
Insecticidal Properties of Erythritol on Four Tropical Tephritid Fruit Flies, Zeugodacus cucurbitae, Ceratitis capitata, Bactrocera dorsalis, and B. latifrons (Diptera: Tephritidae)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Sugars
2.3. Feeding Assay
2.4. Experiment 1: Effect of Non-Nutritive Sugar on Fly Survival
2.5. Experiment 2: Effect of Sucrose Addition on Fly Survival
2.6. Experiment 3: Effect of Different Sugar Concentrations on Fly Survival
2.7. Experiment 4: Effect of Protein Availability on Fly Survival
2.8. Experiment 5: Effect of Water Availability on Fly Survival
2.9. Statistical Analyses
3. Results
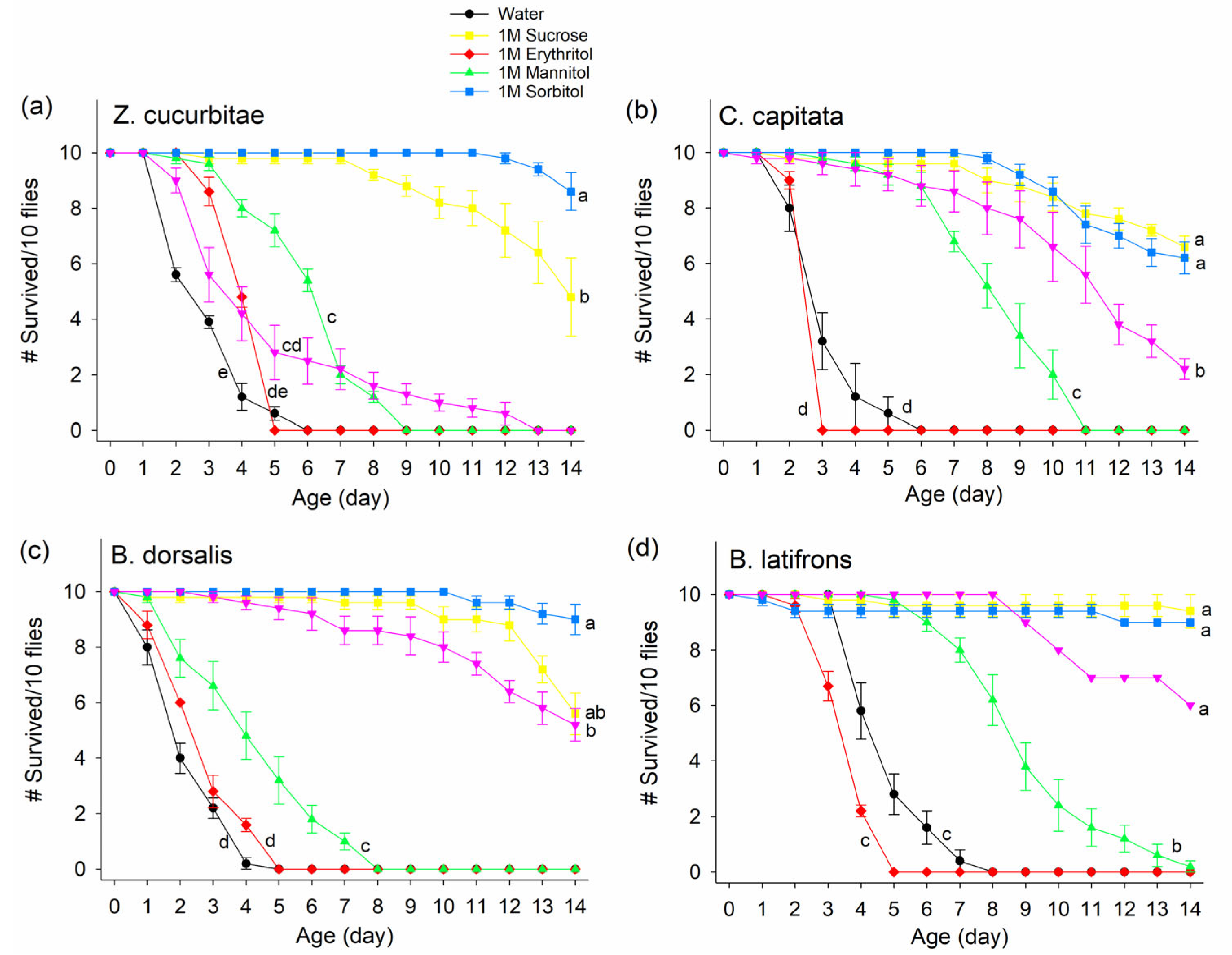
3.1. Experiment 1: Effect of Non-Nutritive Sugar on Fly Survival
3.2. Experiment 2: Effect of Sugar Concentration on Fly Survival
3.3. Experiment 3: Effect of Different Sugar Concentrations on Fly Survival
3.4. Experiment 4: Effect of Age and Protein Availability on Fly Survival
3.5. Experiment 5: Effect of Water Availability on Fly Survival
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shelly, T.; Epsky, N.D.; Jang, E.; Reyes-Flores, J.; Vargas, R. Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies: Lures, Area-Wide Programs, and Trade Implications; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Vargas, R.I.; Mau, R.F.L.; Jang, E.B.; Faust, R.M.; Wong, L. The Hawaii fruit fly area-wide pest management program. In Areawide Pest Management: Theory and Implementation; Koul, O., Cuperus, G.W., Elliott, N.C., Eds.; CABI Publishing: Wallingford, UK, 2008; pp. 300–325. [Google Scholar]
- Hardy, D.E. Studies in Hawaiian fruit flies (Diptera, Tephritidae). Proc. Entomol. Soc. Wash. 1949, 51, 181–205. [Google Scholar]
- Jang, E.B. Fruit Flies and Their Impact on Agriculture in Hawaii. Proc. Hawaii. Entomol. Soc. 2007, 39, 117–119. [Google Scholar]
- Papadopoulos, N.T.; Plant, R.E.; Carey, J.R. From trickle to flood: The large-scale, cryptic invasion of California by tropical fruit flies. Proc. Biol. Sci. 2013, 280, 20131466. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.W.; Vargas, R.I.; Prokopy, R.J.; Mackey, B.E. State-dependent attractiveness of protein bait and host fruit odor to Bactrocera cucurbitae (Diptera: Tephritidae) females Ann. Entomol. Soc. Am. 2004, 97, 1063–1068. [Google Scholar] [CrossRef]
- Vargas, R.I.; Pinero, J.C.; Leblanc, L. An Overview of Pest Species of Bactrocera Fruit Flies (Diptera: Tephritidae) and the Integration of Biopesticides with Other Biological Approaches for Their Management with a Focus on the Pacific Region. Insects 2015, 6, 297–318. [Google Scholar] [CrossRef]
- Moreno, D.S.; Mangan, R.L. Responses of the Mexican fruit fly (Diptera: Tephritidae) to two hydrolyzed proteins and incorporation of phloxine B to kill adults. In Light Activated Pest Control; ACS Publications: Washington, DC, USA, 1995; pp. 257–279. [Google Scholar]
- Stark, J.D.; Vargas, R.; Miller, N. Toxicity of spinosad in protein bait to three economically important tephritid fruit fly species (Diptera: Tephritidae) and their parasitoids (Hymenoptera: Braconidae). J. Econ. Entomol. 2004, 97, 911–915. [Google Scholar] [CrossRef]
- Guillem-Amat, A.; Sanchez, L.; Lopez-Errasquin, E.; Urena, E.; Hernandez-Crespo, P.; Ortego, F. Field detection and predicted evolution of spinosad resistance in Ceratitis capitata. Pest Manag. Sci. 2020, 76, 3702–3710. [Google Scholar] [CrossRef]
- Hsu, J.C.; Chou, M.Y.; Mau, R.F.; Maeda, C.; Shikano, I.; Manoukis, N.C.; Vargas, R.I. Spinosad resistance in field populations of melon fly, Zeugodacus cucurbitae (Coquillett), in Hawaii. Pest Manag. Sci. 2021, 77, 5439–5444. [Google Scholar] [CrossRef]
- Baudier, K.M.; Kaschock-Marenda, S.D.; Patel, N.; Diangelus, K.L.; O’Donnell, S.; Marenda, D.R. Erythritol, a non-nutritive sugar alcohol sweetener and the main component of Truvia®, is a palatable ingested insecticide. PLoS ONE 2014, 9, e98949. [Google Scholar] [CrossRef]
- Choi, M.Y.; Tang, S.B.; Ahn, S.J.; Amarasekare, K.G.; Shearer, P.; Lee, J.C. Effect of non-nutritive sugars to decrease the survivorship of spotted wing drosophila, Drosophila suzukii. J. Insect Physiol. 2017, 99, 86–94. [Google Scholar] [CrossRef]
- Sampson, B.J.; Werle, C.T.; Stringer, S.J.; Adamczyk, J.J. Ingestible insecticides for spotted wing Drosophila control: A polyol, erythritol, and an insect growth regulator, lufenuron. J. Appl. Entomol. 2017, 141, 8–18. [Google Scholar] [CrossRef]
- Zheng, C.; Zeng, L.; Xu, Y. Effect of sweeteners on the survival and behaviour of Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Pest Manag. Sci. 2016, 72, 990–996. [Google Scholar] [CrossRef]
- Fisher, M.L.; Fowler, F.E.; Denning, S.S.; Watson, D.W. Survival of the House Fly (Diptera: Muscidae) on Truvia and Other Sweeteners. J. Med. Entomol. 2017, 54, 999–1005. [Google Scholar] [CrossRef]
- Burgess, E.R.t.; King, B.H. Insecticidal Potential of Two Sugar Alcohols to Musca domestica (Diptera: Muscidae). J. Econ. Entomol. 2017, 110, 2252–2258. [Google Scholar] [CrossRef]
- Burgess, E.R.t.; Geden, C.J. Larvicidal potential of the polyol sweeteners erythritol and xylitol in two filth fly species. J. Vector Ecol. 2019, 44, 11–17. [Google Scholar] [CrossRef]
- Gilkey, P.L.; Bolshakov, D.T.; Kowala, J.G.; Taylor, L.A.; O’Donnell, S.; Marenda, D.R.; Sirot, L.K. Lethal effects of erythritol on the mosquito Aedes aegypti Linnaeus (Diptera: Culicidae). J. Appl. Entomol. 2018, 142, 873–881. [Google Scholar] [CrossRef]
- Maestas, L.; Lee, J.C.; Choi, M.-Y. Effect of erythritol and sucralose formulation on the survivorship of the mosquito, Aedes aegypti. Florida Entomol. 2023, 106. (in press). [Google Scholar]
- Wentz, K.; Cooper, W.R.; Horton, D.R.; Kao, R.; Nottingham, L.B. The Artificial Sweetener, Erythritol, Has Insecticidal Properties Against Pear Psylla (Hemiptera: Psyllidae). J. Econ. Entomol. 2020, 113, 2293–2299. [Google Scholar] [CrossRef]
- Price, B.E.; Breece, C.; Galindo, G.; Greenhalgh, A.; Sagili, R.; Choi, M.Y.; Lee, J.C. Nonnutritive Sugars for Spotted-Wing Drosophila (Diptera: Drosophilidae) Control Have Minimal Nontarget Effects on Honey Bee Larvae, a Pupal Parasitoid, and Yellow Jackets. Environ. Entomol. 2023, 52, 47–55. [Google Scholar] [CrossRef]
- Choi, M.Y.; Lucas, H.; Sagili, R.; Cha, D.H.; Lee, J.C. Effect of Erythritol on Drosophila suzukii (Diptera: Drosophilidae) in the Presence of Naturally-Occurring Sugar Sources, and on the Survival of Apis mellifera (Hymenoptera: Apidae). J. Econ. Entomol. 2019, 112, 981–985. [Google Scholar] [CrossRef]
- Schmidt-Jeffris, R.A.; Beers, E.H.; Smytheman, P.; Rehfield-Ray, L. Erythritol, an Artificial Sweetener, Is Acaricidal Against Pest Mites and Minimally Harmful to a Predatory Mite. J. Econ. Entomol. 2021, 114, 1701–1708. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.B.; Lee, J.C.; Jung, J.K.; Choi, M.Y. Effect of erythritol formulation on the mortality, fecundity and physiological excretion in Drosophila suzukii. J. Insect Physiol. 2017, 101, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Price, B.E.; Yoon, J.S.; Choi, M.Y.; Lee, J.C. Effects of nonnutritional sugars on lipid and carbohydrate content, physiological uptake, and excretion in Drosophila suzukii. Arch. Insect Biochem. Physiol. 2022, 109, e21860. [Google Scholar] [CrossRef] [PubMed]
- Munro, I.C.; Berndt, W.O.; Borzelleca, J.F.; Flamm, G.; Lynch, B.S.; Kennepohl, E.; Bar, E.A.; Modderman, J. Erythritol: An interpretive summary of biochemical, metabolic, toxicological and clinical data. Food Chem. Toxicol. 1998, 36, 1139–1174. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kearsley, M.W. Sweeteners and Sugar Alternatives in Food Technology, 2nd ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012. [Google Scholar]
- Sols, A.; Cadenas, E.; Alvarado, F. Enzymatic basis of mannose toxicity in honey bees. Science 1960, 131, 297–298. [Google Scholar] [CrossRef]
- De la Fuente, M.; Penas, P.F.; Sols, A. Mechanism of mannose toxicity. Biochem. Biophys. Res. Commun. 1986, 140, 51–55. [Google Scholar] [CrossRef]
- Caetano, F.H. Morfologia comparada do trato digestivo de formigas da subfamília Myrmicinae (Hymenoptera: Formicidae). Papéis Avulsos Zool. 1984, 35, 257–305. [Google Scholar]
- Nelson, D.L.; Cox, M.M. Lehniger Pricinpals of Biochemistry, 3rd ed.; Worth Publishers: Duffield, UK, 2000. [Google Scholar]
- Price, B.E.; Lee, J.C.; Choi, M.Y. Erythritol combined with non-nutritive sucralose increases feeding by Drosophila suzukii, quickens mortality and reduces oviposition. Crop Prot. 2021, 150, 105812. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cha, D.H.; Skabeikis, D.; Kim, B.-S.; Lee, J.C.; Choi, M.-y. Insecticidal Properties of Erythritol on Four Tropical Tephritid Fruit Flies, Zeugodacus cucurbitae, Ceratitis capitata, Bactrocera dorsalis, and B. latifrons (Diptera: Tephritidae). Insects 2023, 14, 472. https://doi.org/10.3390/insects14050472
Cha DH, Skabeikis D, Kim B-S, Lee JC, Choi M-y. Insecticidal Properties of Erythritol on Four Tropical Tephritid Fruit Flies, Zeugodacus cucurbitae, Ceratitis capitata, Bactrocera dorsalis, and B. latifrons (Diptera: Tephritidae). Insects. 2023; 14(5):472. https://doi.org/10.3390/insects14050472
Chicago/Turabian StyleCha, Dong H., Dominick Skabeikis, Bong-Soo Kim, Jana C. Lee, and Man-yeon Choi. 2023. "Insecticidal Properties of Erythritol on Four Tropical Tephritid Fruit Flies, Zeugodacus cucurbitae, Ceratitis capitata, Bactrocera dorsalis, and B. latifrons (Diptera: Tephritidae)" Insects 14, no. 5: 472. https://doi.org/10.3390/insects14050472