
Trophic Interactions of Ceutorhynchinae spp. (Coleoptera: Curculionidae) with Their Host Plants (Brassicaceae) and Their Parasitoids in the Agroecosystem of Quebec, Canada

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Emergence Boxes
2.3. Parasitoids and Weevil Identification
3. Results
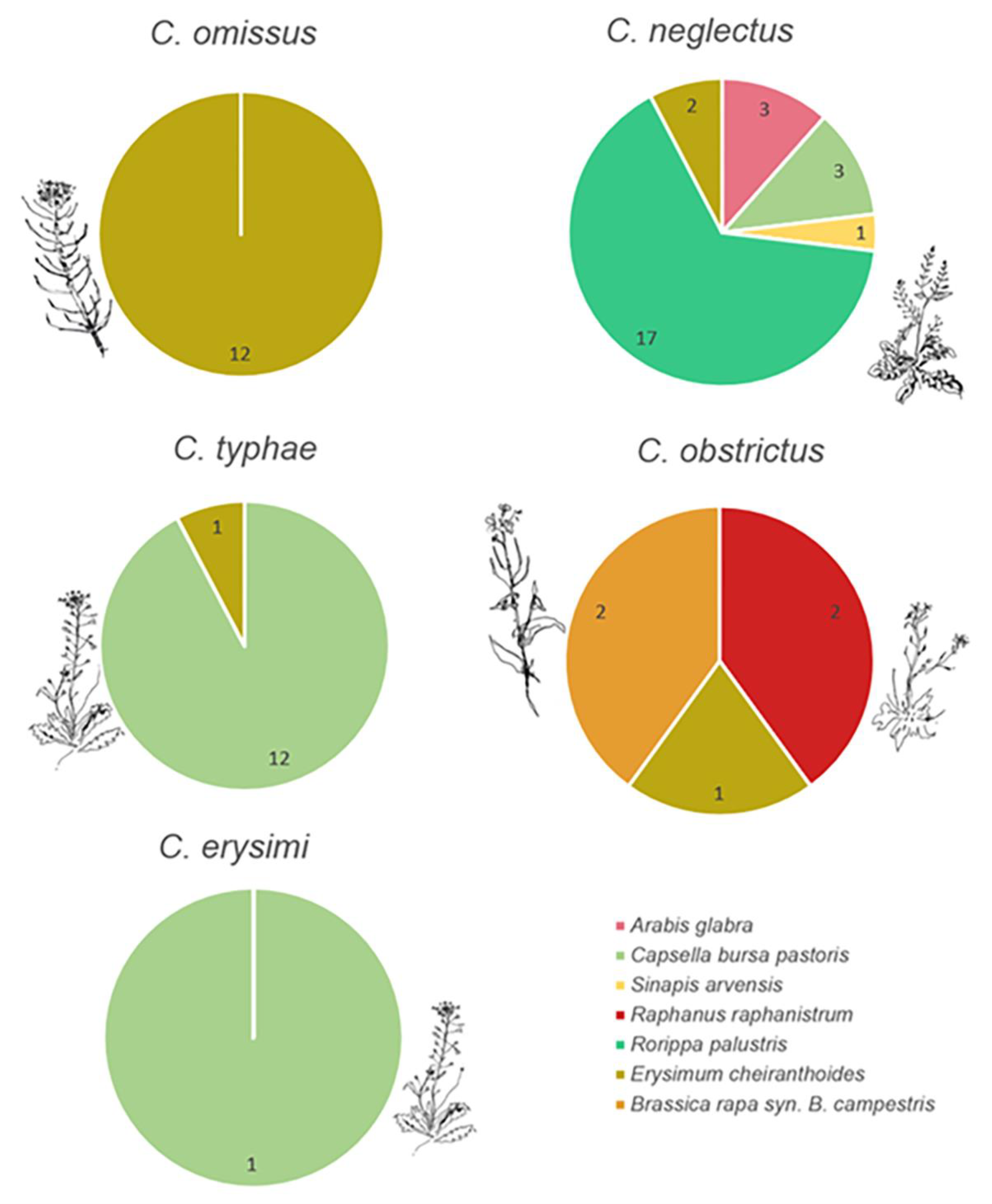
3.1. Ceutorhynchus and Their Host Plant
3.2. Parasitoids Collected
3.3. Ceutorhynchus Trophic Associations
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Korotyaev, B.A. Geographical distribution of the weevil subfamily Ceutorhynchinae (Coleoptera, Curculionidae). Entomol. Rev. 2008, 88, 928–947. [Google Scholar] [CrossRef]
- Letsch, H.; Gottsberger, B.; Metzl, C.; Astrin, J.; Friedman, A.L.L.; McKenna, D.D.; Fiedler, K. Climate and host-plant associations shaped the evolution of ceutorhynch weevils throughout the Cenozoic. Evolution 2018, 72, 1815–1828. [Google Scholar] [CrossRef] [Green Version]
- Colonnelli, E. Catalogue of Ceutorhynchinae of the World, with a Key to Genera (Coleoptera: Curculionidae); Argania Editio: Barcelona, Spain, 2004; 124p. [Google Scholar]
- Anderson, R.S. Curculionidae. In American Beetles; CRC Press: Boca Raton, FL, USA, 2002; Volume 2, pp. 722–815. [Google Scholar]
- Mason, P.G.; Miall, J.H.; Bouchard, P.; Brauner, A.; Gillespie, D.R.; Gibson, G.A.P. The parasitoid communities associated with Ceutorhynchus species (Coleoptera: Curculionidae) in Ontario and Québec, Canada. Can. Entomol. 2014, 146, 224–235. [Google Scholar] [CrossRef]
- Frank van Veen, F.J.; Morris, R.J.; Godfray, H.C.J. Apparent Competition, Quantitative food webs, and the Structure of phytophagous insect communities. Annu. Rev. Entomol. 2006, 51, 187–208. [Google Scholar] [CrossRef]
- Miličić, M.; Popov, S.; Branco, V.V.; Cardoso, P. Insect threats and conservation through the lens of global experts. Conserv. Lett. 2021, 14, e12814. [Google Scholar] [CrossRef]
- Wagner, D.L.; Grames, E.M.; Forister, M.L.; Berenbaum, M.R.; Stopak, D. Insect decline in the Anthropocene: Death by a thousand cuts. Proc. Natl. Acad. Sci. USA 2021, 118, e2023989118. [Google Scholar] [CrossRef]
- Desroches, C.; Bouchard, P.; Labrie, G.; Lucas, E. Assemblage of Ceutorhynchinae Weevils Associated with Brassicaceae in Quebec (Canada) Agroecosystems. Environ. Èntomol. 2022, 52, 18–30. [Google Scholar] [CrossRef]
- Van Driesche, R.; Hoddle, M.; Center, T.D. Control of Pests and Weeds by Natural Enemies: An Introduction to Biological Control, 1st ed.; Blackwell Pub: Hoboken, NJ, USA, 2008; 484p. [Google Scholar]
- Norris, R.F.; Kogan, M. Interactions between weeds, arthropod pests, and their natural enemies in managed ecosystems. Weed Sci. 2000, 48, 94–158. [Google Scholar] [CrossRef]
- Vidal, M.C.; Murphy, S.M. Bottom-up vs. top-down effects on terrestrial insect herbivores: A meta-analysis. Ecol. Lett. 2018, 21, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Vidal, S. Identification of Hymenopterous Parasitoids Associated with Oilseed Rape Pests. In Biocontrol of Oilseed Rape Pests; Alford, D.V., Ed.; Blackwell Science Ltd.: Hoboken, NJ, USA, 2003; pp. 161–180. [Google Scholar] [CrossRef]
- Mason, P.G.; Miall, J.H.; Bouchard, P.; Gillespie, D.R.; Broadbent, A.B.; Gibson, G.A.P. The parasitoid communities associated with an invasive canola pest, Ceutorhynchus obstrictus (Coleoptera: Curculionidae), in Ontario and Quebec, Canada. Can. Entomol. 2011, 143, 524–537. [Google Scholar] [CrossRef]
- Gibson, G.A.P.; Huber, J.T.; Woolley, J.B. Annoted Keys to the Genera of Nearctic Chalcidoidea (Hymenoptera); NRC Research Press: Ottawa, ON, Canada, 1997; 794p. [Google Scholar]
- Marie-Victorin; Brouillet, L.; Rouleau, E.; Goulet, I.; Hay, S. Flore Laurentienne; Presses de l’Université de Montréal: Montréa, QC, Canada, 2001; 1084p. [Google Scholar]
- Bouchard, P.; Lesage, L.; Goulet, H.; Bostanian, N.J.; Vincent, C.; Zmudzinska, A.; Lasnier, J. Weevil (Coleoptera: Curculionoidea) Diversity and Abundance in Two Quebec Vineyards. Ann. Entomol. Soc. Am. 2005, 98, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Brodeur, J.; Leclerc, L.-A.; Fournier, M.; Roy, M. Cabbage seedpod weevil (Coleoptera: Curculionidae): New pest of canola in northeastern North America. Can. Entomol. 2001, 133, 709–711. [Google Scholar] [CrossRef]
- Mason, P.G.; Baute, T.; Olfert, O.; Roy, M. Cabbage Seedpod Weevil, Ceuthorhyncus obstrictus (Marsham) (Coleoptera: Curculionidae) in Ontario and Quebec. J. Entomol. Soc. Ont. 2003, 134, 107–113. [Google Scholar]
- Kovács, G.; Kaasik, R.; Metspalu, L.; Williams, I.H.; Luik, A.; Veromann, E. Could Brassica rapa, Brassica juncea and Sinapis alba facilitate the control of the cabbage seed weevil in oilseed rape crops? Biol. Control 2013, 65, 124–129. [Google Scholar] [CrossRef]
- Simard, M.-J.; Laforest, M.; Martin, S.; Cuerrier, M.-E.; Miville, D.; Marcoux, J.; Flores-Meja, S. Malherbologie, Distribution de la Moutarde des Oiseaux Résistante au Glyphosate au Québec, Agri Réseau. 2021, Bulletin D’information No 5. Available online: https://www.agrireseau.net/documents/106928/malherbologie-bulletin-d_information-no-5-distribution-de-la-moutarde-des-oiseaux-resistante-au-glyphosate-au-quebec?a=1&r=moutarde+des+oiseaux (accessed on 20 April 2023).
- Cárcamo, H.A.; Dunn, R.; Dosdall, L.M.; Olfert, O. Managing cabbage seedpod weevil in canola using a trap crop—A commercial field scale study in western Canada. Crop Prot. 2007, 26, 1325–1334. [Google Scholar] [CrossRef]
- Barari, H.; Cook, S.M.; Clark, S.J.; Williams, I.H. Effect of a turnip rape (Brassica rapa) trap crop on stem-mining pests and their parasitoids in winter oilseed rape (Brassica napus). BioControl 2005, 50, 69–86. [Google Scholar] [CrossRef]
- Buechi, R. Investigations on the use of turnip rape as trap plant to control oilseed rape pests. Bull. Int. Union Biol. Sci. West Paleartic Reg. Sect. 1990, 13, 32–39. [Google Scholar]
- Dosdall, L.M.; Moisey, D.W.A. Developmental Biology of the Cabbage Seedpod Weevil, Ceutorhynchus obstrictus (Coleoptera: Curculionidae), in Spring Canola, Brassica napus, in Western Canada. Ann. Entomol. Soc. Am. 2004, 97, 458–465. [Google Scholar] [CrossRef]
- Kumar, S.; Bhowmick, M.K.; Ray, P. Weeds as alternate and alternative hosts of crop pests. Indian J. Weed Sci. 2021, 53, 14–29. [Google Scholar] [CrossRef]
- Dosdall, L.M.; Gibson, G.A.P.; Olfert, O.O.; Mason, P.G. Responses of Chalcidoidea (Hymenoptera) parasitoids to invasion of the cabbage seedpod weevil (Coleoptera: Curculionidae) in western Canada. Biol. Invasions 2009, 11, 109–125. [Google Scholar] [CrossRef]
- Veromann, E.; Williams, I.H.; Kaasik, R.; Luik, A. Potential of parasitoids control populations of the weevil Ceutorhynchus obstrictus (Marsham) on winter oilsead rape. Int. J. Pest Manag. 2010, 57, 85–92. [Google Scholar] [CrossRef]
- Haye, T.; Olfert, O.; Weiss, R.; Mason, P.G.; Gibson, G.; Gariepy, T.D.; Gillespie, D.R. Bioclimatic analyses of Trichomalus perfectus and Mesopolobus morys (Hymenoptera: Pteromalidae) distributions, two potential biological control agents of the cabbage seedpod weevil in North America. Biol. Control 2018, 124, 30–39. [Google Scholar] [CrossRef]
- D’Ottavio, M.; Boquel, S.; Labrie, G.; Lucas, E. Landscape Effects on the Cabbage Seedpod Weevil, Ceutorhynchus obstrictus (Coleoptera: Curculionidae), and on Its Parasitoid, Trichomalus perfectus (Hymenoptera: Pteromalidae), in Canola. Insects 2023, 14, 327. [Google Scholar] [CrossRef] [PubMed]
- Mulatu, B.; Applebaum, S.W.; Coll, M. A recently acquired host plant provides an oligophagous insect herbivore with enemy-free space. Oikos 2004, 107, 231–238. [Google Scholar] [CrossRef]
- Jeffries, M.J.; Lawton, J.H. Enemy free space and the structure of ecological communities. Biol. J. Linn. Soc. 1984, 23, 269–286. [Google Scholar] [CrossRef]
- Haye, T.; Mason, P.G.; Dosdall, L.M.; Kuhlmann, U. Ceutorhynchus obstrictus (Marsham), Cabbage Seedpod Weevil (Coleoptera: Curculionidae), 1st ed.; P Biological Control Programms in Canada 2001–2010; Mason, P.G., Gillespie, D.R., Eds.; CABI: Wallingford, UK, 2013; pp. 119–128. [Google Scholar]
- Haye, T.; Mason, P.G.; Gillespie, D.R.; Miall, J.H.; Gibson, G.A.P.; Diaconu, A.; Brauner, A.M.; Kuhlmann, U. Determining the host specificity of the biological control agent Trichomalus perfectus (Hymenoptera: Pteromalidae): The importance of ecological host range. Biocontrol Sci. Technol. 2015, 25, 21–47. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. neglectus | C. omissus | C. typhae | Unidentified Ceutorhynchus | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Rorippa palustris Marsh Yellow Cress | Sinapis arvensis Wild Mustard | Erysimum cheiranthoides Wormseed Mustard | Arabis glabra Tower Rockcress | Erysimum cheiranthoides Wormseed Mustard | Capsella bursa-pastoris Shepherd’s Purse | Rorippa palustris Marsh Yellow Cress | Capsella bursa-pastoris Shepherd’s Purse | Erysimum cheiranthoides Wormseed Mustard | |
| Pteromalidae | |||||||||
| Chlorocytus species | 1 | 12 | 15 | 2 | |||||
| Pteromalus species | 1 | 1 | 1 | 1 | 11 | 7 | 1 | 4 | 22 |
| Trichomalus lucidus (Walker) | 1 | ||||||||
| Trichomalus perfectus (Walker) | 1 | 1 | |||||||
| Trichomalus species | 1 | ||||||||
| Unidentified Pteromalidae species | 2 | 7 | |||||||
| Elachertodomyia species | 3 | ||||||||
| Eulophidae | |||||||||
| Necremnus duplicatus Gahan | 9 | ||||||||
| Unidentified Eulophidae species | 2 | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Desroches, C.; Moisan-De Serres, J.; Rodrigue, É.; Labrie, G.; Lucas, É. Trophic Interactions of Ceutorhynchinae spp. (Coleoptera: Curculionidae) with Their Host Plants (Brassicaceae) and Their Parasitoids in the Agroecosystem of Quebec, Canada. Insects 2023, 14, 607. https://doi.org/10.3390/insects14070607
Desroches C, Moisan-De Serres J, Rodrigue É, Labrie G, Lucas É. Trophic Interactions of Ceutorhynchinae spp. (Coleoptera: Curculionidae) with Their Host Plants (Brassicaceae) and Their Parasitoids in the Agroecosystem of Quebec, Canada. Insects. 2023; 14(7):607. https://doi.org/10.3390/insects14070607
Chicago/Turabian StyleDesroches, Claudine, Joseph Moisan-De Serres, Émilien Rodrigue, Geneviève Labrie, and Éric Lucas. 2023. "Trophic Interactions of Ceutorhynchinae spp. (Coleoptera: Curculionidae) with Their Host Plants (Brassicaceae) and Their Parasitoids in the Agroecosystem of Quebec, Canada" Insects 14, no. 7: 607. https://doi.org/10.3390/insects14070607