; Nayak, A.K.; Jena, M.; Singh, O.N.; Samal, P.; Sharma, S.G. Rice Research Book: Rice Research for Enhancing Productivity; ICAR-National Rice Research Institute: Cuttack, India, 2018; p. Px+542. [Google Scholar]
- Jena, M.; Panda, R.S.; Sahu, R.K.; Mukherjee, A.K.; Dhua, U. Evaluation of rice genotypes for rice brown plant hopper resistance through phenotypic reaction and genotypic analysis. Crop Prot. 2015, 78, 119. [Google Scholar] [CrossRef]
- Min, S.; Lee, S.W.; Choi, B.R.; Lee, S.H.; Kwon, D.H. Insecticide resistance monitoring and correlation analysis to select appropriate insecticides against Nilaparvata lugens (Stål), a migratory pest in Korea. J. Asia-Pac. Entomol. 2014, 7, 711–716. [Google Scholar] [CrossRef]
- Pati, P.; Jena, M.; Annamalai, M.; Behera, S.K.; Raghu, S.; Bhattacharya, S.; Sanghamitra, P.; Guru, P.P.; Meena, S. Screening of red rice accessions for resistance to brown planthopper Nilaparvata lugens Stål. ORYZA 2018, 55, 596–601. [Google Scholar] [CrossRef]
- Alagar, M.; Suresh, S.; Saravanan, P.A. Feeding behavior of Nilaparvata lugens on selected rice genotypes. Ann. Plant Prot. Sci. 2008, 6, 43–45. [Google Scholar]
- Catindig, J.L.A.; Arida, G.S.; Baehaki, S.E. Situation of planthoppers in Asia. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; Heong, K.L., Hardy, B., Eds.; International Rice Research Institute: Los Banos, Philippines, 2009; Volume 191, p. 220. [Google Scholar]
- Win, S.S.; Muhamad, R.; Ahmad, Z.A.M.; Adam, N.A. Population fluctuations of brown planthopper Nilaparvata lugens Stål and white backed planthopper Sogatella furcifera Horvath on rice. J. Entom. 2011, 8, 83–190. [Google Scholar]
- Sandhu, R.K.; Sarao, P.S. Evaluation of antixenosis resistance in wild rice accessions against brown planthopper, Nilaparvatalugens (Stål). Int. J. Trop. Insect Sci. 2020, 41, 65–73. [Google Scholar] [CrossRef]
- Jena, K.K.; Kim, S.M. Current status of brown planthopper (BPH) resistance and genetics. Rice 2010, 3, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Anant, A.; Govindharaj, G.P.P.; Gajendra, C.; Gowda, G.B.; Patil, N.; Annamalai, M.; Adak, T.; Rath, P.; Jena, M. Evaluation of Brown plant hopper Nilaparvata lugens (Stål.) resistance. Indian J. Entomol. 2021, 81, 223–225. [Google Scholar] [CrossRef]
- Balachiranjeevi, C.; Prahalada, G.D.; Mahender, A. Identification of a novel locus, Bph38(t), conferring resistance to brown planthopper (Nilaparvata lugens Stål.) using early backcross population in rice (Oryza sativa L.). Euphytica 2019, 215, 185. [Google Scholar] [CrossRef] [Green Version]
- Jena, K.K.; Hechanova, S.L.; Verdeprado, H.; Prahalada, G.D.; Kim, S.R. Development of 25 near-isogenic lines (NILs) with ten BPH resistance genes in rice (Oryza sativa L.): Production, resistance spectrum, and molecular analysis. Theor. Appl. Genet. 2017, 130, 2345–2360. [Google Scholar] [CrossRef]
- Jia, Y.; Gealy, D. Weedy red rice has novel sources of resistance to biotic stress. Crop J. 2018, 6, 443–450. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, W.L. Genetic and biochemical mechanisms of rice resistance to planthopper. Plant Cell Rep. 2016, 35, 1559–1572. [Google Scholar]
- Deepa, K.; Ariavanamkatha, P.M.; Murugesan, N. Biochemical bases of resistance to brown planthopper (Nilaparvata lugens) Stål) in different rice accessions. Agric. Sci. Dig. Res. J. 2016, 36, 102–105. [Google Scholar] [CrossRef] [Green Version]
- Heinrichs, E.A.; Medrano, F.G.; Rapusas, H.R. Genetic Evaluation for Insect Resistance in Rice; International Rice Research Institute: Los Banos, Philippines, 1985. [Google Scholar]
- Zieslinl, N.; Ben Zaken, R. Peroxidase activity and presence of phenolic substances in peduncles of rose flowers. Plant Physiol. Biochem. 1993, 31, 333–339. [Google Scholar]
- Hartree, E.F. Modern Methods of Plant Analysis; Springer Publ.: Berlin, Germany, 1955; Volume IV, p. 231. [Google Scholar]
- Hammerschmidt, R.; Nuckles, E.M.; Kuc, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Mayer, A.M.; Harel, E.; Shaul, R.B. Assay of catechol oxidase, a critical comparison of methods. Phytochemistry 1965, 5, 783–789. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the Superoxide Anion Radical in the Autoxidation of Pyrogallol and a Convenient Assay for Superoxide Dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Dekock, P.C.; Commisiong, K.; Farmer, V.C.; Inkson, H.E. Interrelationships of catalase, peroxidase, hematin and chlorophyll. Plant Physiol. 1960, 35, 599–604. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.; Rosebrough, N.J.; Lewis Farr, A.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1965, 193, 265–275. [Google Scholar] [CrossRef]
- Dai, W.; Zhang, K.-Q.; Duan, B.-W.; Sun, C.-X.; Zheng, K.-L.; Cai, R.; Zhuang, J.-Y. Rapid determination of silicon content in rice. Rice Sci. 2005, 12, 145–147. [Google Scholar]
- Muduli, L.; Pradhan, S.K.; Mishra, A.; Bastia, D.N.; Samal, K.C.; Agrawal, P.K.; Dash, M. Understanding Brown Planthopper Resistance in Rice: Genetics, Biochemical and Molecular Breeding Approaches. Rice Sci. 2021, 28, 532–546. [Google Scholar] [CrossRef]
- Senthil-Nathan, S.; Kalaivani, K.; Choi, M.Y.; Paik, C.H. Effects of jasmonic acid-induced resistance in rice on the brown planthopper, Nilaparvata lugens Stål (Homoptera: Delphacidae). Pestic. Biochem. Physiol. 2009, 95, 77–84. [Google Scholar] [CrossRef]
- Khush, G.S.; Brar, D.S. Genetics of resistance to insects in crop plants. Adv. Agron. 1991, 45, 223–274. [Google Scholar]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought from genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–364. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.W.; Tang, K.X. MAP Kinase cascades responding to environmental stress in plants. Acta Bot. Sin. Engl. Ed. 2004, 46, 127–136. [Google Scholar]
- Zheng, X.; Zhu, L.; He, G. Genetic and molecular understanding of host rice resistance and Nilaparvata lugens adaptation. Curr. Opin. Insect Sci. 2021, 45, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.N.; Wang, L.P.; He, Y.P.; Liang, J.G.; Tao, R.X. Expression of defense genes and activities of antioxidant enzymes in rice resistance to rice stripe virus and small brown planthopper. Plant Physiol. Biochem. 2011, 49, 744–751. [Google Scholar] [CrossRef]
- Duan, C.X.; Yu, J.J.; Bai, J.Y.; Zhu, Z.D.; Wang, X.M. Induced defense responses in rice plants against small brown planthopper infestation. Crop J. 2014, 2, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Belete, T. Defense mechanisms of plants to insect pests: From morphological to biochemical approach. Trends Tech. Sci. Res. 2018, 2, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Djamin, A.; Pathak, M.D. Role of silica in resistance to Asiatic rice borer, Chilo suppressalis Walker in rice varieties. J. Econ. Entomol. 1967, 60, 347–351. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Li, P.; Gong, S.; Yang, L.; Wen, L.; Hou, M. Defense Responses in Rice Induced by Silicon Amendment against Infestation by the Leaf Folder Cnaphalocrocis medinalis. PLoS ONE 2016, 11, e0153918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, R.; Uthamasamy, S. Feeding induced changes in phenolics and pathogenesis-related proteins: Implications in host resistance to Bemisia tabaci Genn and Helicoverpa armigera Hub. in tomato accessions. Pest Manag. Hortic. Ecosyst. 2004, 10, 95–106. [Google Scholar]
- Usha Rani, P.; Jyothsna, Y. Biochemical and enzymatic changes in rice as a mechanism of defense. Acta Physiol. Plant. 2010, 32, 695–701. [Google Scholar] [CrossRef]
- Duffey, S.S.; Stout, M.J. Antinutritive and toxic components of plant defense against insects. Arch. Insect Biochem. Physiol. 1996, 32, 3–37. [Google Scholar] [CrossRef]
- Saravanakumar, D.; Vijayakumar, C.; Kumar, N.; Samiyappan, R. PGPR Induced Defense Responses in Tea Plants against Blister Blight Disease. Crop Prot. 2007, 26, 556–565. [Google Scholar] [CrossRef]
- Saravanakumar, D.; Lavanya, N.; Muthumeena, K.; Raguchander, T.; Samiyappan, R. Fluorescent pseudomonad mixtures mediate disease resistance in rice plants against sheath rot (Sarocladium oryzae) disease. Biocontrol 2009, 54, 273–286. [Google Scholar] [CrossRef]
- Androcioli, H.G.; Hoshino, A.T.; Ventura, M.U.; Hata, F.T.; Brugnerotto, M.d.R.; Constantino, L.V.; Marques, F.D.A. Resistance of Common Bean Genotypes to the Broad Mite, Polyphagotarsonemus latus (Banks, 1904) (Acari: Tarsonemidae): Offspring Development and Biochemical Basis. Insects 2021, 12, 910. [Google Scholar] [CrossRef]
- Radjacommare, R.; Nandakumar, R.; Kandan, A. Pseudomonas fluorescens based bio-formulation for the management of sheath blight and leaf folder insect in rice. Crop Prot. 2002, 21, 671–677. [Google Scholar] [CrossRef]
- Alagar, M.; Suresh, S.; Saravanakumar, D.; Samiyappan, R. Feeding-induced changes in defence enzymes and PR proteins and their implications in host resistance to Nilaparvata lugens. J. Appl. Entomol. 2009, 134, 123–131. [Google Scholar] [CrossRef]
- Taggar, G.K.; Gill, R.S.; Gupta, A.K.; Sandhu, S. Fluctuations in peroxidase and catalase activities of resistant and susceptible black gram (Vigna mungo (L.) Hepper) genotypes elicited by Bemisia tabaci (Gennadius) feeding. Plant Signal. Behav. 2012, 7, 1321–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lima Toledo, C.A.; da Silva Ponce, F.; Oliveira, M.D.; Aires, E.S.; Seabra Júnior, S.; Lima, G.P.P.; de Oliveira, R.C. Change in the Physiological and Biochemical Aspects of Tomato Caused by Infestation by Cryptic Species of Bemisia tabaci MED and MEAM1. Insects 2021, 12, 1105. [Google Scholar] [CrossRef] [PubMed]
- Walling, L.L. Recycling or regulation? The role of amino-terminal modifying enzymes. Curr. Opin. Plant Biol. 2006, 9, 227–233. [Google Scholar] [CrossRef]
- Isman, M.B.; Duffey, S.S. Toxicity of tomato phenolic compounds to the fruit worm, Heliothis zea. Entomol. Exp. Appl. 1982, 31, 370–376. [Google Scholar] [CrossRef]
- Isman, M.B.; Duffey, S.S. Phenolic compounds in foliage of commercial tomato cultivars as growth inhibitors to the fruitworms, Heliothis zea. J. Am. Soc. Hortic. Sci. 1982, 107, 167–170. [Google Scholar] [CrossRef]
- Saravanakumar, D.; Muthumeena, B.; Lavanya, N.; Suresh, S.; Rajendran, L.; Raguchander, T.; Samiyappan, R. Pseudomonas induced defense molecules in rice against leaf folder (Cnephalocrocis medinalis) pest. Pest Manag. Sci. 2007, 63, 714–721. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Ahmad, T. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.Z.; Hua, B.Z.; Zhang, F. Induction of the activities of antioxidative enzymes and the levels of malondialdehyde in cucumber seedlings as a consequence of Bemisia tabaci (Hemiptera: Aleyrodidae) infestation. Arthropod-Plant Interact. 2008, 2, 209–213. [Google Scholar] [CrossRef]
- Bednarski, W.; Borowiak-Sobkowiak, B.; Wilkaniec, B.; Samardakiewicz, S.; Morkunas, I. Oxidative stress in pea seedling leaves in response to Acyrthosiphon pisum infestation. Phytochemistry 2013, 93, 49–62. [Google Scholar]
- Jannoey, P.; Channel, D.; Kotcharek, J.; Pongprasert, W.; Nomura, M. Expression Analysis of Genes related to rice resistance against brown planthopper, Nilaparvata lugens. Rice Sci. 2017, 24, 163–172. [Google Scholar] [CrossRef]
- Zhao, H.; Sun, X.; Xue, M.; Zhang, X.; Li, Q. Antioxidant Enzyme Responses Induced by Whiteflies in Tobacco Plants in Defense against Aphids: Catalase May Play a Dominant Role. PLoS ONE 2016, 11, e0165454. [Google Scholar] [CrossRef] [Green Version]
- Afiyanti, M.; Chen, H.J. Catalase activity is modulated by calcium and calmodulin in detached mature leaves of sweet potato. J. Plant Physiol. 2014, 171, 35–47. [Google Scholar] [CrossRef]
- Overmyer, K.; Brosche, M.; Kangasjärvi, J. Reactive oxygen species and hormonal control of cell death. Trends Plant Sci. 2003, 8, 335–342. [Google Scholar] [CrossRef]
- Chen, Z.; Silva, H.; Klessig, D.F. Active oxygen species in the induction of plant systemic acquired resistance by salicylic acid. Science 1993, 262, 1883–1886. [Google Scholar] [CrossRef]
- Mohase, L.; van der Westhuizen, A. Salicylic acid is involved in resistance responses in the Russian wheat aphid–wheat interaction. J. Plant Physiol. 2002, 159, 585–590. [Google Scholar] [CrossRef]
- Bi, J.L.; Felton, G.W. Foliar oxidative stress and insect herbivory: Primary compounds, secondary metabolites and reactive oxygen species as components of induced resistance. J. Chem. Ecol. 1995, 21, 1511–1530. [Google Scholar] [CrossRef]
- Alagar, M.; Suresh, S.; Samiyappan, R.; Saravanakumar, D. Reaction of resistant and susceptible rice genotypes against brown plant hopper (Nilaparvata lugens). Phytoparasitica 2007, 35, 346. [Google Scholar] [CrossRef]
- Ma, C.; Gui, W.; Zhang, Y.; Shah, A.Z.; Xu, G.; Yang, G. Combined Physio-biochemical and Transcriptome Analyses Illuminate the Resistance Reponse of rice priming with Decoyinine against Nilaparvata lugens. Agronomy 2022, 12, 3098. [Google Scholar] [CrossRef]
- Edwards, P.J.; Wratten, S.D. Wound induced defenses in plants and their consequences for patterns of insect grazing. Oecologia 1983, 59, 88–93. [Google Scholar] [CrossRef]
- Padmavathi, C.; Katti, G.; Padmakumari, A.P.; Voleti, S.R.; Subba Rao, L.V. The effect of leaf folder Cnaphalocrocis medinalis (Guenee) (Lepidoptera: Pyralidae) injury on the plant physiology and yield loss in rice. J. Appl. Entomol. 2013, 137, 249–256. [Google Scholar] [CrossRef]
- Yuan, H.; Chen, X.; Zhu, L.; He, G. Identification of genes responsive to brown planthopper Nilaparvatalu gens Stål (Homoptera: Delphacidae) feeding in rice. Planta 2005, 221, 105–112. [Google Scholar] [CrossRef]
- Nayak, A.; Baig, M.J.; Mohapatra, P.K.; Behera, K.S. Effect of insect feeding on biochemical changes in rice plant. J. Entomol. Zool. Stud. 2019, 7, 138–142. [Google Scholar]
- Jayasimha, G.T.; Nalini, R.; Chinniah, C.; Muthamilan, M.; Mini, M.L. Evaluation of biochemical constituents in healthy and brown planthopper, Nilaparvata lugens (Stål.) (Hemiptera: Delphacidae) damaged rice plants. Curr. Biot. 2015, 9, 129–136. [Google Scholar]
- Yang, L.; Han, Y.; Li, P.; Li, F.; Ali, S.; Hou, M. Silicon amendment is involved in the induction of plant defense responses toa phloem feeder. Sci. Rep. 2017, 7, 4232. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Nikolic, M.; Belanger, R.; Gong, H.; Song, A. Silicon and insect pest resistance. In Silicon in Agriculture, from Theory to Practice; Liang, Y., Nikolic, M., Belanger, R., Geong, H., Song, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 197–207. [Google Scholar]
- Massey, F.P.; Ennos, A.R.; Hartley, S.E. Silica in grasses as a defence against insect herbivores: Contrasting effects on folivores and a phloem feeder. J. Anim. Ecol. 2006, 75, 595–596. [Google Scholar] [CrossRef]
- Massey, F.P.; Hartley, S.E. Physical defences wear you down: Progressive and irreversible impacts of silica on insect herbivores. J. Anim. Ecol. 2009, 78, 281–291. [Google Scholar] [CrossRef]
- Han, Y.; Lei, W.; Wen, L.; Hou, M. Silicon- mediated resistance in a susceptible rice variety to the rice leaf folder, Cnaphalocrocis medinalis Guenée (Lepidoptera: Pyralidae). PLoS ONE 2015, 10, e01205. [Google Scholar] [CrossRef]
- Yang, L.; Han, Y.; Li, P.; Wen, L.; Hou, M. Silicon amendment to rice plants impairs sucking behaviors and population growth in the phloem feeder Nilaparvata lugens (Hemiptera: Delphacidae). Sci. Rep. 2017, 7, 1101. [Google Scholar] [CrossRef] [Green Version]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pati, P.; Jena, M.; Bhattacharya, S.; Behera, S.K.; Pal, S.; Shivappa, R.; Dhar, T. Biochemical Defense Responses in Red Rice Genotypes Possessing Differential Resistance to Brown Planthopper, Nilaparvata lugens (Stål). Insects 2023, 14, 632. https://doi.org/10.3390/insects14070632
Pati P, Jena M, Bhattacharya S, Behera SK, Pal S, Shivappa R, Dhar T. Biochemical Defense Responses in Red Rice Genotypes Possessing Differential Resistance to Brown Planthopper, Nilaparvata lugens (Stål). Insects. 2023; 14(7):632. https://doi.org/10.3390/insects14070632
Chicago/Turabian StylePati, Prajna, Mayabini Jena, Swarnali Bhattacharya, Santhosh Kumar Behera, Subhajit Pal, Raghu Shivappa, and Tapamay Dhar. 2023. "Biochemical Defense Responses in Red Rice Genotypes Possessing Differential Resistance to Brown Planthopper, Nilaparvata lugens (Stål)" Insects 14, no. 7: 632. https://doi.org/10.3390/insects14070632