
Diversity and Dynamics of Bacterial Communities in the Digestive and Excretory Systems across the Life Cycle of Leafhopper, Recilia dorsalis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Tissue Sample Collection
2.3. DNA Extraction and PCR Amplification
2.4. High-Throughput Sequencing and Analysis
2.5. Statistical Analysis
3. Results
3.1. General Characteristics of the Sequencing Data
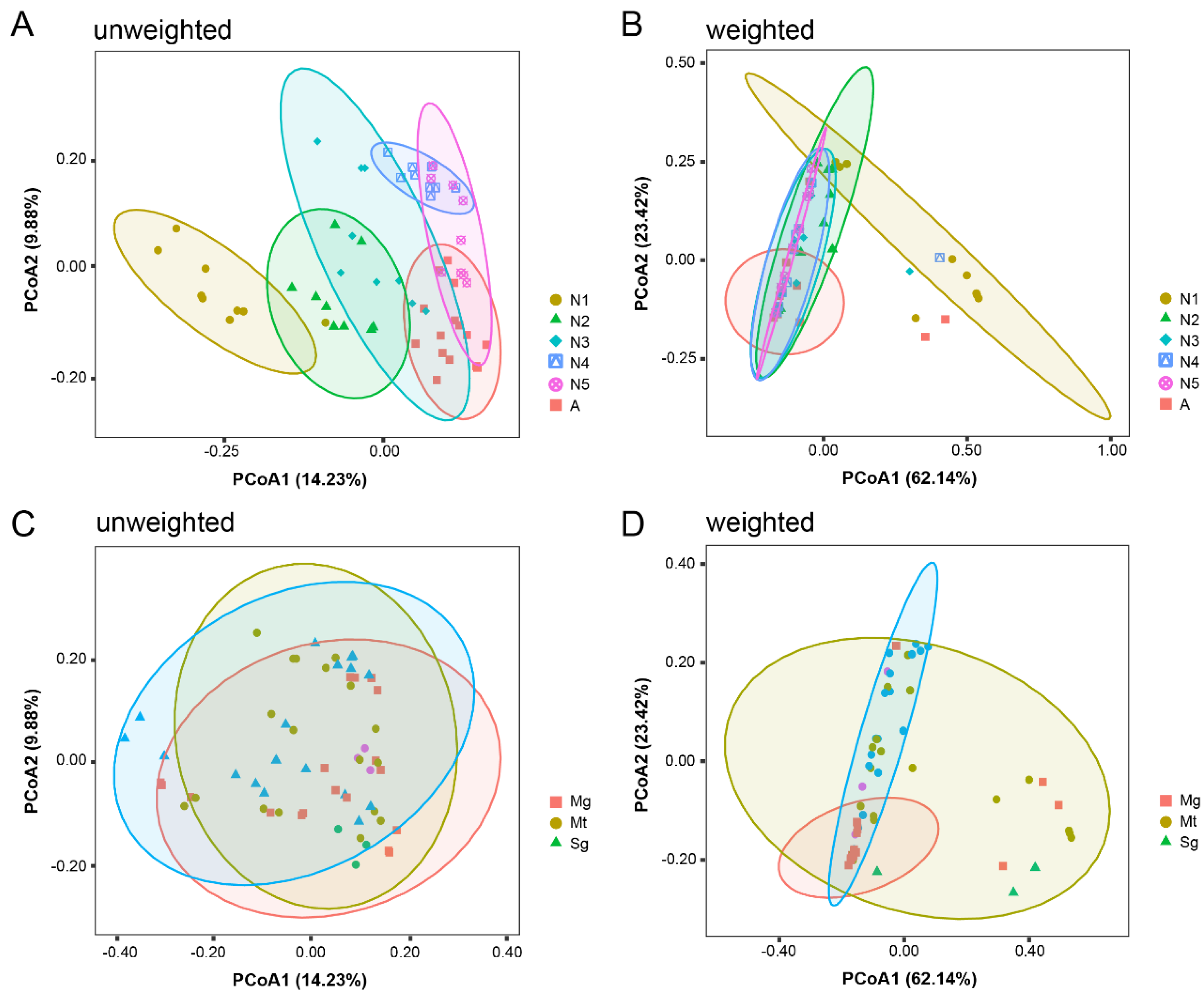
3.2. Diversity of Bacterial Communities in Different Tissues throughout the Life Cycle of R. dorsalis
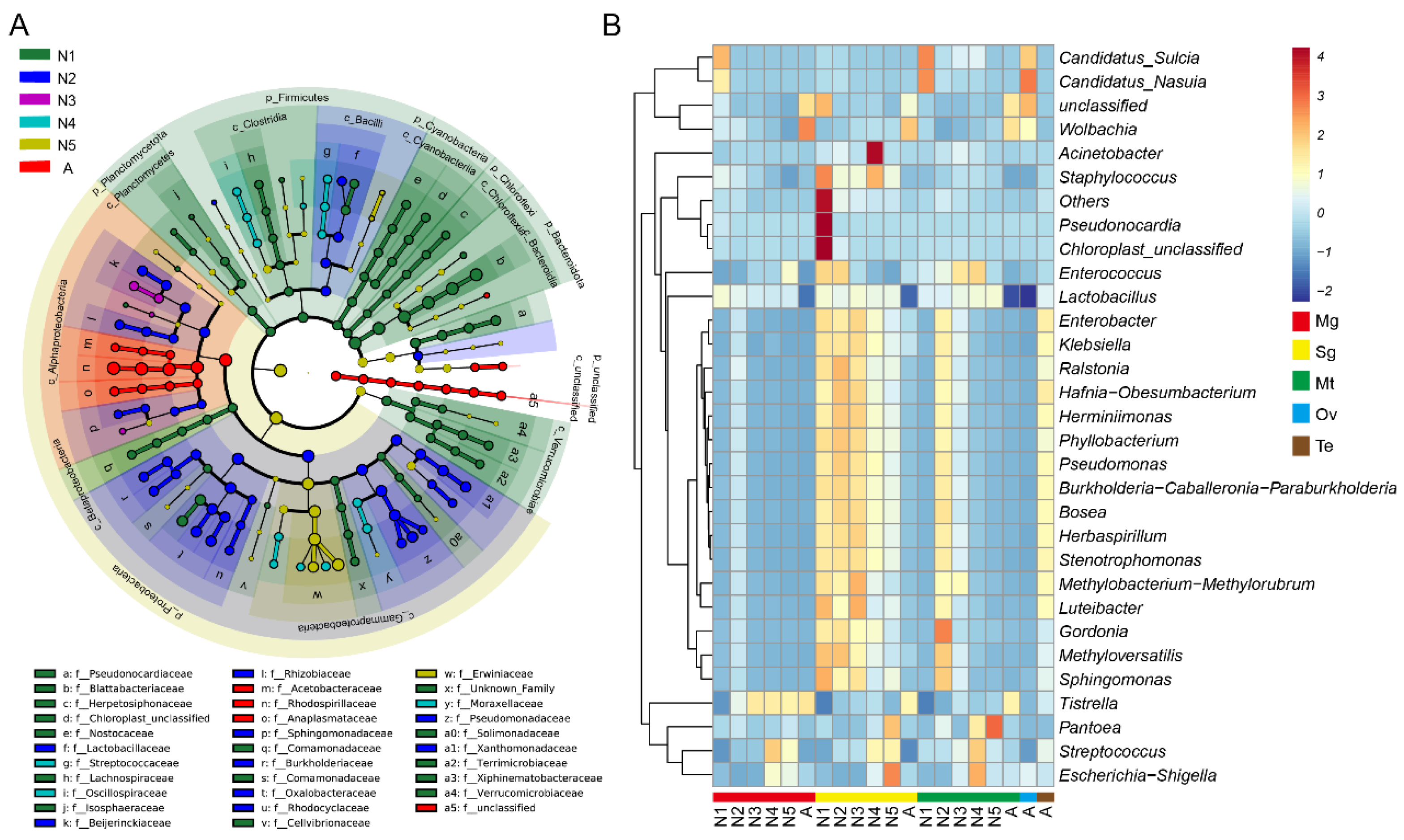
3.3. Bacterial Community at Different Developmental Stages and in Various Tissues of R. dorsalis
3.4. Significantly Different Bacterial Communities at Different Developmental Stages and in Various Tissues of R. dorsalis
3.5. Functional Analysis of Bacterial Communities at Different Developmental Stages and in Various Tissues of R. dorsalis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Basset, Y.; Cizek, L.; Cuénoud, P.; Didham, R.K.; Guilhaumon, F.; Missa, O.; Novotny, V.; Ødegaard, F.; Roslin, T.; Schmidl, J. Arthropod diversity in a tropical forest. Science 2012, 338, 1481–1484. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. The molecular basis of bacterial–insect symbiosis. J. Mol. Biol. 2014, 426, 3830–3837. [Google Scholar] [CrossRef] [Green Version]
- Ankrah, N.Y.; Douglas, A.E. Nutrient factories: Metabolic function of beneficial microorganisms associated with insects. Environ. Microbiol. 2018, 20, 2002–2011. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. The microbial dimension in insect nutritional ecology. Funct. Ecol. 2009, 23, 38–47. [Google Scholar] [CrossRef]
- Gaio, A.d.O.; Gusmão, D.S.; Santos, A.V.; Berbert-Molina, M.A.; Pimenta, P.F.; Lemos, F.J. Contribution of midgut bacteria to blood digestion and egg production in Aedes aegypti (Diptera: Culicidae) (L.). Parasites Vectors 2011, 4, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brummel, T.; Ching, A.; Seroude, L.; Simon, A.F.; Benzer, S. Drosophila lifespan enhancement by exogenous bacteria. Proc. Natl. Acad. Sci. USA 2004, 101, 12974–12979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosengaus, R.B.; Zecher, C.N.; Schultheis, K.F.; Brucker, R.M.; Bordenstein, S.R. Disruption of the termite gut microbiota and its prolonged consequences for fitness. Appl. Environ. Microbiol. 2011, 77, 4303–4312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, H.-S.; Song, S.-L.; Chua, K.-O.; Lim, P.-E. High diversity of bacterial communities in developmental stages of Bactrocera carambolae (Insecta: Tephritidae) revealed by Illumina MiSeq sequencing of 16S rRNA gene. Curr. Microbiol. 2017, 74, 1076–1082. [Google Scholar] [CrossRef]
- Shao, Y.; Chen, B.; Sun, C.; Ishida, K.; Hertweck, C.; Boland, W. Symbiont-derived antimicrobials contribute to the control of the lepidopteran gut microbiota. Cell Chem. Biol. 2017, 24, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Johnston, P.R.; Rolff, J. Host and symbiont jointly control gut microbiota during complete metamorphosis. PLoS Pathog. 2015, 11, e1005246. [Google Scholar] [CrossRef] [Green Version]
- Cirimotich, C.M.; Dong, Y.; Clayton, A.M.; Sandiford, S.L.; Souza-Neto, J.A.; Mulenga, M.; Dimopoulos, G. Natural microbe-mediated refractoriness to Plasmodium infection in Anopheles gambiae. Science 2011, 332, 855–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirimotich, C.M.; Ramirez, J.L.; Dimopoulos, G. Native microbiota shape insect vector competence for human pathogens. Cell Host Microbe 2011, 10, 307–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Huang, L.; Mao, Q.; Wei, J.; Li, J.; Zhao, Y.; Zhang, Q.; Jia, D.; Wei, T. Interaction of viral pathogen with porin channels on the outer membrane of insect bacterial symbionts mediates their joint transovarial transmission. Philos. Trans. R. Soc. B 2019, 374, 20180320. [Google Scholar] [CrossRef] [Green Version]
- Jia, D.; Mao, Q.; Chen, Y.; Liu, Y.; Chen, Q.; Wu, W.; Zhang, X.; Chen, H.; Li, Y.; Wei, T. Insect symbiotic bacteria harbour viral pathogens for transovarial transmission. Nat. Microbiol. 2017, 2, 17025. [Google Scholar] [CrossRef] [PubMed]
- García-Fraile, P. Roles of bacteria in the bark beetle holobiont–how do they shape this forest pest? Ann. Appl. Biol. 2018, 172, 111–125. [Google Scholar] [CrossRef]
- Douglas, A.E. Multiorganismal insects: Diversity and function of resident microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Moran, N.A.; McCutcheon, J.P.; Nakabachi, A. Genomics and evolution of heritable bacterial symbionts. Annu. Rev. Genet. 2008, 42, 165–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, N.A.; Bennett, G.M. The tiniest tiny genomes. Annu. Rev. Microbiol. 2014, 68, 195–215. [Google Scholar] [CrossRef]
- Oliver, K.M.; Degnan, P.H.; Burke, G.R.; Moran, N.A. Facultative symbionts in aphids and the horizontal transfer of ecologically important traits. Annu. Rev. Entomol. 2010, 55, 247–266. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Hulcr, J.; Sun, J. The role of symbiotic microbes in insect invasions. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 487–505. [Google Scholar] [CrossRef]
- Adair, K.L.; Douglas, A.E. Making a microbiome: The many determinants of host-associated microbial community composition. Curr. Opin. Microbiol. 2017, 35, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Girard, M.; Luis, P.; Valiente Moro, C.; Minard, G. Crosstalk between the microbiota and insect postembryonic development. Trends Microbiol. 2023, 31, 181–196. [Google Scholar] [CrossRef] [PubMed]
- Moll, R.M.; Romoser, W.S.; Modrakowski, M.C.; Moncayo, A.C.; Lerdthusnee, K. Meconial Peritrophic Membranes and the Fate of Midgut Bacteria During Mosquito (Diptera: Culicidae) Metamorphosis. J. Med. Entomol. 2001, 38, 29–32. [Google Scholar] [CrossRef]
- Juma, E.O.; Allan, B.F.; Kim, C.-H.; Stone, C.; Dunlap, C.; Muturi, E.J. Effect of life stage and pesticide exposure on the gut microbiota of Aedes albopictus and Culex pipiens L. Sci. Rep. 2020, 10, 9489. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.N.A.; Ng, P.; Douglas, A.E. Low-diversity bacterial community in the gut of the fruitfly Drosophila melanogaster. Environ. Microbiol. 2011, 13, 1889–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Teh, B.-S.; Sun, C.; Hu, S.; Lu, X.; Boland, W.; Shao, Y. Biodiversity and activity of the gut microbiota across the life history of the insect herbivore Spodoptera littoralis. Sci. Rep. 2016, 6, 29505. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Sun, S.; Yang, X.; Cheng, J.; Wei, H.; Li, Z.; Michaud, J.; Liu, X. Variability of gut microbiota across the life cycle of Grapholita molesta (Lepidoptera: Tortricidae). Front. Microbiol. 2020, 11, 1366. [Google Scholar] [CrossRef]
- Sudakaran, S.; Salem, H.; Kost, C.; Kaltenpoth, M. Geographical and ecological stability of the symbiotic mid-gut microbiota in European firebugs, Pyrrhocoris apterus (Hemiptera, Pyrrhocoridae). Mol. Ecol. 2012, 21, 6134–6151. [Google Scholar] [CrossRef]
- Brown, J.J.; Rodríguez-Ruano, S.M.; Poosakkannu, A.; Batani, G.; Schmidt, J.O.; Roachell, W.; Zima, J.; Hypša, V.; Nováková, E. Ontogeny, species identity, and environment dominate microbiome dynamics in wild populations of kissing bugs (Triatominae). Microbiome 2020, 8, 146. [Google Scholar] [CrossRef]
- Ren, Z.; Zhang, Y.; Cai, T.; Mao, K.; Xu, Y.; Li, C.; He, S.; Li, J.; Wan, H. Dynamics of microbial communities across the life stages of Nilaparvata lugens (Stål). Microb. Ecol. 2022, 83, 1049–1058. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, T.; Chen, B.; Zhou, G. Transmission biology of rice stripe mosaic virus by an efficient insect vector Recilia dorsalis (Hemiptera: Cicadellidae). Front. Microbiol. 2017, 8, 2457. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Lu, C.; Li, M.; Wu, W.; Zhou, G.; Wei, T. Adverse effects of rice gall dwarf virus upon its insect vector Recilia dorsalis (Hemiptera: Cicadellidae). Plant Dis. 2016, 100, 784–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCutcheon, J.P.; Moran, N.A. Functional convergence in reduced genomes of bacterial symbionts spanning 200 My of evolution. Genome Biol. Evol. 2010, 2, 708–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, H.; Watanabe, K.; Kawai, S.; Yukuhiro, F.; Miyoshi, T.; Tomizawa, M.; Koizumi, Y.; Nikoh, N.; Fukatsu, T. Bacteriome-associated endosymbionts of the green rice leafhopper Nephotettix cincticeps (Hemiptera: Cicadellidae). Appl. Entomol. Zool. 2012, 47, 217–225. [Google Scholar] [CrossRef]
- Iasur-Kruh, L.; Weintraub, P.G.; Mozes-Daube, N.; Robinson, W.E.; Perlman, S.J.; Zchori-Fein, E. Novel Rickettsiella bacterium in the leafhopper Orosius albicinctus (Hemiptera: Cicadellidae). Appl. Environ. Microbiol. 2013, 79, 4246–4252. [Google Scholar] [CrossRef] [Green Version]
- Aguin-Pombo, D.; Rodrigues, M.C.; Voetdijk, B.; Breeuwer, J.A. Parthenogenesis and sex-ratio distorting bacteria in Empoasca (Hemiptera: Cicadellidae) Leafhoppers. Ann. Entomol. Soc. Am. 2021, 114, 738–749. [Google Scholar] [CrossRef]
- Zheng, Z.; Wang, D.; He, H.; Wei, C. Bacterial diversity of bacteriomes and organs of reproductive, digestive and excretory systems in two cicada species (Hemiptera: Cicadidae). PLoS ONE 2017, 12, e0175903. [Google Scholar] [CrossRef] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Knight, R.; Vrbanac, A.; Taylor, B.C.; Aksenov, A.; Callewaert, C.; Debelius, J.; Gonzalez, A.; Kosciolek, T.; McCall, L.-I.; McDonald, D.; et al. Best practices for analysing microbiomes. Nat. Rev. Microbiol. 2018, 16, 410–422. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, Y.; Hosokawa, T.; Fukatsu, T. Specific developmental window for establishment of an insect-microbe gut symbiosis. Appl. Environ. Microbiol. 2011, 77, 4075–4081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Illiano, P.; Brambilla, R.; Parolini, C. The mutual interplay of gut microbiota, diet and human disease. FEBS J. 2020, 287, 833–855. [Google Scholar] [CrossRef] [Green Version]
- Szablewski, L. Human Gut Microbiota in Health and Alzheimer’s Disease. J. Alzheimer Dis. 2018, 62, 549–560. [Google Scholar] [CrossRef]
- Tan, F.H.P.; Liu, G.; Lau, S.A.; Jaafar, M.H.; Park, Y.H.; Azzam, G.; Li, Y.; Liong, M.T. Lactobacillus probiotics improved the gut microbiota profile of a Drosophila melanogaster Alzheimer’s disease model and alleviated neurodegeneration in the eye. Benef. Microbes 2020, 11, 79–89. [Google Scholar] [CrossRef]
- Stasi, C.; Sadalla, S.; Milani, S. The Relationship Between the Serotonin Metabolism, Gut-microbiota and the Gut-brain Axis. Curr. Drug Metab. 2019, 20, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Strandwitz, P. Neurotransmitter modulation by the gut microbiota. Brain Res. 2018, 1693, 128–133. [Google Scholar] [CrossRef]
- Lakhan, S.; Caro, M.; Hadzimichalis, N. NMDA Receptor Activity in Neuropsychiatric Disorders. Front. Psychiatry 2013, 4, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Sharma, S.; Maurya, R.K.; De, T.D.; Thomas, T.; Lata, S.; Singh, N.; Pandey, K.C.; Valecha, N.; Dixit, R. Salivary glands harbor more diverse microbial communities than gut in Anopheles culicifacies. Parasites Vectors 2014, 7, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, D.-Y.; Liu, G.-H.; Cheng, T.-Y. Microbiome analysis of the saliva and midgut from partially or fully engorged female adult Dermacentor silvarum ticks in China. Exp. Appl. Acarol. 2020, 80, 543–558. [Google Scholar] [CrossRef] [PubMed]
- Mancini, M.V.; Damiani, C.; Accoti, A.; Tallarita, M.; Nunzi, E.; Cappelli, A.; Bozic, J.; Catanzani, R.; Rossi, P.; Valzano, M.; et al. Estimating bacteria diversity in different organs of nine species of mosquito by next generation sequencing. BMC Microbiol. 2018, 18, 126. [Google Scholar] [CrossRef] [Green Version]
- Backus, E.A.; Rangasamy, M.; Stamm, M.; McAuslane, H.J.; Cherry, R. Waveform Library for Chinch Bugs (Hemiptera: Heteroptera: Blissidae): Characterization of Electrical Penetration Graph Waveforms at Multiple Input Impedances. Ann. Entomol. Soc. Am. 2013, 106, 524–539. [Google Scholar] [CrossRef] [Green Version]
- Backus, E.A.; Holmes, W.J.; Schreiber, F.; Reardon, B.J.; Walker, G.P. Sharpshooter X Wave: Correlation of an Electrical Penetration Graph Waveform with Xylem Penetration Supports a Hypothesized Mechanism for Xylella fastidiosa Inoculation. Ann. Entomol. Soc. Am. 2009, 102, 847–867. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.; Wang, J.; Huang, J.; Zhang, S.; Vogler, A.P.; Liu, Q.; Li, Y.; Yang, M.; Li, Y.; Zhou, X. Host Phylogeny and Diet Shape Gut Microbial Communities Within Bamboo-Feeding Insects. Front. Microbiol. 2021, 12, 633075. [Google Scholar] [CrossRef]
- Anderson, K.E.; Sheehan, T.H.; Mott, B.M.; Maes, P.; Snyder, L.; Schwan, M.R.; Walton, A.; Jones, B.M.; Corby-Harris, V. Microbial ecology of the hive and pollination landscape: Bacterial associates from floral nectar, the alimentary tract and stored food of honey bees (Apis mellifera). PLoS ONE 2013, 8, e83125. [Google Scholar] [CrossRef] [Green Version]
- Dinant, S.; Bonnemain, J.-L.; Girousse, C.; Kehr, J. Phloem sap intricacy and interplay with aphid feeding. Comptes Rendus Biol. 2010, 333, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.-L.; Wang, T.-Z.; Zhu, H.-F.; Pan, H.-B.; Yu, X.-P. Diversity and dynamics of microbial communities in brown planthopper at different developmental stages revealed by high-throughput amplicon sequencing. Insect Sci. 2020, 27, 883–894. [Google Scholar] [CrossRef]
- Indiragandhi, P.; Yoon, C.; Yang, J.O.; Cho, S.; Sa, T.M.; Kim, G.H. Microbial communities in the developmental stages of B and Q biotypes of sweetpotato whitefly, Bemisia tabaci (hemiptera: Aleyrodidae). J. Korean Soc. Appl. Biol. Chem. 2010, 53, 605–617. [Google Scholar] [CrossRef]
- Martinez, A.J.; Onchuru, T.O.; Ingham, C.S.; Sandoval-Calderón, M.; Salem, H.; Deckert, J.; Kaltenpoth, M. Angiosperm to Gymnosperm host-plant switch entails shifts in microbiota of the Welwitschia bug, Probergrothius angolensis (Distant, 1902). Mol. Ecol. 2019, 28, 5172–5187. [Google Scholar] [CrossRef] [PubMed]
- Tsukimoto, M.; Nagaoka, M.; Shishido, Y.; Fujimoto, J.; Nishisaka, F.; Matsumoto, S.; Harunari, E.; Imada, C.; Matsuzaki, T. Bacterial Production of the Tunicate-Derived Antitumor Cyclic Depsipeptide Didemnin B. J. Nat. Prod. 2011, 74, 2329–2331. [Google Scholar] [CrossRef] [PubMed]
- Ahir, U.N.; Vyas, T.K.; Gandhi, K.D.; Faldu, P.R.; Patel, K.G. In Vitro Efficacy for Chlorpyrifos Degradation by Novel Isolate Tistrella sp. AUC10 Isolated from Chlorpyrifos Contaminated Field. Curr. Microbiol. 2020, 77, 2226–2232. [Google Scholar] [CrossRef]
- Walterson, A.M.; Stavrinides, J. Pantoea: Insights into a highly versatile and diverse genus within the Enterobacteriaceae. FEMS Microbiol. Rev. 2015, 39, 968–984. [Google Scholar] [CrossRef] [Green Version]
- Yuan, F.; Wei, C. Gene expression profiles in Malpighian tubules of the vector leafhopper Psammotettix striatus (L.) revealed regional functional diversity and heterogeneity. BMC Genom. 2022, 23, 67. [Google Scholar] [CrossRef]
- Szklarzewicz, T.; Michalik, A. Transovarial transmission of symbionts in insects. Oocytes Matern. Inf. Funct. 2017, 63, 43–67. [Google Scholar]
- Wang, D.; Wei, C. Bacterial communities in digestive and excretory organs of cicadas. Arch. Microbiol. 2020, 202, 539–553. [Google Scholar] [CrossRef]
- Harbecke, R.; Meise, M.; Holz, A.; Klapper, R.; Naffin, E.; Nordhoff, V.; Janning, W. Larval and imaginal pathways in early development of Drosophila. Int. J. Dev. Biol. 2003, 40, 197–204. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Q.; Shan, H.-W.; Chen, J.-P.; Wu, W. Diversity and Dynamics of Bacterial Communities in the Digestive and Excretory Systems across the Life Cycle of Leafhopper, Recilia dorsalis. Insects 2023, 14, 545. https://doi.org/10.3390/insects14060545
Huang Q, Shan H-W, Chen J-P, Wu W. Diversity and Dynamics of Bacterial Communities in the Digestive and Excretory Systems across the Life Cycle of Leafhopper, Recilia dorsalis. Insects. 2023; 14(6):545. https://doi.org/10.3390/insects14060545
Chicago/Turabian StyleHuang, Qiuyan, Hong-Wei Shan, Jian-Ping Chen, and Wei Wu. 2023. "Diversity and Dynamics of Bacterial Communities in the Digestive and Excretory Systems across the Life Cycle of Leafhopper, Recilia dorsalis" Insects 14, no. 6: 545. https://doi.org/10.3390/insects14060545